Гормональная регуляция сперматогенеза. Лютропин аденогипофиза. Стероидогенный эффект лютропина. Ингибин. Активин. Функция эстрогенов.
Добавил пользователь Владимир З. Обновлено: 28.01.2026
Гормональная регуляция сперматогенеза многообразна (рис. 19–6). Гипоталамо-гипофизарная система при помощи гонадолиберина активирует синтез и секрецию гонадотропных гормонов гипофиза, влияющих на активность клеток Ляйдига (синтез и секреция тестостерона) и Сертоли (синтез и секреция ингибинов, подробнее см. выше в разделе «Сперматогенез»). В свою очередь вырабатываемые в яичке тестостерон и ингибины корректируют эндокринную деятельность гипоталамо-гипофизарной системы.
Тестостерон. Сперматогенез в яичках поддерживает тестостерон — главный активатор этого процесса.
ФСГ. Для полноценного осуществления сперматогенеза необходим также ФСГ. Мишенью ФСГ в извитых семенных канальцах служат клетки Сертоли. ФСГ поступает в интерстиций яичка по мелким артериолам (рис. 19–8), затем диффундирует через базальную мембрану извитых канальцев и связывается со специфическими мембранными рецепторами на клетках Сертоли. Стимуляция рецепторов ФСГ приводит к синтезу внутриклеточных рецепторов андрогенов и андроген-связывающего белка (АСБ). Затем клетки Сертоли секретируют АСБ, и он связывает тестостерон, образуемый клетками Ляйдига и диффундирующий внутрь извитых семенных канальцев. АСБ переносит андрогены к сперматогенным клеткам, где они воздействуют на премейотические клетки (сперматоциты I порядка), имеющие андрогеновые рецепторы. Зависимость функций клеток Сертоли от ФСГ сопоставима с функциями их гомологов (фолликулярных клеток) в яичнике. Как и фолликулярные клетки яичника, клетки Сертоли секретируют ингибины. Ингибины вместе с тестостероном тормозят образование ФСГ у мужчин.
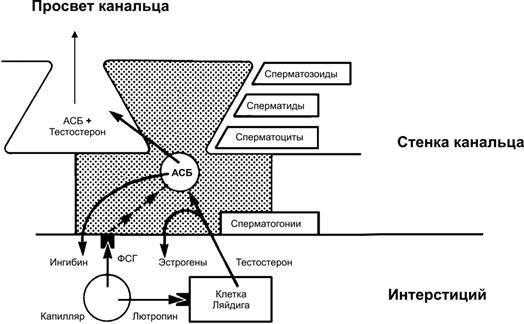
Рис. 19–8. Гормональная регуляция секреторной функции клеток Сертоли (заштрихована на рисунке). Фоллитропин стимулирует секрецию андроген-связывающего белка (АСБ), поддерживающего высокий уровень тестостерона в верхних слоях сперматогенного эпителия. Часть тестостерона в клетках Сертоли путём ароматизации превращается в эстрогены. [11].
Функции клеток Ляйдига. Клетки Ляйдига дозозависимым образом отвечают на стимуляцию ЛГ синтезом тестостерона. Кроме рецепторов ЛГ, клетки Ляйдига имеют рецепторы пролактина и ингибинов. Эти гормоны усиливают стимулирующий эффект ЛГ на образование тестостерона, но без ЛГ синтеза тестостерона не происходит.
Гонадолиберин
Гонадолиберин синтезируется в нейросекреторных клетках гипоталамуса. Синтез гонадолиберина подавляют тестостерон и ингибины (см. рис. 19–6). Гонадолиберин по аксонам нервных клеток транспортируется к срединному возвышению и поступает в кровь из аксонов нейросекреторных клеток в пульсирующем режиме с пиковыми интервалами около двух часов. Достигая по гипоталамо-гипофизарной системе кровотока передней доли гипофиза, гонадолиберин активирует синтезирующие ФСГ и ЛГ эндокринные клетки (см. рис. 19–6).
Гипофизарные гонадотропины
Гонадотропные гормоны (фолликулостимулирующий — ФСГ и лютеинизирующий — ЛГ), как и гонадолиберин, высвобождаются в кровь в пульсирующем режиме, что особенно характерно для ЛГ, эпизоды подъёма концентрации которого в крови мужчин происходят с интервалами 90–120 минут.
Регуляторы синтеза и секреции (см. рис. 19–6). Секреция гонадотропных гормонов контролируется как гонадолиберином (активирует), так и тестикулярными гормонами (подавляют). Супрессорный эффект тестостерона на секрецию ЛГ в основном проявляется на уровне гипоталамуса (через синтез гонадолиберина), тогда как эстрогены снижают чувствительность гонадотропных клеток к гонадолиберину. В то же время половые стероиды слабо влияют на секрецию ФСГ, в то время как ингибины оказывает выраженное подавляющее действие на гонадотропные клетки, синтезирующие ФСГ.
Мишени гонадотропных гормонов — яички. Клетки Сертоли имеют рецепторы ФСГ, а клетки Ляйдига — ЛГ.
ФСГ активирует в клетках Сертоли синтез и секрецию андроген-связывающего белка, ингибинов, эстрогенов, трансферрина, активаторов плазминогена (см. рис. 19–8).
Лютропин. Клетки Ляйдига имеют рецепторы ЛГ. ЛГ стимулирует в клетках Ляйдига синтез и секрецию тестостерона и отчасти эстрогенов.
Гормональная регуляция сперматогенеза. Лютропин аденогипофиза. Стероидогенный эффект лютропина. Ингибин. Активин. Функция эстрогенов.
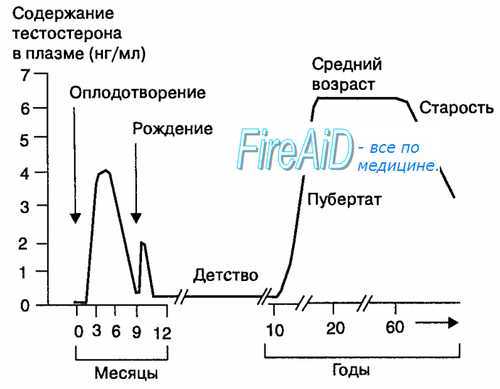
В пубертатный период в мужском организме происходит развитие и становление репродуктивной функции, характерной для организма взрослого мужчины. При этом во время пубертатного периода в коре надпочечников начинается секреция адреналовых мужских половых гормонов — андрогенов. В среднем в 11-летнем возрасте у мальчиков в семенниках начинается увеличение количества клеток Лейдига и благодаря их функции уровень секреции тестостерона постепенно достигает плато взрослого мужчины (порядка 6 нг/мл плазмы крови). В течение пубертатного периода происходит рост и развитие тела, изменение в регуляции многих функций организма, в том числе ЦНС и гипоталамуса. Секреция гонадолиберина в гипоталамусе и гонадотропинов (фоллитропина и лютропина) в гипофизе постепенно достигает уровня взрослого мужчины и сохраняется на относительно постоянном уровне в среднем до 50 лет, обусловливая характер синтеза и секреции тестестерона в мужских половых железах (рис. 16.3). Рис. 16.3. Изменение концентрации тестостерона в плазме крови мужского организма в различные периоды жизни. На протяжении жизни мужского организма секреторная функция клеток Лейдига существенно изменяется. Высокая концентрация тестостерона отмечается в поздний период внутриутробного развития (4 нг/мл), когда он необходим для регуляции интенсивного развития наружных половых органов. Низкий уровень тестостерона в плазме крови поддерживается до пред-пубертатного периода, поскольку отсутствуют клетки Лейдига. Следующий подъем секреции тестостерона отмечается в 11 лет и удерживается в течение пубертатного периода. По окончании пубертатного периода секреция тестостерона достигает уровня взрослого мужчины и сохраняется вплоть до 50 лет жизни.
Функции семенников
Основными функциями мужских гонад являются сперматогенез и секреция мужских половых гормонов. Активность мужской репродуктивной функции является непрерывной и не имеет циклических проявлений, как в женском организме. Семенники развиваются в брюшной полости плода, но примерно за 2—3 мес до рождения под влиянием нарастающей концентрации тестостерона в плазме крови семенники опускаются в мошонку. В мошонке температура составляет примерно 34 °С и ниже, что является адекватной температурной средой для нормального течения сперматогенеза. Оптимальная для сперматогенеза температура в яичках регулируется в процессе противоточного температурного обмена между притекающей артериальной кровью и оттекающей от тканей яичек венозной кровью, а также при сокращении или расслаблении гладкой мышцы, поднимающей яичко, что изменяет положение последнего относительно области центральной температуры тела.
Сперматогенез осуществляется в семенных канальцах, стенки которых выстланы сперматогенным эпителием, содержащим сперматозоиды и предшествующие им клетки — сперматогонии, сперматоциты I и II порядка, а также сперматиды, которые расположены между поддерживающими клетками Сертоли (рис. 16.4). Плотные контакты между клетками Сертоли создают надежный барьер с кровью (гематотестикулярный), который препятствует развитию аутоиммунных реакций организма на антигены, расположенные на мембране развивающихся половых клеток, а также действию на сперматогенный эпителий токсичных веществ, попадающих в организм из внешней среды. Клетки Сертоли выполняют трофическую функцию в отношении развивающегося сперматозоида, фагоцитируют половые клетки, утратившие способность к делению, а также остатки цитоплазмы делящихся половых клеток.
Таблица 16.2. Роль основных андрогеиов в мужской репродуктивной функции.
| Гормон | Внутриутробный период | Пубертатный период | Взрослый мужчина |
| Тестостерон | Стимуляция дифференциации: семявынося-щие протоки семенные пузырьки придатки яичек | Стимуляция пубертатного роста организма Рост пениса, семенных трубочек Сперматогенез Секреция гонадотропинов Регуляция по механизму обратной связи: гипоталамус-гипофиз—половые железы Формирование вторичных половых признаков: развитие голосового аппарата, мышц, распределение жира | Сперматогенез Секреция гонадотропинов Половые стимулы Половое поведение Регуляция по механизму обратной связи: гипоталамус-гипофиз—половые железы Ткани пениса, семенных пузырьков Вторичные половые признаки: голос, мышцы, распределение жира Количество эритроцитов к крови и содержание в плазме крови липидов |
| Дигидротестостерон | ------------------- | Рост волос по мужскому типу Развитие сальных желез, простаты | Сперматогенез Секреция гонадотропинов Половые стимулы Половое поведение Наружные половые органы Семенные пузырьки Регуляция по механизму обратной связи: гипоталамус-гипофиз—половые железы |
Клетки Сертоли синтезируют белок, способный связывать тестостерон. Это поддерживает высокую концентрацию в сперматогенном эпителии гормона, необходимого для стимуляции сперматогенеза.
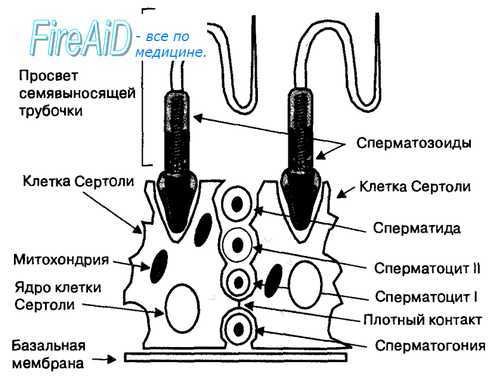
Рис. 16.4. Расположение сперматогенных клеток в эпителии семявыносящей трубочки. Семявыносящая трубочка окружена базальной мембраной, на которой находятся клетки Сертоли и сперматогенные клетки (сперматогонии, сперматоциты I и II порядка, сперматиды). Между клетками Сертоли расположены сперматогенные клетки, находящиеся на различной стадии деления (митоз — сперматогонии и мейоз — сперматоциты I и II порядка) и морфологической дифференциации (сперматиды). Плотные контакты между клетками Сертоли представляют эффективный барьер между кровью и внутренней средой семявыносящей трубочки, препятствуя попаданию вырабатываемых половых клеток в кровь и развитию иммунной реакции мужского организма на эти клетки.
Клетки Лейдига синтезируют мужские половые гормоны андрогены и находятся в соединительной ткани между семенными канальцами. Высокая активность этих клеток и соответственно высокая секреция тестостерона имеет место в поздний период развития плода, что необходимо для развития у него наружных половых органов. После рождения, к концу первого года жизни происходит интенсивная регрессия количества клеток Лейдига и снижение секреции в гонадах ребенка тестостерона в связи со снижением секреции лютропина в гипофизе. В результате до начала пубертатного периода в организме ребенка мужского пола не развиваются первичные и вторичные половые признаки, которые характерны для нормального взрослого организма мужчины. Начало пубертатного периода в развитии мужского организма связано с началом секреции гонадолиберина в гипоталамусе и лютропина в аденогипофизе. Под влиянием этих гормонов, клетки Лейдига дифференцируются во взрослый тип и в них начинается секреция тестостерона и дигидротестостерона, обусловливающих половое развитие мужского организма (табл. 16.2).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Сперматогенез стимулирует лютропин аденогипофиза. Под влиянием лютропина интерстициальные клетки Лейдига секретируют мужской половой гормон тестостерон. Гормон диффундирует через базальную мембрану в семенные канальцы, связывается со специфическим белком, который секретируется клетками Сертоли, и, таким образом, накапливается в сперматоген-ном эпителии, стимулируя сперматогенез. Концентрация тестостерона во внутренней среде семенных канальцев в 100 раз выше, чем в плазме крови. Тестостерон, образующийся в клетках Лейдига, поступает в клетки Сертоли и в них превращается в более активный гормон — дигидротестостерон, который транспортируется в жидкость канальцев, и в присутствии которого происходит деление сперматогоний.
Тестостерон стимулирует формирование метаботропных рецепторов фоллитропина в клетках Сертоли. Фоллитропин через эти рецепторы активирует клетки Сертоли, благодаря трофической и барьерной функции которых осуществляются процессы митотических и мейоти-ческих делений, а также спер-миогенез мужских половых клеток. Гормон роста аденоги-пофиза через метаботропные рецепторы мембраны клеток Сертоли стимулирует их метаболическую активность, а в отношении сперматогоний — активирует начало их митотического деления. Под влиянием фоллитропина в клетках Сертоли тестостерон ароматизируется в эстрадиол-17(3, который поддерживает нормальное течение спермио-генеза мужских половых клеток. Фоллитропин и тестостерон являются си-нергистами стимуляции в клетках Лейдига синтеза белка, который транспортирует эстрогены и тестостерон от клеток Лейдига в жидкость, заполняющую семенные канатики, в которых эти гормоны активируют сперма-тонгенез.
На рис. 16.6 представлена общая схема стероидогенеза, происходящего в клетках Лейдига и Сертоли.
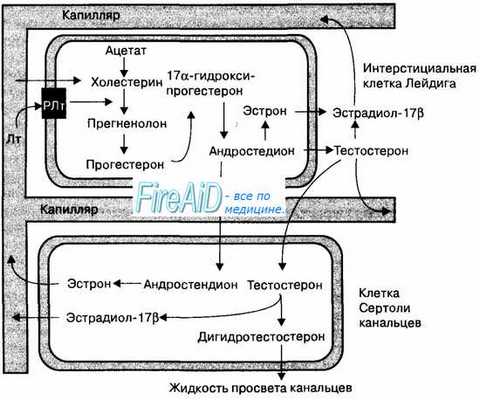
Рис. 16.6. Схема стероидогенеза в клетках мужских половых желез. Биосинтез основного мужского полового гормона тестостерона происходит в клетках Лейдига. Импульсная секреция лютропина (Лт) в аденогипофизе стимулирует в клетках Лейдига секрецию тестостерона, концентрация которого в плазме крови также имеет незначительные колебания в течение суток. Лютропин стимулирует секрецию клеток Лейдига путем связывания с метаботропными рецепторами на их мембране (РЛт). Превращение тестостерона в эстрадиол-17b и дигидротестостерон происходит в клетках Сертоли. Гормон эстрон синтезируется в клетках Сертоли из андростендиона, образующегося в клетках Лейдига. Тестостерон и другие мужские половые гормоны транспортируются в кровь и жидкость семенных канальцев, оказывая физиологические эффекты на ткани мужского организма.
Лютропин контролирует метаболическую активность клеток Лейдига, связываясь, с мембранными рецепторами системы G-белков, которые активируют энзимы аденилатциклазу и фосфолипазу С. Вторичным посредником действия лютропина на клетки Лейдига является цАМФ. При этом в клетках Лейдига в первые минуты действия гормона возникает быстрый стероидогенный эффект. Эффект проявляется в резком увеличении содержания цАМФ и росте продукции тестостерона за счет активации перемещения холестерина из цитозоля клеток на внутреннюю мембрану митохондрий. Лютропин оказывает метаботропный (трофический) долговременный эффект на структуру и функцию клетки Лейдига. Тестостерон, который синтезируется в клетках Лейдига, под влиянием лютропина диффундирует через базальную мембрану в семенные канатики, где его нормальная концентрация необходима для процесса сперматогенеза. Гормон аденогипо-физа пролактин и фактор роста ингибин, секретируемые в клетках Сертоли в качестве модуляторов, повышают стимулирующее действие лютропина на функцию клеток Лейдига.
В клетках Сертоли секретируются так называемые факторы роста ингибин и активин, которые тормозят по механизму отрицательной обратной связи секрецию гонадотропинов в аденогипофизе, а следовательно, сперматогенез.
Функция эстрогенов. Концентрация эстрогенов (эстрон и эстрадиол-17р) в плазме крови мужского организма поддерживается за счет их поступления в кровь из клеток Сертоли (ароматизируются из андрогенов), клеток пучковой и сетчатой зон коры надпочечников.
Эстрогены по механизму отрицательной обратной связи тормозят синтез тестостерона в клетках Лейдига, в результате уменьшается активность всех стадий сперматогенеза, а также количество рецепторов фоллитропина на мембране клеток Лейдига и синтез в них адроген-транспортного белка.
Кроме того, эстрогены через мембранные и ядерные эстрогенные рецепторы р-типа оказывают влияние на зрелые клетки Лейдига семенников взрослых мужчин, а также на сперматогонии, сперматоциты и развивающиеся сперматиды. Однако эстрогены не способны оказывать свое физиологическое действие на зрелые сперматозоиды, поскольку в их ядре нет эс-трогеновых рецепторов. С (3-типом эстрогенных рецепторов на мембране клеток Лейдига способны связываться такие внеклеточные гуморальные субстанции, как пептидные факторы роста и цитокины, и, таким образом, стимулировать секреторную функцию этих клеток. Роль эстрогенов заключается в модуляции образования рецепторов к аквапорину-I на мембране клеток проксимальных отделов протоков придатков яичек. Под влиянием аквапорина-1 увеличивается реабсорбция воды из жидкости семенных канальцев и, таким образом, регулируется нормальное осмотическое давление в канальцевой среде, окружающей зрелые сперматозоиды.
Сперматогенез. Стадия размножения ( сперматогония ). Стадия развития. Стадия созревания ( спермиогенез ). Структурные компоненты сперматозоида.
В пубертатный период развития мужского организма активируется продукция сперматозоидов, которая сохраняется на протяжении репродуктивного периода жизни мужчины вплоть до глубокой старости. При этом ежедневно продуцируется 200—400 млн. сперматозоидов. Сперматогенез, или образование зрелых сперматозоидов, происходит в семенных канальцах на протяжении стадий размножения, развития и созревания. Стадия размножения начинается с митотического деления сперматогонии (см. рис. 16.1). Несколько митотических делений сменяются двумя мейозами, во время которых диплоидное число хромосом (первичные сперматоциты) уменьшается наполовину, т. е. становится гаплоидным (сперматоциты I и II порядка). В стадии развития сперматоциты остаются связанными между собой цитоплазматическими мостиками, которые исчезают только на финальной стадии сперматогенеза. Одна первичная сперматогония образует в результате деления 16 сперматид. В стадии созревания (спермиогенез) одна спер-матида продуцирует один сперматозоид. Зрелый сперматозоид обладает способностью транспортировать хромосомы к ооциту, после того как он окажется в женском репродуктивном тракте. Период превращения сперматогоний в зрелые сперматозоиды продолжается 70—80 сут.
Рис. 16.1. Схема сперматогенеза и оогенеза. Первичные сперматоциты образуются путем митоза из сперматогоний и содержат диплоидное число хромосом. В результате первого мейотического деления образуются сперматоциты, содержащие гаплоидное число двухроматидных хромосом (22 единичные соматические хромосомы и одна половая — X или Y). После второго мейотического деления образуются сперматиды, содержащие гаплоидный набор монохроматидных хромосом. Оогенез начинается со стадии первичного ооцита, содержащего диплоидное число хромосом. Первичный ооцит путем первого мейотического деления превращается во вторичный ооцит, содержащий гаплоидное число двухроматидных хромосом (22 единичные соматические хросомы и одна половая Х-хро-мосома). В процессе оплодотворения ооцита сперматозоидом образуется зигота, содержащая 22 пары соматических хромосом и одну пару половых (XX или XY). Таким образом, в зиготе восстанавливается диплоидный набор хромосом. Рис. 16.5. Схематическое изображение строения зрелого сперматозоида. Головка сперматозоида содержит ядро с гаплоидным набором хромосом и имеет акросому. Акросома в процессе акросомальной реакции выделяет энзимы, с помощью которых сперматозоид проникает через прозрачную оболочку внутрь ооцита во время его оплодотворения. Средний отдел сперматозоида содержит митохондрии, которые производят энергию для двигательных филамент жгутика. Зрелый сперматозоид способен доставлять в женском репродуктивном тракте хромосомы к ооциту благодаря двигательной активности жгутика.
Зрелый сперматозоид состоит из следующих структурных компонентов (рис.16.5). Головка сперматозоида (2—5 мкм в диаметре) содержит ядро, две центриоли и акросому. Ядро занимает большую часть головки сперматозоида и содержит гаплоидный набор хромосом. Акросома головки содержит ионы Са2+ и энзимы, необходимые для проникновения сперматозоида в ооцит. Жгутик (хвост) сперматозоида имеет длину порядка 100 мкм и содержит комплекс микротрубочек, необходимых для движения сперматозоида. Митохондрии продуцируют энергию для сократительных белков, осуществляющих волнообразные движения жгутика сперматозоида
Научная электронная библиотека

Половые железы выполняют инкреторную (стрероидогенез) и экскреторную (сперматогенез) функции. Для гонадотропинов характерен синергизм при регуляции биосинтеза половых стероидов. ФСГ увеличивает число рецепторов к ЛГ на плазматической мембране клеток Лейдига и чувствительность последних к стимулирующему действию ЛГ. Действие ЛГ непрямое, а опосредовано влиянием на сперматогенез тестостерона. Тестостерон, проникая в половые клетки, стимулирует развитие сперматоцитов в сперматиды, которые затем превращаются в сперматозоиды. Центральная регуляция сперматогенеза осуществляется в основном с помощью ФСГ, который действует на сперматогонии и на сперматоциты, способствуя их дифференцировке и созреванию. Установлено, что в течение каждой минуты в организме мужчины вырабатывается 50 000 сперматозоидов. В течение каждого часа его яички вырабатывают 3 000 000 сперматозоидов. В течение каждого дня 72 000 000 сперматозоидов. Созревание половых клеток у взрослого занимает около 72 дней в яичках и 12 дней в придатках, чтобы достигнуть уровня зрелости, то есть в общей сложности почти 3 месяца.
Клетки Сертоли составляют 10–15 % клеточных элементов канальцев. Помимо фагоцитарной активности и регуляции сперматогенеза они вырабатывают ингибин, регулирующий выработку ФСГ, а в эмбриональной жизни – фактор, ингибирующий развитие мюллеровых каналов.
Клетки Сертоли продуцируют андрогенсвязывающие белки (АСБ), которые секретируются в полость семенных канальцев. Синтез АСБ находится под контролем ФСГ. Синтез и секреция АСБ стимулируются также тестостероном. Таким образом, клетки Сертоли реагируют продукцией одного и того же белка (андроген-связывающего) как на ФСГ, так и на тестостерон. Одновременное воздействие обоих гормонов приводит к продукций большого количества андроген-связывающего белка, чем действие каждого из них в отдельности. Эти белки ответственны за транспорт андрогенов внутри канальцев и поддержание оптимальной их концентрации, необходимой для обеспечения метаболических процессов в половых клетках. Позднее открыты кислотный эпидидемальный гликопротеид (КЭГ) и иростатеин (ПТ). Предполагается, что КЭГ способствует созреванию сперматозоидов в придатке. ПТ вырабатывается в вентральной доле предстательной железы. Он присутствует в семенной жидкости и покрывает мембрану сперматозоидов. ПТ способен обеспечивать высокие уровни андрогенов вблизи эпителиальных клеток простаты. Высокая; внутргонадная концентрация тестостерона необходима для успешного осуществления сперматогенеза, хотя в придатке яичка основным метаболитом, оказывающим влияние на сперматогенез, является дигидротестостерон (ДГТ).
Для проявления сперматогенного эффекта ФСГ в организме должны быть андрогены, в частности, тестостерон. Действие андрогенов на сперматогенез своеобразно и во многом сопряжено с механизмом обратной связи системы гипоталамус – гипофиз – семенники. В эксперименте небольшие дозы тестостерона вызывают активацию сперматогенеза в течение 30–45 дней, после чего наступает его угнетение на фоне атрофии интестициальной ткани. Большие дозы тестостерона угнетают сперматогенез и гормональную функцию яичек. В первую очередь подавляется функция клеток Лейдига. Угнетение сперматогенеза происходит на стадии сперматоцитов с понижением числа митозов и прогрессированием дегенеративных процессов в семенных канальцах. ЯГ регулирует секрецию тестостерона. В организме мужчин основной его мишенью являются интерстициальные клетки Лейдига. ЛГ называют также гормоном, стимулирующим интерстициальные клетки (ГСИК). ФСГ дает морфогенетический эффект, а именно пролиферацию клеток Сертоли и сперматогенного эпителия, что необходимо для последующей активации сперматогенеза. Для проявления эффекта ФСГ необходимо присутствие небольшого количества ЛГ и тестостерона в яичках. Наряду с гонадотропинами важное значение имеет действие на половую систему другого гормона аденогипофиза – пролактина (ПЛ). Он значительно усиливает действие ЛГ на стероидогенез в клетках Лейдига, контролирует уровень предшественников тестостерона, влияя тем самым на его продукцию. Пролактин оказывает модулирующее воздействие на механизмы отрицательного обратного влияния тестостерона на тестикулярном и центральном уровнях. ПЛ также увеличиваег количество рецепторов андрогенов в тканях предстательной железы и семенных пузырьков, тем самым стимулирует их рост за счет усиления действия стероидных гормонов в этом направлении. С другой стороны, между секрецией ПЛ и уровнем гонадотропных гормонов существуют реципрокные отношения – повышение уровня ПЛ сопровождается снижением секреции ЛГ, что отражается и на продукции половых гормонов. Дефицит или избыток ПЛ часто сопровождается снижением репродуктивной функции.
Кроме центральной, существует и местная (внутритестикулярная) регуляция сперматогенеза, осуществляющаяся за счет факторов, вырабатываемых в основном клетками Сертоли. Они синтезируют эсградиол, ингибирующий секрецию клетками Лейдига тестостерона. В семенниках эстрогены из клеток Сертоли регулируют биосинтез тестостерона клетками Лейдига. В клетках Сертоли семенниками вырабатывается тестикулярный гонадолиберин, который оказывает паракринное действие на клетки Лейдига в семеннике, активируя их к продукции и секреции тест остерона.
Учитывая, что введение тестостерона не влияет на секрецию ФСГ, а для подавления ее необходимы дозы эстрогенов выше физиологических, исследователи пришли к заключению о непричастности этих гормонов к регуляции ФСГ. Данную функцию выполняет инГибин. Образование ингибина клетками Сертоли стимулируется андрогенами. Поддержание количественно нормальной секреции ингибина требует совместного действия обоих гонадотропинов. Обнаружение ингибина в изолированных клетках гипофиза привело к выводу, что он оказывает прямое действие на гипофиз, в частности, угнетает секрецию ФСГ. В норме существует отрицательная обратная связь между ФСГ и ингибином, секретируемым клетками Сертоли в ответ на ФСГ. При воздействии тестостерона снижается уровень ингибина и подавляется процесс сперматогенеза. Около 30 % секретируемого ингибина не зависит от гонадотропинов. Циркулирующий в сыворотке крови ингибин В служит маркером функционирования клеток Сертоли. Важная физиологическая роль ингибина заключается в контроле секреции ФСГ по принципу обратной связи! Клетки Сертоли выполняют еще ряд важных функций, к которым относится: синтез фактора роста семенных канальцев, способность фагоцитировать остатки продуктов сперматогенеза, формирование и поддержание целостности гемато-тестикулярного барьера. Полагают* что инсулиноподобный фактор роста-1 принимает участие в регуляции дйфференцировки сперматогоний и сперматоцитов, клеток Лейдига и самих клеток Сертоли. Клетки Сертоли секретируют также ряд других белков – трансферрин, церулоплазмин, соматомедин-подобное вещество, активатор плазминогена и др. Регуляция функций семенников осуществляется также паракринным путем. Клетки Сертоли регулируют размножение и созревание зародышевых клеток, которые. в свою очередь, циклически регулируют функцию клеток Сертоли, а через них и клеток Лейдига. Секреция тестикулярных андрогенов регулируется ЛГ и ФСГ.
Читайте также:
- Осложнения катетеризации центральных вен у новорожденного
- Как выполнить блокаду подглазничного нерва, интраорально
- Ошибки, осложнения спинальной и эпидуральной анестезии
- Паралич лицевого нерва - паралич Белла. Токсичные для ушей лекарства
- Врожденное расщепление позвоночника. Причины расщепления позвоночника эмбриона