Иннервация сердца. Хронотропный эффект. Дромотропный эффект. Инотропный эффект. Батмотропный эффект.
Добавил пользователь Владимир З. Обновлено: 22.01.2026
Работа сердца регулируется двумя нервами: блуждающим (или вагусом), относящимся к парасимпатической нервной системе, и симпатическим. Эти нервы образованы двумя нейронами. Тела первых нейронов, отростки которых составляют блуждающий нерв, расположены в продолговатом мозге. Отростки этих нейронов заканчиваются в инграмуральных ганглиях сердца. Здесь находятся вторые нейроны, отростки которых идут к проводящей системе, миокарду и коронарным сосудам.
Первые нейроны симпатической нервной системы, регулирующей работу сердца, лежат в боковых рогах I-V грудных сегментов спинного мозга. Отростки этих нейронов заканчиваются в шейных и верхних грудных симпатических узлах. В этих узлах находятся вторые нейроны, отростки которых идут к сердцу. Большая часть симпатических нервных волокон направляются к сердцу от звездчатого ганглия. Нервы, идущие от правого симпатического ствола, в основном подходят к синусному узлу и к мышцам предсердий, а нервы левой стороны — к атриовентрикулярному узлу и мышцам желудочков
Характеристика хронотропного, инотропного, батмотропного, дромотропного и клинотропного регуляторных эффектов на сердце.
хронотропный (изменяют частоту сердечных сокращений);
инотропный (изменяют силу сердечных сокращений);
батмотропный (влияют на возбудимость миокарда);
дромотропный (влияет на проводимость);
тонотропный (влияют на тонус миокарда);
Сердце — обильно иннервированный орган. Среди чувствительных образований сердца основное значение имеют две популяции механорецепторов, сосредоточенных, главным образом, в предсердиях и левом желудочке: А-рецепторы реагируют на изменение напряжения сердечной стенки, а В-рецепторы возбуждаются при ее пассивном растяжении. Афферентные волокна, связанные с этими рецепторами, идут в составе блуждающих нервов. Свободные чувствительные нервные окончания, расположенные непосредственно под эндокардом, представляют собой терминали афферентных волокон, проходящих в составе симпатических нервов. Эфферентная иннервация сердца осуществляется при участии обоих отделов вегетативной нервной системы. Тела симпатических преганглионарных нейронов, участвующих в иннервации сердца, располагаются в сером веществе боковых рогов трех верхних грудных сегментов спинного мозга. Преганглионарные волокна направляются к нейронам верхнего грудного (звездчатого) симпатического ганглия. Постганглионар-ные волокна этих нейронов вместе с парасимпатическими волокнами блуждающего нерва образуют верхний, средний и нижний сердечные нервы, Симпатические волокна пронизывают весь орган и иннервируют не только миокард, но и элементы проводящей системы. Тела парасимпатических преганглионарных нейронов, участвующих в иннервации сердца, располагаются в
продолговатом мозге. Их аксоны идут в составе блуждающих нервов. После вхождения блуждающего нерва в грудную полость от него отходят веточки, которые включаются в состав сердечных нервов. Отростки блуждающего нерва, проходящие в составе сердечных нервов, представляют собой парасимпатические преганглионарные волокна. С них возбуждение передается на интрамуральные нейроны и далее — преимущественно на элементы проводящей системы. Влияния, опосредованные правым блуждающим нервом, адресованы, в основном, клеткам синоатриального, а левым — клеткам атриовентрикулярного узла. Прямого влияния на
желудочки сердца блуждающие нервы не оказывают. Иннервируя ткань водителей ритма, вегетативные нервы способны менять их возбудимость, тем самым вызывая изменения частоты генерации потенциалов действия и сокращений сердца (хронотропный эффект). Нервные влияния изменяют скорость электротонической передачи возбуждения и, следовательно, длительности фаз сердечного цикла. Такие эффекты называют дромотропными. Поскольку действие медиаторов вегетативной нервной системы заключается в изменении уровня циклических нуклеотидов и энергетического обмена, вегетативные нервы в целом способны влиять и на силу сердечных сокращений (инотропный эффект). В лабораторных условиях получен эффект изменения величины порога возбуждения кардиомиоцитов под действием нейромедиаторов, его обозначают как батмотропный. Перечисленные пути воздействия нервной системы на сократительную активность миокарда и насосную функцию сердца представляют собой хотя и исключительно важные, но вторичные по отношению к миогенным механизмам модулирующие влияния.
40.Экстракардиальные нервы сердца.Характеристика хронотропного, инотропного, батмотропного, дромотропного и клинотропного регуляторных эффектов на сердце.
Иннервация сердца. Хронотропный эффект. Дромотропный эффект. Инотропный эффект. Батмотропный эффект.
Сердце — обильно иннервированный орган. Среди чувствительных образований сердца основное значение имеют две популяции механорецепторов, сосредоточенных, главным образом, в предсердиях и левом желудочке: А-рецепторы реагируют на изменение напряжения сердечной стенки, а В-рецепторы возбуждаются при ее пассивном растяжении. Афферентные волокна, связанные с этими рецепторами, идут в составе блуждающих нервов. Свободные чувствительные нервные окончания, расположенные непосредственно под эндокардом, представляют собой терминали афферентных волокон, проходящих в составе симпатических нервов.
Эфферентная иннервация сердца осуществляется при участии обоих отделов вегетативной нервной системы. Тела симпатических преганглионарных нейронов, участвующих в иннервации сердца, располагаются в сером веществе боковых рогов трех верхних грудных сегментов спинного мозга. Преганглионарные волокна направляются к нейронам верхнего грудного (звездчатого) симпатического ганглия. Постганглионар-ные волокна этих нейронов вместе с парасимпатическими волокнами блуждающего нерва образуют верхний, средний и нижний сердечные нервы, Симпатические волокна пронизывают весь орган и иннервируют не только миокард, но и элементы проводящей системы.
Тела парасимпатических преганглионарных нейронов, участвующих в иннервации сердца, располагаются в продолговатом мозге. Их аксоны идут в составе блуждающих нервов. После вхождения блуждающего нерва в грудную полость от него отходят веточки, которые включаются в состав сердечных нервов.
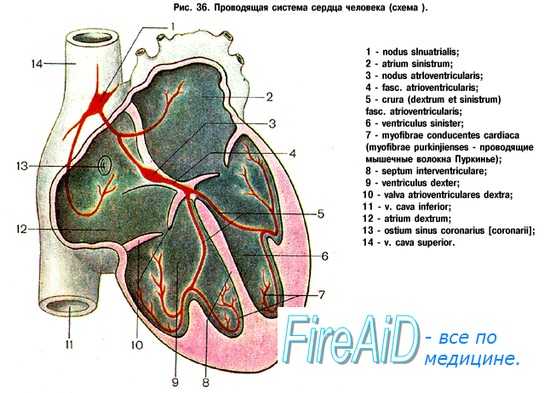
Отростки блуждающего нерва, проходящие в составе сердечных нервов, представляют собой парасимпатические преганглионарные волокна. С них возбуждение передается на интрамуральные нейроны и далее — преимущественно на элементы проводящей системы. Влияния, опосредованные правым блуждающим нервом, адресованы, в основном, клеткам синоатриального, а левым — клеткам атриовентрикулярного узла. Прямого влияния на желудочки сердца блуждающие нервы не оказывают.
Иннервируя ткань водителей ритма, вегетативные нервы способны менять их возбудимость, тем самым вызывая изменения частоты генерации потенциалов действия и сокращений сердца (хронотропный эффект). Нервные влияния изменяют скорость электротонической передачи возбуждения и, следовательно, длительности фаз сердечного цикла. Такие эффекты называют дромотропными.
Поскольку действие медиаторов вегетативной нервной системы заключается в изменении уровня циклических нуклеотидов и энергетического обмена, вегетативные нервы в целом способны влиять и на силу сердечных сокращений (инотропный эффект). В лабораторных условиях получен эффект изменения величины порога возбуждения кардиомиоцитов под действием нейромедиаторов, его обозначают как батмотропный.
Перечисленные пути воздействия нервной системы на сократительную активность миокарда и насосную функцию сердца представляют собой хотя и исключительно важные, но вторичные по отношению к миогенным механизмам модулирующие влияния.
Парасимпатические воздействия на сердце. Влияние на сердце блуждающего нерва. Вагусные воздействия на сердце.
Результатом стимуляции этих нервов является отрицательный хронотропный эффект сердца (рис. 9.17), на фоне которого проявляются также отрицательные и дромотропный инотропный эффекты. Существуют постоянные тонические влияния на сердце со стороны бульбарных ядер блуждающего нерва: при его двусторонней перерезке частота сердцебиений возрастает 1,5—2,5 раза. При длительном сильном раздражении влияние блуждающих нервов на сердце постепенно ослабевает или прекращается, что получило название «эффекта ускользания» сердца из-под влияния блуждающего нерва.
Различные отделы сердца по-разному реагируют на возбуждение парасимпатических нервов. Так, холинергические влияния на предсердия вызывают значительное угнетение автоматии клеток синусного узла и спонтанно возбудимой ткани предсердий. Сократимость рабочего миокарда предсердий в ответ на стимуляцию блуждающего нерва снижается. Рефрактерный период предсердий при этом также уменьшается в результате значительного укорочения длительности потенциала действия предсердных кардиомиоцитов. С другой стороны, рефрактерность кардиомиоцитов желудочков под влиянием блуждающего нерва, напротив, значительно возрастает, а отрицательный парасимпатический инотропный эффект на желудочки выражен в меньшей степени, чем на предсердия.
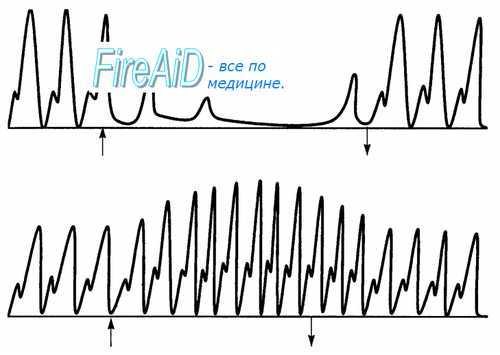
Рис. 9.17. Электрическое раздражение эфферентных нервов сердца. Вверху — уменьшение частоты сокращений при раздражении блуждающего нерва; внизу-увеличение частоты и силы сокращений при раздражении симпатического нерва. Стрелками отмечены начало и конец раздражения.
Электрическая стимуляция блуждающего нерва вызывает урежение или прекращение сердечной деятельности вследствие торможения автоматической функции водителей ритма синоатриального узла. Выраженность этого эффекта зависит от силы и частоты раздражения блуждающего нерва. По мере увеличения силы раздражения отмечается переход от небольшого замедления синусного ритма до полной остановки сердца.
Отрицательный хронотропный эффект раздражения блуждающего нерва связан с угнетением (замедлением) генерации импульсов в водителе ритма сердца синусного узла. Поскольку при раздражении блуждающего нерва в его окончаниях выделяется медиатор — ацетилхолин, при его взаимодействии с мускариночувствительными рецепторами сердца повышается проницаемость поверхностной мембраны клеток водителей ритма для ионов калия. Вследствие этого возникает гиперполяризация мембраны, которая замедляет (подавляет) развитие медленной спонтанной диастолической деполяризации, и поэтому мембранный потенциал позже достигает критического уровня. Это приводит к урежению ритма сокращений сердца.
При сильных раздражениях блуждающего нерва диастолическая деполяризация подавляется, возникают гиперполяризация водителя ритма и полная остановка сердца. Развитие гиперполяризации в клетках водителей ритма снижает их возбудимость, затрудняет возникновение очередного автоматического потенциала действия и тем самым приводит к замедлению или даже остановке сердца. Стимуляция блуждающего нерва, усиливая выход калия из клетки, увеличивает мембранный потенциал, ускоряет процесс реполяризации и при достаточной силе раздражающего тока укорачивает длительность потенциала действия клеток водителя ритма.
При вагусных воздействиях имеет место уменьшение амплитуды и длительности потенциала действия кардиомиоцитов предсердия. Отрицательный инотропный эффект связан с тем, что уменьшенный по амплитуде и укороченный потенциал действия не способен возбудить достаточное количество кардиомиоцитов. Кроме того, вызванное ацетилхолином повышение калиевой проводимости противодействует потенциалзависимому входящему току кальция и проникновению его ионов внутрь кардиомиоцита. Холинергический медиатор ацетилхолин может также угнетать АТФ-азную активность миозина и, таким образом, уменьшать величину сократимости кардиомиоцитов. Возбуждение блуждающего нерва приводит к повышению порога раздражения предсердий, подавлению автоматии и замедлению проводимости атриовентрикулярного узла. Указанное замедление проводимости при холинергических влияниях может вызвать частичную или полную атриовентрикулярную блокаду.
Возбуждение миокарда. Сокращение миокарда. Сопряжение возбуждения и сокращения миокарда.
Инициатором сокращения миокарда, как и в скелетной мышце, является потенциал действия, распространяющийся вдоль поверхностной мембраны кардиомиоцита. Поверхностная мембрана волокон миокарда образует впячивания, так называемые поперечные трубочки (Т-система), к которым примыкают продольные трубочки (цистерны) саркоплазматического рети-кулума, являющиеся внутриклеточным резервуаром кальция (рис. 9.10). Саркоплазматический ретикулум в миокарде выражен в меньшей степени, чем в скелетной мышце. Нередко к поперечной Т-трубочке примыкают не две продольные трубочки, а одна (система диад, а не триад, как в скелетной мышце). Считается, что потенциал действия распространяется с поверхностной мембраны кардиомиоцита вдоль Т-трубочки в глубь волокна и вызывает деполяризацию цистерны саркоплазматического ретикулума, что приводит к освобождению из цистерны ионов кальция.
Следующим этапом электромеханического сопряжения является перемещение ионов кальция к сократительным протофибриллам. Сократительная система сердца представлена сократительными белками — актином и миозином, и модуляторными белками — тропомиозином и тропонином. Молекулы миозина формируют толстые нити саркомера, молекулы актина—тонкие нити. В состоянии диастолы тонкие актиновые нити входят своими концами в промежутки между толстыми и более короткими миози-новыми нитями. На толстых нитях миозина располагаются поперечные мостики, содержащие АТФ, а на нитях актина — модуляторные белки — тропомиозин и тропонин. Эти белки образуют единый комплекс, блокирующий активные центры актина, предназначенные для связывания миозина и стимуляции его АТФазной активности.
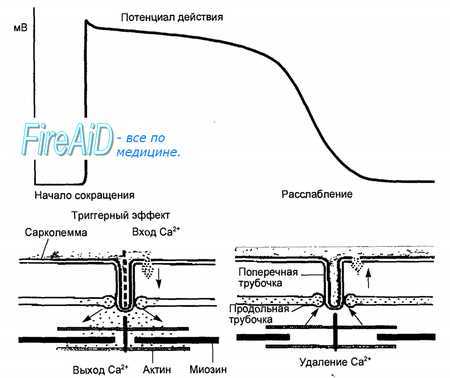
Рис.Рис. 9.10. Схема соотношений между возбуждением, током Са2+ и активацией сократительного аппарата. Начало сокращения связано с выходом Са2+ из продольных трубочек при деполяризации мембраны. Са2+, входящий через мембраны кардио-миоцита в фазу плато потенциала действия, пополняет запасы Са2+ в продольных трубочках.
Сокращение волокон миокарда начинается со связывания тропонином вышедшего из саркоплазматического ретикулюма в межфибриллярное пространство кальция. Связывание кальция вызывает изменения конформации тропонин-тропомиози-нового комплекса. В результате этого открываются активные центры и происходит взаимодействие актиновых и миозиновых нитей. При этом стимулируется АТФазная активность миозиновых мостиков, происходит распад АТФ и выделяющаяся энергия используется на скольжение нитей друг относительно друга, приводящее к сокращению миофибрилл. В отсутствие ионов кальция тропонин препятствует образованию актомиозиново-го комплекса и усилению АТФазной активности миозина. Морфологические и функциональные особенности миокарда свидетельствуют о тесной связи между внутриклеточным депо кальция и внутриклеточной средой. Так как запасы кальция во внутриклеточных депо невелики, большое значение имеет вход кальция в клетку во время генерации потенциала действия (см. рис. 9.10). Потенциал действия и сокращение миокарда совпадают по времени. Поступление кальция из наружной среды в клетку создает условия для регуляции силы сокращения миокарда. Большая часть входящего в клетку кальция, очевидно, пополняет его запасы в цистернах сарко-плазматического ретикулума, обеспечивая последующие сокращения.
Удаление кальция из клеточного пространства приводит к разобщению процессов возбуждения и сокращения миокарда. Потенциалы действия при этом регистрируются почти в неизменном виде, но сокращения миокарда не происходит. Вещества, блокирующие вход кальция во время генерации потенциала действия, вызывают аналогичный эффект. Вещества, угнетающие кальциевый ток, уменьшают длительность фазы плато и потенциала действия и понижают способность миокарда к сокращению. При повышении содержания кальция в межклеточной среде и при введении веществ, усиливающих вход этого иона в клетку, сила сердечных сокращений увеличивается. Таким образом, потенциал действия играет роль пускового механизма, вызывая освобождение кальция из цистерн саркоплаз-матического ретикулума, регулирует сократимость миокарда, а также пополняет запасы кальция во внутриклеточных депо.
Читайте также: