Гликолиз глюкозы и высвобождение энергии. Цикл лимонной кислоты или цикл Кребса
Добавил пользователь Дмитрий К. Обновлено: 28.01.2026
Цикл Кребса и гликолиз - два этапа клеточного дыхания. Клеточное дыхание - это биологическое окисление органического соединения, глюкозы для выделения химической энергии. Эта химическая энергия используется в качестве источника энергии в клеточных функциях. Цикл Кребса наступает после гликолиза. главное отличие между циклом Кребса и гликолиза является то, что Цикл Кребса участвует в полном окислении пировиноградной кислоты в диоксид углерода и воду, тогда как гликолиз превращает глюкозу в две молекулы пировиноградной кислоты, Цикл Кребса происходит внутри митохондрий у эукариот. Гликолиз происходит в цитоплазме всех живых организмов. Цикл Кребса также известен как цикл лимонной кислоты или же цикл трикарбоновых кислот (цикл ТСА), Гликолиз также известен как путь Embden-Meyerhof-Parnas (EMP).
Ключевые области покрыты
1. Что такое цикл Кребса (или цикл лимонной кислоты или цикл TCA)
- Определение, характеристики, процесс
2. Что такое гликолиз
- определение, характеристики, процесс
3. Каковы сходства между циклом Кребса и гликолизом
- Краткое описание общих черт
4. В чем разница между циклом Кребса и гликолизом
- Сравнение основных различий
Ключевые термины: ацетил-КоА, АТФ, клеточное дыхание, цикл лимонной кислоты, FADH, гликолиз, глюкоза, GTP, цикл Кребса, NADH, окислительное декарбоксилирование, пируват, цикл TCA
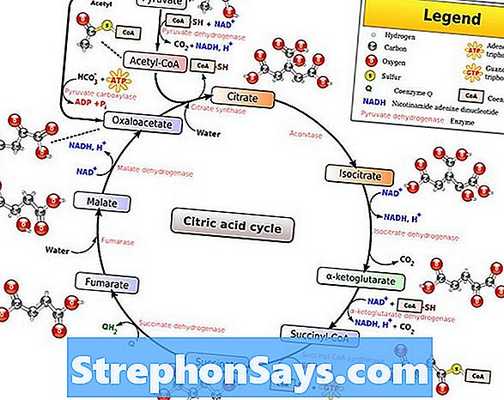
Что такое цикл Кребса?
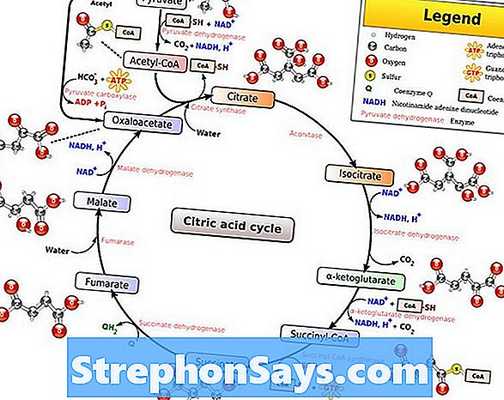
Цикл Кребса, также известный как цикл лимонной кислоты или же цикл трикарбоновых кислот (цикл ТСА)является вторым этапом аэробного дыхания в живых организмах. Во время цикла Кребса пируват полностью окисляется до углекислого газа и воды. Пируват образуется при гликолизе, который является первым этапом клеточного дыхания. Эти пируваты затем импортируются в матрицу митохондрий для прохождения окислительное декарбоксилирование, Во время окислительного декарбоксилирования пируват превращается в ацетил-КоА путем удаления молекулы диоксида углерода и окисления до уксусной кислоты. Затем коэнзим А присоединяется к уксусной части, образуя ацетил-КоА. Этот ацетил-КоА затем входит в цикл Кребса.
Рисунок 1: Окислительное декарбоксилирование цикла пирувата и Кребса
Во время цикла Кребса ацетильная часть ацетил-КоА присоединяется к молекуле оксалоацетата с образованием молекулы цитрата. Цитрат представляет собой молекулу из шести атомов углерода. Этот цитрат окисляется в результате ряда стадий, в результате чего из него выделяются две молекулы углекислого газа. Сначала лимонная кислота превращается в изоцитрат и окисляется до α-кетоглутарата путем восстановления NAD + молекулы. Α-Кетоглутарат снова окисляется до сукцинил-КоА. Сукцинил-КоА берет гидроксильную группу из воды и образует сукцинат. Сукцинат окисляется до фумарата с помощью FAD. Добавление молекулы воды к фумарату приводит к образованию малата. Затем малат окисляется обратно в оксалоацетат с помощью НАД + , Общие реакции цикла Кребса дают шесть NADH, два FADH2и две молекулы АТФ / ГТФ на одну молекулу глюкозы. Процесс окислительного декарбоксилирования наряду с циклом Кребса показан в Рисунок 1.
Что такое гликолиз
Гликолиз - это первая стадия клеточного дыхания у всех живых организмов. Это означает, что гликолиз происходит как при аэробном, так и при анаэробном дыхании. Гликолиз происходит в цитоплазме. Он участвует в расщеплении глюкозы на две молекулы пирувата. Фосфатная группа добавляется к молекуле глюкозы ферментом гексокиназой, образуя глюкозо-6-фосфат. Затем глюкозо-6-фосфат изомеризуется в фруктозо-6-фосфат. Фруктоза 6-фосфат превращается в фруктозу 1,6-бисфосфат. Фруктоза 1,6-бисфосфат под действием фермента альдозы расщепляется на дигидроксиацетон и глицеральдегид. Как дигидроксиацетон, так и глицеральдегид легко превращаются в дигидроацетонфосфат и глицеральдегид 3-фосфат. Глицеральдегид 3-фосфат окисляется до 1,3-бисфосфоглицерата. Одна фосфатная группа из 1,3-бисфосфоглицерата переносится в АДФ с образованием АТФ. Это приводит к образованию молекулы 3-фосфоглицерата. Фосфатная группа 3-фосфоглицерата переносится во второе положение углерода той же молекулы с образованием молекулы 2-фосфоглицерата. Удаление молекулы воды из 2-фосфоглицерата приводит к образованию фосфоенолпирувата (PEP). Перенос фосфатной группы PEP в молекулу ADP приводит к образованию пирувата.
Рисунок 2: Гликолиз
Полные реакции гликолиза производят две молекулы пирувата, две молекулы NADH, две молекулы АТФ и две молекулы воды. Полный процесс гликолиза показан в фигура 2.
Сходство между циклом Кребса и гликолизом
- Цикл Кребса и гликолиз - две стадии клеточного дыхания.
- И цикл Кребса, и гликолиз происходят в цитоплазме у прокариот.
- И цикл Кребса, и гликолиз приводятся в действие ферментами.
- И цикл Кребса, и гликолиз производят НАДН и АТФ.
Разница между циклом Кребса и гликолизом
Определение
Цикл Кребса: Цикл Кребса, также известный как цикл лимонной кислоты или цикл трикарбоновых кислот (цикл ТСА), относится к серии химических реакций, в которых пируват превращается в ацетил-КоА и полностью окисляется до диоксида углерода и воды.
Гликолиз: Гликолиз относится к серии химических реакций, в которых молекула глюкозы превращается в две молекулы пировиноградной кислоты.
Цикл Кребса: Цикл Кребса является вторым этапом клеточного дыхания.
Гликолиз: Гликолиз - это первая стадия клеточного дыхания.
Место нахождения
Цикл Кребса: Цикл Кребса происходит внутри митохондрий эукариот.
Гликолиз: Гликолиз происходит в цитоплазме.
Аэробное / анаэробное дыхание
Цикл Кребса: Цикл Кребса происходит только при аэробном дыхании.
Гликолиз: Гликолиз происходит как при аэробном, так и при анаэробном дыхании.
Процесс
Цикл Кребса: Цикл Кребса участвует в полном окислении пирувата в углекислый газ и воду.
Гликолиз: Гликолиз участвует в разложении глюкозы на две молекулы пирувата.
Линейный / Циклический
Цикл Кребса: Цикл Кребса является циклическим процессом.
Гликолиз: Гликолиз - это линейный процесс.
Конечный продукт
Цикл Кребса: Конечным продуктом цикла Кребса является неорганическое углеродное вещество.
Гликолиз: Конечным продуктом гликолиза является органическое вещество.
Расход АТФ
Цикл Кребса: Цикл Кребса не потребляет АТФ.
Гликолиз: Гликолиз потребляет две молекулы АТФ.
Чистая прибыль
Цикл Кребса: Цикл Кребса производит шесть молекул NADH и две FADH2 молекулы.
Гликолиз: Гликолиз производит две молекулы пирувата, две молекулы АТФ, две молекулы НАДН.
Чистая прибыль от энергии
Цикл Кребса: Чистый прирост энергии цикла Кребса равен 24 молекулам АТФ.
Гликолиз: Чистый прирост энергии гликолиза равен 8 молекулам АТФ.
Углекислый газ
Цикл Кребса: Углекислый газ выделяется в процессе цикла Кребса.
Гликолиз: В процессе гликолиза не выделяется углекислый газ.
Окислительного фосфорилирования
Цикл Кребса: Цикл Кребса связан с окислительным фосфорилированием.
Гликолиз: Гликолиз не связан с окислительным фосфорилированием.
кислород
Цикл Кребса: Цикл Кребса использует кислород в качестве конечного окислителя.
Гликолиз: Гликолиз не требует кислорода.
Заключение
Цикл Кребса и гликолиз - два этапа клеточного дыхания. Цикл Кребса происходит только при аэробном дыхании. Гликолиз характерен как для аэробного, так и для анаэробного дыхания. Цикл Кребса следует за гликолизом. Во время гликолиза две молекулы пирувата образуются из молекулы глюкозы. Эти молекулы пирувата полностью окисляются в углекислый газ и воду во время цикла Кребса. Основное различие между циклом Кребса и гликолизом заключается в исходных материалах, механизме и конечных продуктах каждого этапа.
Ссылка:
1. «Окислительное декарбоксилирование и цикл Кребса». Метаболические процессы. Hersi, Google Sites,
Гликолиз глюкозы и высвобождение энергии. Цикл лимонной кислоты или цикл Кребса
Физиология обмена глюкозы. Транспорт глюкозы через мембрану клетки
а) Главенствующая роль глюкозы в метаболизме углеводов. Конечными продуктами гидролиза углеводов в желудочно-кишечном тракте являются всего три вещества: глюкоза, фруктоза и галактоза. При этом на долю глюкозы приходится почти 80% общего количества этих моносахаридов. После всасывания в кишечнике большая часть фруктозы и практически вся галактоза преобразуются в печени в глюкозу. Вследствие этого в крови присутствуют только небольшие количества фруктозы и галактозы. В итоге процессов превращения глюкоза становится единственным представителем углеводов, транспортируемым во все клетки организма.
Соответствующие ферменты, необходимые клеткам печени для обеспечения процессов взаимного превращения моносахаридов — глюкозы, фруктозы и галактозы — показаны на рисунке ниже.
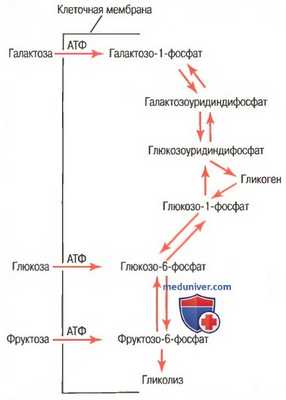
Взаимные превращения трех основных моносахаридов (глюкозы, фруктозы и галактозы) в клетках печени
В результате этих реакций, когда печень высвобождает моносахариды обратно в кровь, окончательным продуктом, попадающим в кровь, становится глюкоза. Причина этого явления заключается в том, что клетки печени содержат большое количество глюкозофосфатазы, поэтому глюкозо-6-фосфат может расщепляться на глюкозу и фосфат. Затем глюкоза транспортируется через мембраны клеток обратно в кровь.
Хотелось бы еще раз подчеркнуть, что обычно более 95% всех моносахаридов, циркулирующих в крови, представлены конечным продуктом превращения — глюкозой.
б) Транспорт глюкозы через мембрану клетки. Прежде чем глюкоза будет использована клетками тканей, она должна транспортироваться через мембраны клеток в цитоплазму. Однако глюкоза не может свободно диффундировать через поры в клеточных мембранах, т.к. максимальная молекулярная масса частиц должна быть в среднем равна 100, в то время как молекулярная масса глюкозы составляет 180. Тем не менее глюкоза может относительно легко проникать внутрь клеток благодаря механизму облегченной диффузии. Основы этого механизма обсуждались в главе 4, напомним его основные моменты.
Насквозь прободая липидную мембрану клеток, белки-переносчики, количество которых в мембране достаточно велико, могут взаимодействовать с глюкозой. В такой связанной форме глюкоза может транспортироваться белком-переносчиком с одной стороны мембраны на другую и там отделяться; если с одной стороны мембраны концентрация глюкозы выше, чем с другой, то глюкоза будет транспортироваться туда, где ее концентрация ниже, а не в противоположном направлении. Транспорт глюкозы через клеточные мембраны в большинстве тканей резко отличается от транспорта, который наблюдается в желудочно-кишечном тракте или в эпителиоцитах канальцевого аппарата почек.
В обоих упомянутых случаях транспорт глюкозы опосредован сопряженным с механизмом активного транспортом натрия. Активный транспорт натрия обеспечивает энергией процесс всасывания глюкозы против градиента концентрации. Такой сопряженный с натрием активный механизм транспорта глюкозы встречается только в специализированных эпителиоцитах, приспособленных для активного процесса абсорбции глюкозы. В других клеточных мембранах глюкоза транспортируется только из областей с высокой концентрацией в область низких концентраций с помощью механизма облегченной диффузии, возможность которого создается особыми свойствами расположенного в мембране белка-переносчика глюкозы.
Подробно механизм облегченной диффузии применительно к транспорту через клеточные мембраны изложен в отдельной статье на сайте (просим вас пользоваться формой поиска выше).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
а) Высвобождение энергии из молекулы глюкозы путем гликолиза. Полное окисление 1 грамм-молекулы глюкозы сопровождается выделением 686000 калорий энергии, при этом только 12000 калорий необходимо для образования 1 грамм-молекулы АТФ. Если бы сразу вся глюкоза окислялась до воды и углекислого газа при образовании 1 молекулы АТФ, потери энергии были бы неизбежны. К счастью, во всех клетках организма присутствуют особые белковые ферменты, обеспечивающие последовательное поэтапное расщепление молекулы глюкозы при образовании молекулы АТФ. При этом выделяемая на каждом этапе небольшими порциями энергия используется для образования АТФ, что обеспечивает образование 38 моль АТФ при окислении каждого моля глюкозы.
б) Гликолиз и образование пировиноградной кислоты. Едва ли не самый важный способ преобразования молекулы глюкозы, приводящий к высвобождению энергии, начинается процессом гликолиза. Конечные продукты гликолиза подлежат последующему окислению, что сопровождается высвобождением энергии. Гликолиз — это последовательность реакций, в результате которых одна молекула глюкозы расщепляется с образованием двух молекул пировиноградной кислоты.
Гликолиз обеспечивается 10 последовательными реакциями, представленными на рисунке. Каждый из этих этапов катализируется одним из специфических белков-ферментов. Обратите внимание, что глюкоза прежде всего превращается в фруктозо-1,6-дифосфат, а затем расщепляется на 2 молекулы, содержащие три атома углерода, глицеральдегид-3-фосфат, каждая из которых, пройдя пять дополнительных этапов, становится пировиноградной кислотой.
в) Образование АТФ во время гликолиза. При расщеплении молекулы глюкозы на большей части этапов выделяется очень небольшое количество свободной энергии, несмотря на многочисленность химических реакций, участвующих в процессе гликолиза. Однако на этапах преобразования 1,3-дифосфоглицериновой кислоты в 3-фосфоглицериновую кислоту и фосфоенолпировиноградной кислоты — в пировиноградную кислоту выделяются порции энергии более 12000 калорий на моль, чем необходимо для образования молекулы АТФ, поэтому эти этапы и сопровождаются образованием АТФ. В итоге из каждого моля фруктозо-1,6-фосфата при его расщеплении до пировиноградной кислоты образуются 4 моля АТФ.
Две молекулы АТФ необходимы для фосфорилирования исходной глюкозы при образовании фруктозо-1,6-дифосфата, т.е. для обеспечения начальных этапов гликолиза, поэтому чистый выход АТФ в процессе гликолиза составляет только 2 моля АТФ на каждый моль использованной глюкозы. При этом количество энергии, запасенной в виде АТФ, эквивалентно 24000 калорий. Во время гликолиза суммарно около 56000 калорий теряется на каждый использованный моль глюкозы, поэтому в целом эффективность этого механизма в пересчете на количество связанной в форме АТФ энергии составляет всего 43%. Остальные 57% энергии теряются в виде тепла.
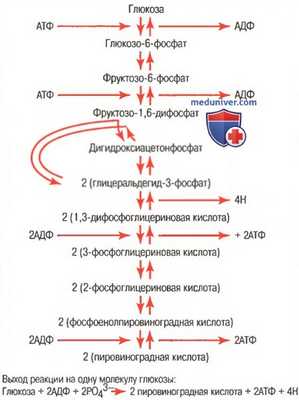
Последовательность химических реакций, ответственных за гликолиз
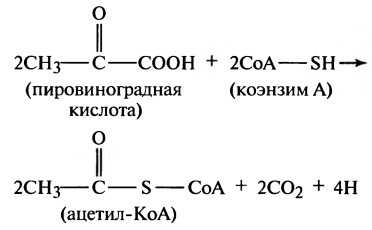
г) Превращение пировиноградной кислоты в ацетилкоэнзим А. Следующая стадия расщепления глюкозы включает два этапа превращения 2 молекул пировиноградной кислоты (для облегчения понимания просим вас изучить рисунок выше) в 2 молекулы ацетилкоэнзима А (ацетил-КоА) в соответствии с приводимыми уравнениями:
В результате этих реакций образуются 2 молекулы углекислого газа и 4 атома водорода, а 2 остатка молекул пировиноградной кислоты связываются с коэнзимом А (производным панто-теновой кислоты) с образованием ацетил-КоА. В итоге этих превращений АТФ не образуется, но в последующем, когда 4 высвободившихся атома водорода будут окислены, образуются 6 молекул АТФ, как будет показано далее.
д) Цикл лимонной кислоты (цикл Кребса). Следующая стадия расщепления глюкозы получила название цикла лимонной кислоты (другое название — цикл трикарбоновых кислот, или цикл Кребса). Этот цикл представляет собой последовательность химических реакций, в результате которых ацетил-КоА расщепляется до углекислого газа и атомов водорода. Эти реакции осуществляются в матриксе митохондрий. Атомы водорода, отщепляемые от промежуточных продуктов в реакциях дегидрирования при гликолизе и в цикле Кребса, будут последовательно окисляться (что обсуждается далее) с выделением огромного количества энергии в виде АТФ.
На рисунке ниже показаны различные этапы химических реакций цикла лимонной кислоты.

Химические реакции цикла лимонной кислоты (цикла Кребса), демонстрирующие высвобождение углекислого газа и количество атомов водорода, образующиеся в этом цикле
Вещества, показанные в левой части рисунка, вступают в химические реакции, а продукты этих реакций изображены в правой части рисунка. Заметьте, что верхняя часть колонки начала химических реакций представлена щавелево-уксусной кислотойу и в конце цепи реакций в основании колонки вновь появляется щавелево-уксусная кислота.
На начальной стадии цикла лимонной кислоты ацетил-КоА взаимодействует с щавелево-уксусной кислотой, образуя лимонную кислоту. Коэнзим А отделяется от ацетил-КоА и может использоваться вновь для образования новых молекул ацетил-КоА из пировиноградной кислоты.
Ацетильная часть может использоваться, становясь составной частью молекулы лимонной кислоты. На протяжении последующих стадий цикла лимонной кислоты в реакцию вступают молекулы воды, как показано в левой части рисунка. В итоге образуются углекислый газ и атомы водорода.
Суммарный итог реакций цикла лимонной кислоты приводится на рисунке. В итоге метаболических процессов из каждой исходной молекулы глюкозы получаются 2 молекулы ацетил-КоА, вступающие в реакции цикла лимонной кислоты наряду с 6 молекулами воды. В результате образуются 4 молекулы углекислого газа, 16 атомов водорода и 2 молекулы коэнзима А. Кроме того, образуются 2 молекулы АТФ.
Видео реакции цикла Кребса кратко и понятно
Регуляция обмена глюкозы. Синтез и распад гликогена
а) Инсулин увеличивает облегченную диффузию глюкозы. Скорость транспорта глюкозы, как и транспорта других моносахаридов, существенно увеличивается инсулином. Если поджелудочная железа продуцирует большие количества инсулина, скорость транспорта глюкозы в большинстве клеток возрастает более чем в 10 раз по сравнению со скоростью транспорта глюкозы при отсутствии инсулина. Напротив, при отсутствии инсулина количество глюкозы, которое может диффундировать внутрь большинства клеток, за исключением клеток мозга и печени, столь мало, что не в состоянии обеспечить нормальный уровень энергетических потребностей.
Скорость потребления глюкозы большинством клеток находится под контролирующим влиянием скорости продукции инсулина поджелудочной железой. Функции инсулина и его регуляторные влияния на обмен углеводов подробно изложены в отдельной статье на сайте (просим вас пользоваться формой поиска выше).
б) Фосфорилирование глюкозы. Как только глюкоза попадает в клетки, она связывается с фосфатными радикалами в соответствии со следующей схемой реакции:
Фосфорилирование осуществляется главным образом ферментом глюкокиназой в печени или гексокиназой в большинстве других клеток. Фосфорилирование глюкозы является почти полностью необратимой реакцией, исключая клетки печени, эпителиоциты почечного тубулярного аппарата и клетки кишечного эпителия, в которых присутствует другой фермент — глюкофосфорилаза. Будучи активирована, она может сделать реакцию обратимой. В большинстве тканей организма фосфорилирование служит способом захвата глюкозы клетками. Это происходит в связи со способностью глюкозы немедленно связываться с фосфатом, а в такой форме она не может выходить обратно из клетки, кроме некоторых особых случаев, в частности из клеток печени, которые располагают ферментом фосфатазой.
в) Гликоген запасается в печени и мышцах. После поступления внутрь клетки глюкоза практически немедленно используется клеткой для энергетических целей либо запасается в виде гликогена, который является крупным полимером глюкозы.
Все клетки организма способны запасать какие-то количества гликогена, но особенно большие его количества депонируются клетками печени, которая может запасать гликоген в количествах, составляющих от 5 до 8% веса этого органа, или мышечными клетками, содержание гликогена в которых составляет от 1 до 3%. Молекула гликогена может полимеризоваться так, что в состоянии иметь практически любую молекулярную массу; в среднем молекулярная масса гликогена составляет около 5 млн. В большинстве случаев гликоген, осаждаясь, образует крупные гранулы.
Превращение моносахаридов в преципитирующее соединение с большой молекулярной массой (гликоген) дает возможность запасать большие количества углеводов без заметного изменения осмотического давления во внутриклеточном пространстве. Высокая концентрация растворимых моносахаридов с низкой молекулярной массой могла бы приводить к катастрофическим последствиям для клеток в связи с формированием огромного градиента осмотического давления по обе стороны клеточной мембраны.
г) Гликогенез — процесс образования гликогена. Химические реакции образования гликогена показаны на рисунке ниже.
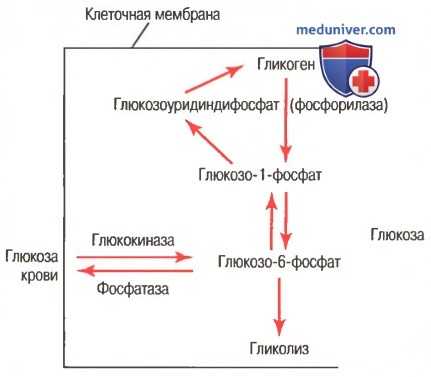
Химические реакции глюкогенеза и гликогенолиза. Показаны также взаимные превращения глюкозы крови и гликогена печени (фосфатаза требуется для высвобождения глюкозы из клеток и представлена в клетках печени, но отсутствует в большинстве других клеток)
На рисунке видно, что глюкозо-6-фосфат становится глюкозо-1-фосфатом, который затем превращается в глюкозоуридинфосфат, в итоге образующий гликоген. Для этих превращений необходимы специфические ферменты. Кроме того, и другие моносахариды, превращаясь в глюкозу, могут участвовать в образовании гликогена. Более мелкие соединения, включая молочную кислоту, глицерол, пировиноградную кислоту и некоторые дезаминированные аминокислоты, также могут превращаться в глюкозу или близкие ей соединения и затем становиться гликогеном.
д) Гликогенолиз — извлечение гликогена из депо. Процесс расщепления гликогена, хранящегося в клетках, который сопровождается высвобождением глюкозы, называют гликогенолизом. Затем глюкоза может использоваться в целях получения энергии. Гликогенолиз невозможен без реакций, обратных реакциям получения гликогена, при этом каждая вновь отщепляющаяся от гликогена молекула глюкозы подвергается фосфорилированию, катализируемому ферментом фосфорилазой. В состоянии покоя фосфорилаза пребывает в неактивном состоянии, поэтому гликоген может храниться в депо. Когда появляется необходимость получить глюкозу из гликогена, прежде всего должна активироваться фосфорилаза. Это может достигаться несколькими путями.
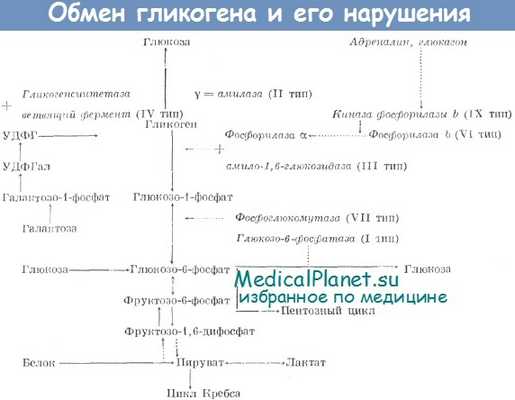
е) Активация фосфорилазы адреналином или глюкагоном. Два гормона — адреналин и глюкагон — могут активировать фосфорилазу и таким образом ускорять процессы гликогенолиза. Начальные моменты влияний этих гормонов связаны с образованием в клетках циклического аденозинмонофосфатау который затем запускает каскад химических реакций, активирующих фосфорилазу.
Адреналин выделяется из мозгового вещества надпочечников под влиянием активации симпатической нервной системы, поэтому одна из ее функций заключается в обеспечении обменных процессов. Эффект адреналина особенно заметен в отношении клеток печени и скелетных мышц, что обеспечивает наряду с влияниями симпатической нервной системы готовность организма к действию.
Глюкагон — гормон, выделяемый альфа-клетками поджелудочной железы, когда концентрация глюкозы в крови снижается до слишком низких значений. Он стимулирует образование циклического АМФ главным образом в клетках печени, что, в свою очередь, обеспечивает превращение в печени гликогена в глюкозу и ее высвобождение в кровь, повышая таким образом концентрацию глюкозы в крови.
Цикл лимонной кислоты (цикл Кребса)
высвобождается энергия Гиббса AG = -2880 кДж/моль. Эта энергия может запасаться в клетке в форме химической энергии фосфатных связей аденозилтрифос- фата АТР. Образующиеся молекулы АТР диффундируют в различные участки клетки, где используется энергия. АТР - это переносчик энергии. Клетка использует эту энергию для выполнения работы. Однако при гликолизе тратится лишь незначительная часть энергии, запасенной в глюкозе (несколько процентов). Основная ее часть передается в цикле Кребса (рис. 9.4), сопряженном с клеточным дыханием.
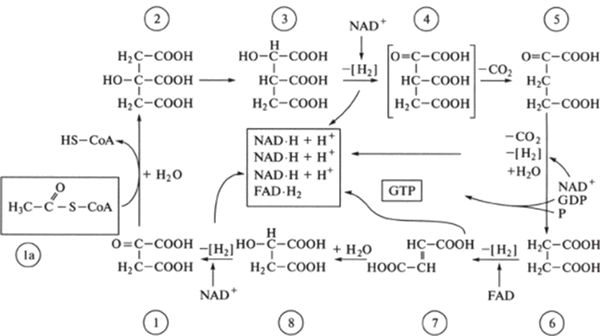
Рис. 9.4. Цикл Кребса (цикл лимонной кислоты)
I - оксалоацетат, 1а ацетил*СоЛ, 2 - лимонная кислота (цитрат). 3 - иэоцитрат. 4 - оксалосукцинат. 5 - кетоглугарат. 6 - янтарная кислота (сукцинат). 7 - фумарат. 8 - яблочная кислота (малат)
Цикл Кребса, или цикл лимонной кислоты, или цикл 3-карбоновых кислот, представляет собой ряд последовательных реакций, протекающих в митохондриях. В ходе этих реакций осуществляется катаболизм ацетильных групп СН3СО-, передаваемых от пирувата, конечного продукта гликолиза. Пируват вступает в реакции цикла Кребса, предварительно превращаясь в ацетил-СоА.
Цикл Кребса, как и гликолиз, представляет собой метаболический путь, состоящий из последовательных стадий - реакций. В отличие от гликолиза, этот путь замкнутый, циклический.
1. Ацетил-СоА - продукт катаболизма углеводов, белков и липидов - вступает в цикл, реагируя (конденсируется) с солью щавелевоуксусной кислоты (оксало- ацстатом). При этом образуется соль лимонной кислоты (цитрат):
2. Цитрат изомеризуется в изоцитрат. Реакция катализируется ферментом ако- нитазой и проходит через образование аконитата с последующим его превращением в изоцитрат:
3. Изоцитрат окисляется до а-кетоглутарата. Реакция катализируется ферментом изоцитратдегидрогеназой:
4. а-Кетоглутарат подвергается окислительному декарбоксилированию с образованием сукцинил-СоА. Катализируеся а-кетоглутарат дегидрогеназой:
5. Сукцинил-СоА превращается в сукцинат. Реакция катализируется ферментом сукцинат-СоА-лигазой:
6. Сукцинат превращается в фумарат. Реакция катализируется ферментом дегидрогеназой:
7. Фумарат гидратируется по двойной связи с образованием малата (соль яблочной кислоты). Катализируется фумаратгидратазой:
8. Манат окисляется до оксапоацетата. Катализируется мапатдегидрогеназой:
Рис. 9.5. Строение митохондрий (увеличение примерно 25 000)
На восьмой стадии цикл замыкается и начинается его новое прохождение.
Все стадии цикла лимонной кислоты протекают во внутренней среде митохондрий - матриксе (рис. 9.5). Здесь находятся все ферменты этого метаболического пути.
Митохондрия (от греч. «mitos» - нить и «chondrion» - зернышко) имеет вытянутую форму; длина 1,5-2 мкм, диаметр 0,5-1 мкм. Органеллы клеток животных находятся в жидкой среде клетки - цитоплазме (см. рис. 6.2).
Внутреннее пространство митохондрий окружено двумя непрерывными мембранами. При этом наружная мембрана гладкая, а внутренняя образует многочисленные складки, или кристы. Внутримитохондриальное пространство ограничено внутренней мембраной, заполнено жидкой средой - матриксом, который примерно на 50% состоит из белка и имеет очень тонкую структуру. Удлиненная форма митохондрий не универсальна. В некоторых тканях, например в поперечно-полосатых скелетных мышцах, митохондрии иногда принимают самые причудливые очертания.
В митохондриях сосредоточено большое количество ферментов.
В клетке может находиться от нескольких сотен до нескольких десятков тысяч митохондрий. Для одного и того же типа клеток число митохондрий более или менее постоянно. Однако следует помнить, что количество митохондрий может меняться в зависимости от стадии развития клетки и ее функциональной активности, а в целом от интенсивности нагрузок на организм.
Митохондрии - энергетические станции, вырабатывающие энергию для жизнедеятельности организма. Особенно много митохондрий в мышечных клетках, где требуются большие затраты энергии.
Образованные в цикле Кребса высокоэнергетические вещества NADH и FADFb (см. рис. 9.4) передают свою энергию в реакции ресинтеза АТР из ADP:
В результате на каждую молекулу NADH образуются 3 молекулы АТР. Эта реакция окислительно-восстановительная, т. е. сопровождается переносом электронов от восстановителя NADH к окислителям (см. разд. 4.3). В качестве окислителя выступает кислород О2. Эта реакция называется окислительным фосфорилированием ADP в АТР.
Окислительное фосфорилирование происходит во внутренней митохондриальной мембране. В трех участках дыхательной цепи запасается энергия в результате синтеза АТР из ADP и Р,.
Реакция протекает в несколько стадий на внутренних мембранах митохондрий (см. рис. 9.5), в системе ферментов, называемой дыхательной цепью. Сюда из клеточной плазмы поступают молекулы ADP. Соответствующий окислительновосстановительный процесс называется клеточным дыханием. Именно здесь расходуется кислород, которым мы дышим.
Молекулы АТР, образованные в матриксе, выходят из митохондрий в плазму клетки, где участвуют в различных биохимических реакциях, протекающих с расходом энергии.
Таким образом, энергия, высвобождающаяся в процессе переноса электронов от восстановителей, используется для окислительного фосфорилирования ADP в АТР.
Предполагают, что энергия, высвобождающаяся вдыхательной цепи, затрачивается непосредственно на перевод внутренней мембраны в новое, богатое энергией конформационное состояние, которое, в свою очередь, становится движущей силой окислительного фосфорилирования, приводящего к образованию АТР. В настоящее время наиболее серьёзное обоснование получила гипотеза хемоосмо- тического сопряжения Митчела.
Таким образом, биосинтез АТР в животном организме осуществляется из ADP и неорганического фосфата Р, при активировании последнего за счёт энергии окисления органических соединений при метаболических процессах.
Окисление органических соединений в живых системах не всегда сопряжено с фосфорилированием, и фосфорилирование не обязательно должно быть окислительным.
Известно несколько сотен реакций окисления. Не менее десятка из них сопряжено с одновременным активированием неорганического фосфата. Такие реакции называют реакциями субстратного фосфорилирования. Здесь реакции расщепления субстрата сопровождаются передачей энергии непосредственно неорганическому фосфату. В результате образуется другой фосфорилированный субстрат с макроэргической связью. В этом случае в процессе не участвует дыхательная цепь ферментов и не происходит превращение энергии, выделяемой при переносе электронов на кислород, в энергию фосфатной связи АТР.
В качестве примера субстратного фосфорилирования можно привести реакцию превращения сукницил-СоА в янтарную кислоту с образованием GTP из GDP и фосфата Р, в лимоннокислом цикле.
В растениях источником энергии для активирования неорганического фосфата и обеспечения синтеза АТР служит энергия солнечного света, улавливаемая фото- синтетическим аппаратом клетки. Такое фосфорилирование называют фотосин- тетическим.
Для удовлетворения потребностей человеческого организма в энергии молекулы АТР на протяжении суток тысячи и тысячи раз расщепляются до молекул ADP и Р, с последующим ресинтезом АТР. Кроме того, скорость ресинтеза АТР должна меняться в широких пределах - от минимальной во время сна до максимальной в периоды напряженной мышечной работы.
Из сказанного можно сделать вывод, что окислительное фосфорилирование не просто непрерывный жизненно важный процесс. Он должен регулироваться в широких пределах, что достигается путем тренировки.
Суммарное уравнение реакций гликолиза и цикла лимонной кислоты записывается следующим образом:
Стандартная энергия Гиббса окисления 1 моля глюкозы СбН^Об равна ДG* = = -2880 кДж (см. разд. 5.1). Стандартная энергия Гиббса гидролиза 38 молей АТР (запасенная энергия) равна ДG°' = -38*30 = -1180 кДж, т. е. запасается лишь 40% энергии глюкозы (коэффициент полезного действия дыхания). Остальная энергия выделяется из организма в виде тепла Q. Этим объясняется разогрев и повышение температу ры тела при интенсивной работе (см. рис. 5.2).
Глюкоза выполняет функцию клеточного топлива в нашем организме. Она получается главным образом либо в процессе пищеварения из углеводов, либо путем синтеза из резервных жиров.
Читайте также: