Медиатор пресинаптической мембраны. Постсинаптическая мембрана
Добавил пользователь Евгений Кузнецов Обновлено: 27.01.2026
В ЦНС нервные клетки связаны друг с другом посредством синапсов. Синапс – это структурно функциональное образование, которое обеспечивает передачу возбуждения или торможения с нервного волокна на иннервируемую клетку.
Синапсы по локализации делятся на центральные (расположены в пределах ЦНС, а также в ганглиях вегетативной нервной системы) и периферические (расположены вне ЦНС, обеспечивают связь с клетками иннервируемой ткани).
В функциональном отношении синапсы делятся на возбуждающие, в которых в результате деполяризации постсинаптической мембраны генерируется возбуждающий постсинаптический потенциал, и тормозные, в пресинаптических окончаниях которых выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала.
По механизму передачи синапсы делятся на химические и электрические. Химические синапсы передают возбуждение или торможение за счет особых веществ – медиаторов. В зависимости от вида медиатора химические синапсы подразделяются на:
1. холинергические (медиатор – ацетилхолин)
2. адренергические (медиаторы – адреналин, норадреналин)
По анатомической классификации синапсы делятся на нейросекреторные, нервно-мышечные и межнейронные.
Синапс состоит из трех основных компонентов:
1. пресинаптической мембраны
2. постсинаптической мембраны
3. синаптической щели
Пресинаптическая мембрана является окончанием отростка нервной клетки. Внутри отростка в непосредственной близости от мембраны имеется скопление пузырьков (гранул), содержащих тот или иной медиатор. Пузырьки находятся в постоянном движении.
Постсинаптическая мембрана является частью клеточной мембраны иннервируемой ткани. Постсинаптическая мембрана в отличие от пресинаптической имеет белковые хеморецепторы к биологически активным (медиаторам, гормонам), лекарственным и токсическим веществам. Важная особенность рецепторов постсинаптической мембраны – их химическая специфичность, т.е. способность вступать в биохимическое взаимодействие только с определенным видом медиатора.
Синаптическая щель представляет собой пространство между пре- и постсинаптичекой мембранами, заполненное жидкостью, близкой по составу к плазме крови. Через нее медиатор медленно диффундирует от пресинаптической мембраны к постсинаптической.
Особенности строения нервно-мышечного синапса обусловливают его физиологические свойства.
1. Одностороннее проведение возбуждения (от пре- к постсинаптической мембране), обусловленное наличием чувствительных к медиатору рецепторов только в постсинаптической мембране.
2. Синаптическая задержка проведения возбуждения (время между приходом импульса в пресинаптическое окончание и началом постсинаптического ответа), связанная с малой скоростью диффузии медиатора в синаптическую щель по сравнению со скоростью прохождения импульса по нервному волокну.
3. Низкая лабильность и высокая утомляемость синапса, обусловленная временем распространения предыдущего импульса и наличием у него периода абсолютной рефрактерности.
4. Высокая избирательная чувствительность синапса к химическим веществам, обусловленная специфичностью хеморецепторов постсинаптической мембраны.
Этапы синаптической передачи.
1. Синтез медиатора. В цитоплазме нейронов и нервных окончаний синтезируются химические медиаторы – биологически активные вещества. Они синтезируются постоянно и депонируются в синаптических пузырьках нервных окончаний.
2. Секреция медиатора. Высвобождение медиатора из синаптических пузырьков имеет квантовый характер. В состоянии покоя оно незначительно, а под влиянием нервного импульса резко усиливается.
3. Взаимодействие медиатора с рецепторами постсинаптической мембраны. Это взаимодействие заключается в избирательном изменении проницаемости ионоселективных каналов эффекторной клетки в области активных центров связывания с медиатором. Взаимодействие медиатора со своими рецепторами может вызвать возбуждение или торможение нейрона, сокращение мышечной клетки, образование и выделение гормонов секреторными клетками. В случае увеличения проницаемости натриевых и кальциевых каналов усиливается поступление Na и Ca в клетку с последующей деполяризацией мембраны, возникновением ПД и дальнейшей передачей нервного импульса. Такие синапсы называются возбуждающими. Если повышается проницаемость калиевых каналов и каналов для хлора, наблюдается избыточный выход К из клетки с одновременной диффузией в нее Cl , что приводит к гиперполяризации мембраны, снижению ее возбудимости и развитию тормозных постсинаптических потенциалов. Передача нервных импульсов затрудняется или совсем прекращается. Такие синапсы называются тормозными.
Рецепторы, взаимодействующие с АХ, называются холинорецепторами. В функциональном отношении они разделяются на две группы: М - и Н-холинорецепторы. В синапсах скелетных мышц присутствуют только Н-холинорецепторы, тогда как в мышцах внутренних органов – преимущественно М-холинорецепторы.
Рецепторы, взаимодействующие с НА, называются адренорецепторами. В функциональном отношении они делятся на альфа- и бета-адренорецепторы. В постсинаптической мембране гладкомышечных клеток внутренних органов и кровеносных сосудов часто соседствуют оба вида адренорецепторов. Действие НА является деполяризующим, если он взаимодействует с альфа-адренорецепторами (сокращение мышечной оболочки стенок кровеносных сосудов или кишечника), или тормозным – при взаимодействии с бета-адренорецепторами (их расслабление).
4. Инактивация медиатора. Инактивация (полная потеря активности) медиатора необходима для реполяризации постсинаптической мембраны и восстановления исходного уровня МП. Наиболее важным путем инактивации является гидролитическое расщепление с помощью ингибиторов. Для АХ ингибитором является холинэстераза, для НА и адреналина – моноаминоксидаза и катехолоксиметилтрансфераза.
Другой путь удаления медиатора из синаптической щели – «обратный захват» его пресинаптическими окончаниями (пиноцитоз) и обратный аксонный транспорт, особенно выраженный для катехоламинов.
В основе координационной деятельности ЦНС лежит взаимодействие процессов возбуждения и торможения.
Возбуждение – это активный процесс, представляющий собой ответную реакцию ткани на раздражение и характеризующийся повышением функций ткани.
Торможение – это активный процесс, представляющий собой ответную реакцию ткани на раздражение и характеризующийся снижением функций ткани.
Первичное торможение в ЦНС возникает за счет тормозных нейронов. Это особый вид вставочных нейронов, которые при передаче импульса выделяют тормозной медиатор. Различают 2 вида первичного торможения: постсинаптическое и пресинаптическое.
Постсинаптическое торможение возникает, если аксон тормозного нейрона образует синапс с телом нейрона и, выделяя медиатор, вызывает гиперполяризацию клеточной мембраны, тормозя активность клетки.
Пресинаптическое торможение возникает, когда аксон тормозного нейрона образует синапс с аксоном возбуждающего нейрона, препятствуя проведению импульса.
Медиатор пресинаптической мембраны. Постсинаптическая мембрана
Медиатор пресинаптической мембраны. Постсинаптическая мембрана
Мембрану пресинаптической терминали называют пресинаптической мембраной. Она содержит большое количество электроуправляемых кальциевых каналов. Когда потенциал действия деполяризует пресинаптическую мембрану, эти кальциевые каналы открываются, что позволяет большому числу ионов кальция войти в терминалы Количество медиатора, которое затем выделится из терминали в синаптическую щель, находится в прямой зависимости от числа вошедших ионов кальция.
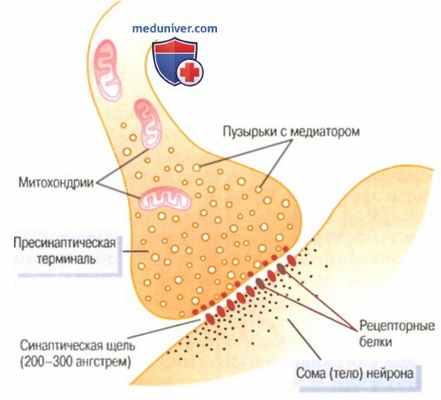
Физиологическая анатомия синапса
Точный механизм, с помощью которого ионы кальция вызывают выделение медиатора, не известен, но предполагают следующее.
Считают, что ионы кальция, войдя в пресинаптическую терминаль, связываются с особыми белковыми молекулами на внутренней поверхности пресинаптической мембраны, которые называют местами освобождения. Это связывание вызывает открытие мест освобождения медиатора сквозь мембрану, в результате после каждого одиночного потенциала действия несколько пузырьков выделяют свой медиатор в щель.
В каждом из пузырьков, содержащих медиатор ацетилхолин, накапливается от 2000 до 10000 его молекул, а общего числа пузырьков в пресинаптической терминали достаточно, чтобы проводить от нескольких сотен до более 10000 потенциалов действия.
а) Действие медиатора на постсинаптический нейрон - функция рецепторных белков. Мембрана постсинаптического нейрона содержит большое количество рецепторных белков (для облегчения понимания просим вас изучить рисунок ниже).
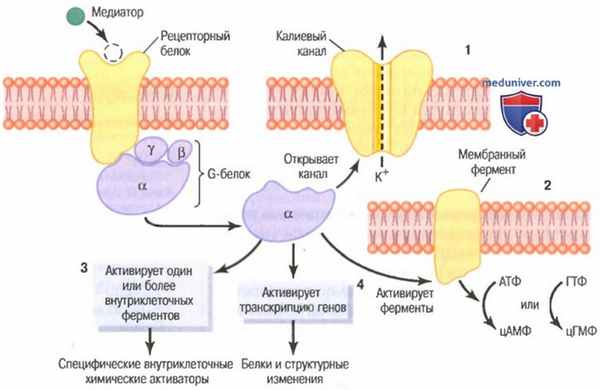
Система вторичного посредника, посредством которого медиатор, выделяемый первым нейроном, может активировать второй нейрон сначала посредством освобождения G белка в цитоплазму второго нейрона. Показаны четыре из последующих возможных эффектов G белка:
1 - открытие ионного канала в мембране второго нейрона,
2 - активация фер ментнои системы в мембране нейрона,
3 - активация внутриклеточной ферментной системы и/или
4 - изменение генной транскрипции во втором нейроне
Молекулы этих рецепторов имеют два важных компонента:
(1) связывающий компонент, который выдается наружу от постсинаптической мембраны в синаптическую щель, где он связывает медиатор, выделившийся из пресинаптической терминали;
(2) ионофорный компонент, проходящий сквозь постсинаптическую мембрану внутрь постсинаптического нейрона.
Ионофор, в свою очередь, может быть одним из двух типов:
(1) ионным каналом, позволяющим определенным ионам проходить через этот канал;
(2) активатором вторичного посредника, представляющим собой молекулу, которая выдается внутрь клетки в ее цитоплазму и активирует одно или более веществ внутри постсинаптического нейрона.
Эти вещества, в свою очередь, служат вторичными посредниками для усиления или угнетения специфических клеточных функций.
б) Ионные каналы. Ионные каналы в мембране постсинаптического нейрона обычно бывают двух типов:
(1) катионные каналы, которые в открытом состоянии обычно пропускают ионы Na+, но иногда также ионы К+ и/или Са 2+ ;
(2) анионные каналы, пропускающие главным образом ионы Cl- и минимальные количества других анионов.
Катионные каналы, проводящие ионы Na+, выстланы отрицательными зарядами. Эти заряды затягивают положительно заряженные ионы Na+ в канал, если диаметр канала превышает размер гидратированного иона Na+. Однако те же самые отрицательные заряды отталкивают ионы Сl- и другие анионы, препятствуя их прохождению.
Если диаметры анионных каналов достаточно велики, ионы Сl- входят в них и проходят на противоположную сторону, тогда как вход натриевых, калиевых и кальциевых катионов блокируется в основном из-за слишком больших размеров их гидратированных ионов.
Далее мы узнаем, что процесс открытия катионных каналов, позволяющий войти в клетку положительно заряженным ионам Na+, способствует возбуждению нейрона, поэтому медиаторы, ведущие к открытию катионных каналов, называют возбуждающими. И наоборот, процесс открытия анионных каналов, позволяющий войти отрицательным электрическим зарядам, тормозит нейрон. Соответственно, медиаторы, открывающие эти каналы, называют тормозными.
При активации медиатором ионного канала он обычно открывается в течение долей миллисекунды; после удаления медиатора канал закрывается так же быстро. Такое открытие и закрытие каналов обеспечивает очень быстрое управление постсинаптическими нейронами.
Видео физиология синапса и нерва - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Медиатор пресинаптической мембраны. Постсинаптическая мембрана
Ткани и органы. Нервная ткань
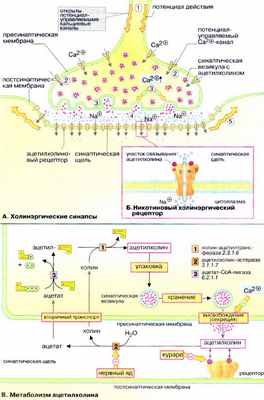
А. Холинэргические синапсы
Передача сигналов между нейронами и от нейронов к мышечным клеткам (так называемая нейронейрональная и нейромышечная трансдукция) происходит в нервных окончаниях (синапсах). С помощью сигнальных веществ, медиаторов. Синапсы образованы мембранами двух контактирующих клеток, пресинаптической и постсинаптической которые разделены узкой синаптической щелью. Медиатор выделяется в синаптическую щель за счет зкзоцитоза, диффундирует к рецепторам постсинаптической мембраны, связывается с ними и передает сигнал соседней клетке. Белки-рецепторы — это лиганд-активируемые ионные каналы (см. рис. 341) либо мембранные белки, которые управляют ионными каналами посредством G-белков (см. рис. 373).
Ацетилхолин — нейромедиатор моторной концевой пластинки. Ацетилхолиновые рецепторы (никотиновый и мускариновый) — это лиганд-активируемые ионные каналы, которые открываются для прохождения ионов Na + и К + . Никотиновые рецепторы (быстрые) локализованы главным образом в месте контакта аксонов со скелетными мышцами. Мускариновые рецепторы (медленные) локализованы в головном мозге, секреторных клетках, гладких и сердечных мышцах.
Процесс передачи сигнала включает следующие этапы. Потенциал действия достигает пресинаптической мембраны ( 1 ). Это вызывает открывание потенциал-управля емых Сa 2+ -каналов ( 2 ). Ионы Са 2+ проникают из внеклеточного пространства в клетку, их уровень в синапсе резко увеличивается, что инициирует процесс экзоцитоза. Синаптические везикулы выделяют содержимое (ацетилхолин) в синаптическую щель ( 3 ). Молекулы ацетилхолина диффундируют через синаптическую щель, связываются с постсинаптическими рецепторами и активируют их ( 4 ). Поток ионов Na + изменяет потенциал покоя постсинаптической мембраны нервной или мышечной клетки настолько, что открываются соседние потенциал-управляемые Na + каналы и возникает потенциал действия ( 5 , см. рис. 341).
Б. Никотиновый холинэргический рецептор
Наиболее детально изучен рецептор ацетилхолина, активируемый никотином. Это трансмембранный комплекс из пяти субъединиц (α 2 βδγ, 250-270 кДа), образующий лиганд-активируемый (хемовозбудимый) ионный канал, проницаемый для ионов Na + и К + . Участки связывания ацетилхолина локализованы на внеклеточной части α-субъединиц. При связывании лиганда в центре молекулы формируется трансмембранный канал, входное отверстие которого имеет форму воронки диаметром около 2 нм. Предполагается, что в формировании канала принимают участие все пять субъединиц. Канал открывается на короткое время для прохождения ионов Na + и К + . Считается, что открывание и закрывание канала происходит в результате аллостерических изменений в заряженных участках полипептидных цепей молекулы рецептора.
Рецептор может связывать различные лекарственные вещества: например, никотин действует как агонист ацетилхолина.
В. Метаболизм ацетилхолина
Ацетилхолин , уксуснокислый эфир холина, образуется в цитоплазме аксонов из ацетил-КоА и холина [ 1 ]. Нейромедиатор хранится в синаптических везикулах , в каждой везикуле содержится примерно 1000-10000 молекул ацетилхолина. После выделения из везикул ацетилхолин попадает в синаптическую щель. Избыток ацетилхолина расщепляется ацетилхолин-эстеразой [ 2 ]. Этот фермент имеет высокое число оборотов, что гарантирует быстрое удаление сигнального вещества. Продукты гидролиза, холин и уксусная кислота , активно захватываются пресинаптической частью синапса и используются для повторного синтеза ацетилхолина [ 3 ].
Соединения, блокирующие остаток серина в активном центре ацетилхолин-эстеразы [ 2 ], например токсин Е605, пролонгируют действие ацетилхолина и действуют как нейротоксины. Напротив, D-тубокурарин ( яд кураре, которым индейцы пропитывали наконечники стрел) является конкурентным ингибитором ацетилхолина при связывании с рецептором.
3. Взаимодействие медиатора с рецепторами постсинаптической мембраны.
Медиатор достигший постсинаптической мембраны действует на ее хеморецепторы, связанные с различными хемо-управляемыми ионными каналами, вследствие чего изменяется проницаемость постсинаптической мембраны для различных ионов (Na + , K + , Cl - ). В зависимости от иона и направления его движения постсинаптическая мембрана деполяризуется или гиперполяризуется и на ней возникает возбуждающий или тормозной потенциал.
Отметим, что электроуправляемых каналов на постсинаптической мембране нет, поэтому сама к генерации ПД она не способна.
Возбуждающий постсинаптический потенциал (ВПСП), возникающий в результате поступления в клетку ионов Na+, электротонически распространяется за пределы постсинаптической мембраны и если его величина достаточно велика, деполяризует уже плазматическую мембрану до критического уровня. В мышечных клетках плазматическая мембрана сразу за пределами синапса имеет потенциал-чувствительные каналы, на ней относительно легко возникает полноценный потенциал действия. В нервных клетках возникновение возбуждения возможно только в аксоном холмике, поэтому активности одного из синапсов, которые, как правило, расположены далеко от него недостаточно для возникновения ПД. Нервная клетка воспринимает (суммирует) несколько синаптических сигналов и генерирует потенциал действия избирательно. В этом заключается интегративная функция нейрона.
Тормозной постсинаптический (ТПСП) потенциал возникает при усилении проницаемости для калия и хлора, он гиперполяризует постсинаптическую мембрану, препятствуя электротоническому распространению возбуждения по мембране нервной клетки. Процесс торможения имеет место только на межнейронных синапсах.
Принципы работы возбуждающего и тормозного синапсов сходны. Разница заключается в том с какими специфическими каналами связаны рецепторы расположенные на постсинаптической мембране, от чего и зависит физиологическая реакция.
4. Инактивация медиатора
Инактивация медиатора - последняя стадия синаптической передачи, обеспечивающая ее прекращение. Она необходима для реполяризации постсинаптической мембраны и восстановления исходного уровня ее мембранного потенциала, что обеспечивает возможность восприятия следующего сигнала. Наиболее распространенным является гидролитическое расщепление медиатора с помощью соответствующих ферментов, постоянно находящихся в синаптической щели. Для ацетилхолина, например, таким ферментом является холинэстераза, для норадреналина - моноаминооксидаза. Эти ферменты начинают разрушать медиатор на исходные компоненты, сразу, как только он попадает в синаптическую щель, и скорость разрушения настолько велика, что при небольших количествах медиатора, он разрушается еще до того, как достигает постсинаптической мембраны.
Продукты расщепления либо поступают в кровь и циркулируют как его предшественники, либо за счет пиноцитоза осуществляется их «обратное всасывание» в синаптическое окончание, где происходит их восстановление и упаковка в синаптические пузырьки и медиатор может использоваться повторно.
Строение синапса и его медиаторы. Виды синапсов
Возбуждение с нервного волокна на нервную, мышечную и железистую клетку передается посредством специального структурного образования — синапса.
Синапс — структура, обеспечивающая проведение сигнала от одной клетки к другой. Термин был введен английским физиологом Ч. Шеррингтоном в 1897 г.
Строение синапса
Синапсы состоят из трех основных элементов: пресинаптической мембраны, постсинаптической мембраны и синаптической щели (рис. 1).
Рис. 1. Строение синапса: 1 — микротрубочки; 2 — митохондрии; 3 — синаптические пузырьки с медиатором; 4 — пресинаптическая мембрана; 5 — постсинаптическая мембрана; 6 — рецепторы; 7 -синаптическая щель
Некоторые элементы синапсов могут иметь и другие названия. Например, синаптическая бляшка — это синапс между нейронами, концевая пластинка — постсинаптическая мембрана нервно-мышечного синапса, моторная бляшка — пресинаптическое окончание аксона на мышечном волокне.
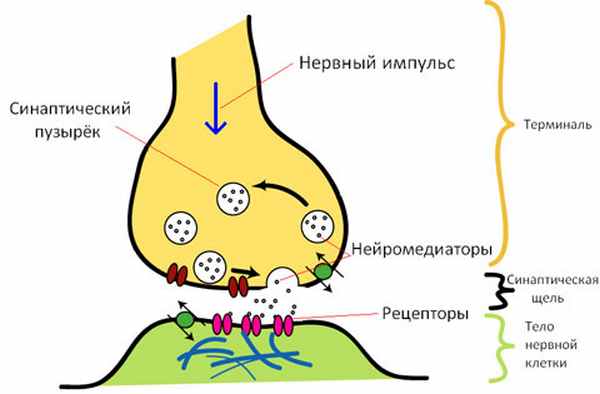
Пресинаптическая мембрана покрывает расширенное нервное окончание, которое представляет собой нейросекреторный аппарат. В пресинаптической части находятся пузырьки и митохондрии, обеспечивающие синтез медиатора. Медиаторы депонируются в гранулах (пузырьках).
Постсинаптическая мембрана — утолщенная часть мембраны клетки, с которой контактирует пресинаптическая мембрана. Она имеет ионные каналы и способна к генерации потенциала действия. Кроме того, на ней расположены специальные белковые структуры — рецепторы, воспринимающие действие медиаторов.
Синаптическая щель представляет собой пространство между пресинаптической и постсинаптической мембранами, заполненное жидкостью, близкой по составу к плазме крови.
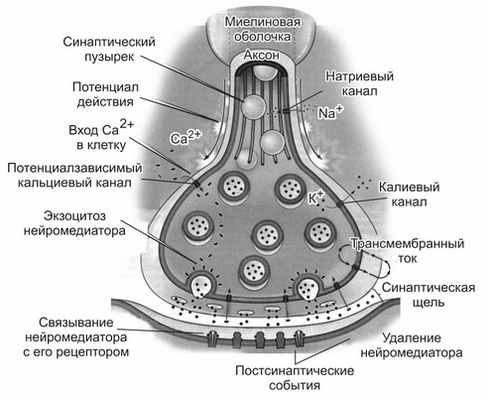
Рис. Строение синапса и процессы, осуществляемые в ходе синаптической передачи сигнала
Виды синапсов
Синапсы классифицируются по местоположению, характеру действия, способу передачи сигнала.
По месту положения выделяют нервно-мышечные синапсы, нервно-железистые и нейро-нейрональные; последние, в свою очередь, делятся на аксо-аксональные, аксо-дендритические, аксо-соматические, дендро-соматические, дендро-дендротические.
По характеру действия на воспринимающую структуру синапсы могут быть возбуждающими и тормозящими.
По способу передачи сигнала синапсы делятся на электрические, химические, смешанные.
Таблица 1. Классификация и виды синапсов

Классификация синапсов и механизм передачи возбуждения
Синапсы классифицируют следующим образом:
- по местоположению — периферические и центральные;
- по характеру их действия — возбуждающие и тормозящие;
- по способу передачи сигналов — химические, электрические, смешанные;
- по медиатору, с помощью которого осуществляется передача, — холинергические, адренергические, серотонинергические и т.д.
В химических синапсах возбуждение передается с помощью медиаторов (посредников).
Медиаторы — молекулы химических веществ, которые обеспечивают передачу возбуждения в синапсах. Другими словами химические вещества, участвующие в передаче возбуждения или торможения от одной возбудимой клетки к другой.
Свойства медиаторов
- Синтезируются в нейроне
- Накапливаются в окончании клетки
- Выделяются при появлении иона Са2+ в пресинаптическом окончании
- Оказывают специфическое действие на постсинаптическую мембрану
По химическому строению медиаторы можно подразделить на амины (норадреналин, дофамин, серотонин), аминокислоты (глицин, гамма-аминомасляная кислота) и полипептиды (эндорфины, энкефалины). Ацетилхолин известен в основном как возбуждающий медиатор и содержится в различных отделах ЦНС. Медиатор находится в пузырьках пресинаптического утолщения (синаптической бляшки). Медиатор синтезируется в клетках нейрона и может ресинтезироваться из метаболитов его расщепления в синаптической щели.
При возбуждении терминалей аксона происходит деполяризация мембраны синаптической бляшки, вызывающая поступление ионов кальция из внеклеточной среды внутрь нервного окончания через кальциевые каналы. Ионы кальция стимулируют перемещение синаптических пузырьков к пресинаптической мембране, их слияние с ней и последующий выход медиатора в синаптическую щель. После проникновения в щель медиатор диффундирует к постсинаптической мембране, содержащей на своей поверхности рецепторы. Взаимодействие медиатора с рецепторами вызывает открытие натриевых каналов, что способствует деполяризации постсинаптической мембраны и возникновению возбуждающего постсинаптического потенциала. В нервно-мышечном синапсе этот потенциал называется потенциалом концевой пластинки. Между деполяризованной постсинаптической мембраной и соседними с ней поляризованными участками этой же мембраны возникают местные токи, которые деполяризуют мембрану до критического уровня с последующей генерацией потенциала действия. Потенциал действия распространяется по всем мембранам, например, мышечного волокна и вызывает его сокращение.
Выделившийся в синаптическую щель медиатор связывается с рецепторами постсинаптической мембраны и подвергается расщеплению соответствующим ферментом. Так, холинэстераза разрушает медиатор ацетилхолин. После этого некоторое количество продуктов расщепления медиатора поступает в синаптическую бляшку, где из них снова ресинтезируется ацетилхолин.
В организме имеются не только возбуждающие, но и тормозные синапсы. По механизму передачи возбуждения они сходны с синапсами возбуждающего действия. В тормозных синапсах медиатор (например, гамма-аминомасляная кислота) связывается с рецепторами постсинаптической мембраны и способствует открытию в ней каналов для ионов хлора. При этом активизируется проникновение этих ионов внутрь клетки и развивается гиперполяризация постсинаптической мембраны, обусловливающая возникновение тормозного постсинаптического потенциала.
В настоящее время выяснено, что один медиатор может связываться с несколькими различными рецепторами и индуцировать различные реакции.
Химические синапсы
Физиологические свойства химических синапсов
Синапсы с химической передачей возбуждения обладают определенными свойствами:
- возбуждение проводится в одном направлении, так как медиатор выделяется только из синаптической бляшки и взаимодействует с рецепторами на постсинаптической мембраны;
- распространение возбуждения через синапсы происходит медленнее, чем по нервному волокну (синаптическая задержка);
- передача возбуждения осуществляется с помощью специфических медиаторов;
- в синапсах изменяется ритм возбуждения;
- синапсы способны утомляться;
- синапсы обладают высокой чувствительностью к различным химическим веществам и гипоксии.
Одностороннее проведение сигнала. Сигнал передается только от пресинаптической мембраны к постсинаптической. Это вытекает из особенностей строения и свойств синаптических структур.
Замедленная передача сигнала. Обусловлена синаптической задержкой в передаче сигнала с одной клетки на другую. Задержка вызывается временными затратами на процессы выброса медиатора, его диффузии к постсинаптической мембране, связывания с рецепторами постсинаптической мембраны, деполяризации и преобразования постсинаптического потенциала в ПД (потенциал действия). Длительность синаптической задержки колеблется от 0,5 до 2 мс.
Способность к суммации эффекта от приходящих к синапсу сигналов. Такая суммация проявляется, если последующий сигнал приходит к синапсу через короткое время (1- 10 мс) после предыдущего. В таких случаях амплитуда ВПСП возрастает и на постсинаптическом нейроне может генерироваться большая частота ПД.
Трансформация ритма возбуждении. Частота нервных импульсов, приходящих к пресинаптической мембране, обычно не соответствует частоте ПД, генерируемых постсинаптическим нейроном. Исключение составляют синапсы, передающие возбуждение с нервного волокна на скелетную мышцу.
Низкая лабильность и высокая утомляемость синапсов. Синапсы могут проводить 50-100 нервных импульсов в секунду. Это в 5-10 раз меньше, чем максимальная частота ПД, которую могут воспроизводить нервные волокна при их электростимуляции. Если нервные волокна считаются практически неутомляемыми, то в синапсах утомление развивается весьма быстро. Это происходит из-за истощения запасов медиатора, энергетических ресурсов, развития стойкой деполяризации постсинаптической мембраны и т.д.
Высокая чувствительность синапсов к действию биологически активных веществ, лекарственных препаратов и ядов. Например, яд стрихнин блокирует функцию тормозных синапсов ЦНС, связываясь с рецепторами, чувствительными к медиатору глицину. Столбнячный токсин блокирует тормозные синапсы, нарушая выделение медиатора из пресинаптической терминали. В обоих случаях развиваются опасные для жизни организма явления. Примеры действия биологически активных веществ и ядов на передачу сигналов в нервно-мышечных синапсах рассмотрены выше.
Свойства облегчения и депрессии синоптической передачи. Облегчение синаптической передачи имеет место, когда нервные импульсы поступают к синапсу через короткое время (10-50 мс) друг за другом, т.е. достаточно часто. При этом в течение некоторого промежутка времени каждый последующий ПД, приходящий к пресинаптической мембране, вызывает увеличение содержания медиатора в синаптической щели, возрастание амплитуды ВПСП и увеличение эффективности синаптической передачи.
Одним из механизмов облегчения является накопление ионов Са 2 в пресинаптической терминали. Для удаления кальциевым насосом порции кальция, вошедшей в синаптическую терминаль при поступлении ПД, необходимо несколько десятков миллисекунд. Если в это время приходит новый потенциал действия, то новая порция кальция входит в терминаль и ее эффект на высвобождение нейромедиатора складывается с остаточным количеством кальция, которое кальциевый насос не успел удалить из нейроплазмы терминали.
Имеются и другие механизмы развития облегчения. Этот феномен в классических руководствах по физиологии называют также посттетанической потенциацией. Облегчение синаптической передачи имеет значение в функционировании механизмов памяти, для образования условных рефлексов и обучения. Облегчение передачи сигналов лежит в основе развития пластичности синапсов и улучшения их функций при частой активации.
Депрессия (угнетение) передачи сигналов в синапсах развивается при поступлении очень частых (для нервно-мышечного синапса более 100 Гц) нервных импульсов к пресинаптической мембране. В механизмах развития явления депрессии имеют значение истощение запасов медиатора в пресинаптической терминали, снижение чувствительности рецепторов постсинаптической мембраны к медиатору, развитие стойкой деполяризации постсинаптической мембраны, затрудняющих генерацию ПД на мембране постсинаптической клетки.
Электрические синапсы
Кроме синапсов с химической передачей возбуждения в организме есть синапсы с электрической передачей. Эти синапсы имеют очень узкую синаптическую щель и пониженное электрическое сопротивление между двумя мембранами. Благодаря наличию поперечных каналов между мембранами и низкому сопротивлению, электрический импульс легко проходит через мембраны. Электрические синапсы обычно характерны для однотипных клеток.
В результате воздействия раздражителя пресинаптический потенциал действия раздражает постсинаптическую мембрану, где возникает распространяющийся потенциал действия.
Электрические синапсы характеризуются большей скоростью проведения возбуждения по сравнению с химическими синапсами и низкой чувствительностью к воздействию химических веществ.
Электрические синапсы бывают с одно- и двусторонней передачей возбуждения.
В организме встречаются и электрические тормозные синапсы. Тормозное влияние развивается за счет действия тока, который вызывает гиперполяризацию постсинаптической мембраны.
В смешанных синапсах может происходить передача возбуждения с помощью как электрических импульсов, так и медиаторов.
Читайте также: