Механизм дивергенции и конвергенции внутриклеточных сигналов
Добавил пользователь Валентин П. Обновлено: 10.01.2026
Мозговая деятельность образована множеством различных процессов в отдельных нейронах, между нейронами, сенсорными системами, другими нервными образованиями и центрами, расположенными на разных «этажах» центральной нервной системы, в высших отделах мозга и коре больших полушарий. Ее функционирование целостно, системно и подчинено обеспечению равновесного состояния внутренней среды организма и его взаимоотношений с внешней средой. Регулирующую роль в мозговой деятельности выполняют особые механизмы — разные исследователи называют их еще закономерностями, принципами, просто процессами.
Механизмы дивергенции 1 и конвергенции [1] [2] обеспечивают растекание и схождение (концентрацию) физиологических процессов в нервной системе. Первый осуществляет диффузное, т.е. ненаправленное, растекание нервного сигнала по разным отделам мозга, центрам, полям, направлениям, что можно сравнить с волнами, разбегающимися по воде от брошенного в него камня, но в нервной системе это не так округло и ритмично. Второй механизм вызывает уменьшение возбужденного мозгового пространства, свертывание возбуждения в обратном дивергенции направлению — к определенному участку (часто к тому, где оно возникло) или взаимосвязанной группе их. Импульс возбуждения, пришедший в головной мозг и его кору по восходящим нервным путям, вызывает возбуждение определенной группы нервных клеток, области коры, и в них возникает так называемый очаг возбуждения — точка, а чаще область повышенного возбуждения (рис. 4.1 А). Благодаря богатым связям нервных клеток, особенно в подкорке и коре мозга, импульс вступает во взаимодействие с соседними и распространяется от места возникновения в разные стороны (рис. 4.1 Б). Этот вид процесса дивергенции называется иррадиацией (от лат. irradio — освещаю лучами, сияю).
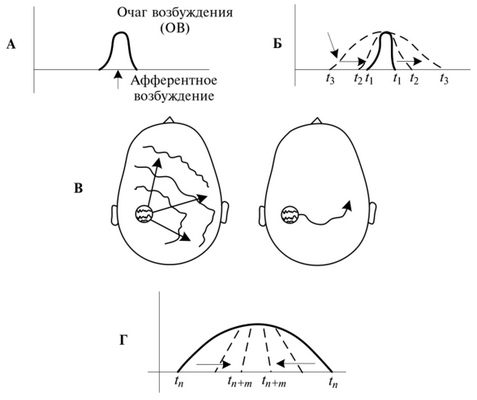
Рис. 4.1. Схема иррадиации и концентрации возбуждения в коре больших полушарий:
А — возникновение очага возбуждения; Б — распространение (иррадиация) возбуждения по коре; В — генерализованный и избирательный виды иррадиации; Г — концентрация возбуждения
Иррадиация возбуждения может быть генерализованной и избирательной (рис. 4.1 В). Область больших полушарий, охватываемая растекающимся из очага возбуждением, зависит от силы раздражения, функционального состояния мозга и сформированных ранее нервных связей. Чем сильнее раздражение и чем выше возбуждение, тем более генерализованный характер носит его иррадиация. Иррадиация способствует мобилизации возможностей мозга и обеспечивает подключение его отделов к выработке ответа на важные (сильные) раздражители. Но в генерализванном виде она может иметь и отрицательное значение, особенно когда приобретает застойный характер. Так, новая обстановка, опасность, первая попытка выполнения ответственного действия вызывают у человека усиленную и продолжающуюся иррадиацию возбуждения, которая может породить известные педагогам и психологам факты чрезмерного волнения, дрожания, растерянности человека и ухудшать его действия.
Избирательная иррадиация выражается в локальном распространении возбуждения (рис. 4.1 В, справа), что сопровождается образованием или использованием сложившихся ранее нервных связей. Она характерна для действий человека в привычных условиях, при выполнении хорошо освоенных им действий, проявлении самообладания (сдерживание растекания возбудительного потока в определенном русле или участке с помощью произвольного сдерживания — торможения). Это экономный вид иррадиации, сохраняющий человеку силы и открывающий возможность для работы мозга по выполнению параллельных действий (например, сохранение спортсменом концентрации внимания, манипулирование водителя органами управления автомашиной с одновременным бдительным наблюдением за дорожной обстановкой, ведение обучающимся конспекта с одновременным внимательным прослушиванием речи преподавателя).
Ослабление или прекращение инициации из очага возбуждения приводит к тому, что иррадиированное возбуждение постепенно возвращается (сходится, конвергирует) к его центру, где сила возбуждения была наивысшей, и поэтому оно наиболее устойчиво (рис. 4.1 Г). Этот процесс называется концентрацией. Иррадиация совершается за несколько секунд (до минуты), а концентрация идет медленнее в 4—5 раз.
В процессах высшей нервной деятельности в больших полушариях и коре обнаруживается взаимовлияние процессов, называемое индукцией. Это влияние сопровождается реципрокностью (противоположностью, противодействием): возбуждение одного из мозговых центров, обслуживающих определенную активность, сопряжено с угнетением более слабого очага иной активности и даже подавлением его. Если это влияние одновременно, то имеет место одновременная (или пространственная) индукция. Но бывает, что после исчезновения очага возбуждения в какой-то части головного мозга развивается торможение или, наоборот, после торможения возникает возбуждение, т.е. происходит последовательная индукция. Эти явления обнаруживаются в общеизвестных фактах: после сна человек испытывает прилив сил; после длительной и трудной умственной работы или сильного возбуждения определенное время существует чувство усталости или может тянуть ко сну; если после длительной ходьбы присесть на 30—40 мин, то возбуждение может успеть смениться торможением и продолжать ходьбу будет трудно.
Дивергенция возбуждения
Лат. diverqere - направляется в разные стороны - способность одиночного нейрона устанавливать многочисленные синаптические связи с различными нервными клетками. Благодаря процессу дивергенции одна и та же клетка может участвовать в организации различных реакций и контролировать большее число нейронов. В то же время каждый нейрон может обеспечивать широкое перераспределение импульсов, что приводит к иррадиации возбуждения.
Иррадиация (от лат. irradio — сияю, испускаю лучи) в физиологии, распространение процесса возбуждения или торможения в центральной нервной системе. Важную роль И. играет в деятельности коры больших полушарий головного мозга. И. возбуждения особенно отчётливо проявляется при сильном раздражении, когда в рефлекторный ответ вовлекаются нервные центры, обычно в нём не участвующие. Так, на умеренное болевое раздражение кожи стопы животное отвечает сгибанием лапы в голеностопном суставе; увеличение силы раздражения приводит к сгибанию ноги в коленном и тазобедренном суставах. При изучении действия тормозного условного раздражителя И. П. Павловым было показано, что торможение также может распространяться (иррадиировать) в клетках коры больших полушарий
Принцип иррадиации, или дивергенции, возбуждения ЦНС. Иррадиация возбуждения (от лат. irradio, озарять, освещать) - это распространение процесса возбуждения из одного участка ЦНС в другой. Каждый нейрон за счет многочисленных ответвлений (дивергенции), заканчивающихся синапсами, и большого числа вставочных нейронов связан со многими другими нейронами. Поэтому нервные импульсы от одного нейрона могут быть направлены к тысяче других нейронов. Процесс иррадиации возбуждения регулируется различными механизмами. Он может быть усилен, например, за счет активации ретикулярной формации ствола мозга. С другой стороны, процесс иррадиации ограничивается, и это важное действие осуществляется с участием многочисленных тормозных нейронов. Например, в спинном мозге ограничение иррадиации осуществляется благодаря механизму возвратного торможения с помощью специальных тормозных интернейронов - клеток Реншоу. Иррадиация возбуждения играет исключительно важную роль, так как позволяет обмениваться многочисленными потоками информации различным структурам мозга. Именно за счет иррадиации происходит обмен информацией между первой и второй сигнальными системами (элективная иррадиация), что существенно увеличивает возможности высшей нервной деятельности человека. Благодаря иррадиации возбуждения осуществляется интеграция деятельности правого и левого полушарий.
Принцип концентрации возбуждения
Возбуждение и торможение могут либо иррадиировать, либо концентрироваться. Концентрация - это явление, противоположное иррадиации. Оно возникает в тот момент, когда иррадиация достигает определенной границы, после чего распространение возбуждения или торможения идет в обратном направлении, т. е. в исходный пункт. Если нейроны находятся в заторможенном состоянии, это препятствует распространению на них процесса возбуждения, и наоборот, если они находятся в состоянии возбуждения, торможением они охватываются труднее. На скорость иррадиации и концентрации нервных процессов оказывает влияние и тип нервной системы, ее индивидуальные особенности.
Принцип индукции нервных процессов. Индукция отражает проявление процессов иррадиации возбуждения и торможения в коре больших полушарий. Принято считать, что индукция - (от лат. Inductio - введение, наведение) обозначает возникновение нервного процесса, противоположного по знаку процессу, вызванному условным раздражителем (положительным или отрицательным, т. е. тормозным). Одновременная индукция заключается в том, что формирование в каком-либо центре коры больших полушарий концентрированного возбуждения вызывает в прилежащих к этому центру зонах торможение (отрицательная одновременная индукция), а такое же концентрированное торможение вызывает в этих зонах возбуждение (положительная одновременная индукция). Таким образом, при одновременной индукции нервный процесс вызывает в другом участке коры процесс, противоположный по знаку (процесс возбуждения вызывает процесс торможения и наоборот), а при последовательной индукции происходит смена противоположных нервных процессов в одном и том же участке.
Принцип конвергенции возбуждения (или принцип общего конечного пути, воронка Шеррингтона). Конвергенция нервных импульсов (от лат. convergo, convergere - сближать, сходиться) означает схождение к одному нейрону двух или нескольких различных возбуждений одновременно. Это явление было открыто Ч. Шеррингтоном. Он показал, что одно и то же движение, например рефлекторное сгибание конечности в коленном суставе, можно вызвать путем раздражения различных рефлексогенных зон. В связи с этим им было введено понятие "общего конечного пути", или "принципа воронки", согласно которому потоки импульсов от различных нейронов могут сходиться на одном и том же нейроне (в данном случае - на альфа-мотонейронах спинного мозга). В частности, Ч. Шеррингтон обнаружил схождение к одним и тем же промежуточным или эфферентным нейронам различных афферентов от разных участков общего рецептивного поля (в спинном и продолговатом мозге) или даже от разных рецептивных полей (в высших отделах головного мозга). В настоящее время показано, что конвергенция возбуждения, так же как и дивергенция возбуждения, - очень распространенное явление в ЦНС. Основой для конвергенции (как и для иррадиации) является определенная морфологическая и функциональная структура различных отделов мозга. Очевидно, что часть конвергентных путей является врожденными, а другая часть (главным образом в коре большого мозга) - приобретенная в результате обучения в процессе онтогенеза. Формирование новых конвергентных отношений для нейронов коры большого мозга в процессе онтогенеза во многом связаны с формированием в коре доминантных очагов возбуждения, которые способны "притягивать" к себе возбуждение от других нейронов.
Принцип доминанты, или господствующего очага возбуждения. Доминанта (от лат. dominans, dominantis - господствующий) - это временно господствующая рефлекторная система, обусловливающая интегральный характер функционирования нервных центров в какой-либо период времени и определяющая целесообразное поведение животного и человека. В целом принцип доминанты означает, что текущая деятельность мозга определяется наличием господствующего (доминантного) очага возбуждения, или господствующего нейронного объединения, которое в данный момент времени подавляет и подчиняет себе деятельность остальных нейронных образований. Таким образом, благодаря формированию доминантного очага (доминантного нейронного объединения) деятельность мозга организуется таким образом, чтобы удовлетворить потребность организма, наличие которой и сформировало доминантный очаг возбуждения. Детальное изучение свойств доминантного очага показало, что для совокупности нейронов, входящих в его состав, характерны повышенная возбудимость, повышенная стойкость возбуждения, или инертность (заключающаяся в том, что у таких нейронов сложно вызвать торможение), повышенная способность к суммированию возбуждения, высокая способность "притягивать" возбуждение от других нервных центров (и тем самым повышать свою активность). Доминанта, как один из основных принципов координационной деятельности ЦНС, имеет важное значение в жизни человека. Например, именно благодаря доминанте возможно сосредоточение психической деятельности (внимание) и выполнение умственной или физической трудовой деятельности (в данном случае - это трудовая доминанта). В период поиска пищи и ее поедания реализуется пищевая доминанта. В настоящее время в отношении человека выделяют различные виды доминант (пищевую, оборонительную, половую, игровую, трудовую и др.).
Принцип субординации, или соподчинения, также относится к категории важнейших принципов организации работы мозга. Согласно этому принципу, деятельность нижележащих отделов мозга контролируется и управляется вышележащими отделами ЦНС. Например, в двигательных системах мозга и в вегетативной нервной системе имеются нейронные объединения (нервные центры), расположенные в спинном мозге или в стволе мозга, которые подчиняются деятельности нейронных объединений (нервных центров), находящихся в гипоталамусе, таламусе, мозжечке, базальных ядрах и коре больших полушарий.
Принцип обратной связи (обратной афферентации) и копий эфферентаций. Согласно этому принципу, для точной координации деятельности различных нейронных объединений (нервных центров, рефлекторных дуг) необходима оптимальная по объему информация о результатах действия. Она поступает в мозг по сенсорным каналам. Отсутствие такой информации приводит к дезинтеграции деятельности мозга. Особенно наглядна роль обратной афферентации при реализации двигательной активности - нарушение проприоцептивной чувствительности, как правило, препятствует выполнению точных движений, а также нарушает возможность формирования и сохранения адекватной для данного движения позы.
Принцип реципрокности (сопряжения) возбуждения и торможения на уровне спинного мозга реализуется с участием реципрокного торможения, благодаря которому возникают безусловные двигательные реципрокные рефлексы. Реципрокное торможение осуществляется по механизму постсинаптического торможения, которое возникает с участием специальных вставочных тормозных нейронов.
Принципы кодирования информации в нервной системе. В целом вся информация или значительная ее часть, передаваемая в ЦНС от одного отдела к другому, заключена в пространственном и временном распределении импульсных потоков, при этом используются различные нейронные коды. Выделяют три основные группы кодов. Неимпульсные сигналы, для которых характерны внутри- и внеклеточные факторы. К внутриклеточным факторам относятся амплитудные характеристики рецепторных и синаптических потенциалов, амплитудные и пространственные характеристики изменений синаптической проводимости, пространственное и временное распределение характеристик мембранного потенциала и градуальные потенциалы в аксонных терминалях. Внеклеточные факторы - это высвобождение медиаторов и ионов калия, нейросекреция, электротонические взаимодействия. Импульсные сигналы в одиночных нейронах. Для импульсных кодов главными кандидатами являются коды пространственные ("меченые линии", т. е. представление информации номером канала) и временные - различные виды частотных или интервальных кодов (взвешенное среднее значение частоты, мгновенное значение частоты, частота разряда, форма интервальных гистограмм и т. д.). Выделяют также микроструктурное кодирование (временный узор импульсов), латентный код (момент появления или фазовые изменения разряда), числовой код (количество импульсов в пачке), код длинной пачки (длительность импульсации), наличие отдельного импульса или его отсутствие) изменение скорости распространения возбуждения в аксоне и пространственную последовательность явлений в аксоне. Ансамблевая активность (кодирование по ансамблю). В большинстве случаев в ЦНС используется пространственно-временное кодирование, когда информация о признаках сигнала передается канально и уточняется различными модификациями временных кодов.
2)Надпочечники (glandula suprarenalis, ед. ч.)-парные железы внутренней секреции, расположенные над верхними полюсами почек. У человека они находятся на уровне XI грудного - I поясничного позвонков , забрюшинно. Правый надпочечник имеет треугольную форму , левый - полулунную; вогнутые основания надпочечников примыкают к выпуклым полюсам почек. Вместе с почками надпочечники заключены в жировую капсулу ( сapsula adiposa ) и покрыты почечной фасцией ( fascia renalis ). Длина надпочечника взрослого человека варьирует от 30 до 70 мм, ширина - от 20 до 35 мм , толщина - от 3 до 10 мм, масса обоих надпочечников составляет 10 -14 г. Снаружи надпочечник покрыт соединительнотканной капсулой, от которой в паренхиму отходят перегородки, заключающие в себе сосуды и нервы и делящие паренхиму надпочечников на группы клеток и клеточные тяжи. В надпочечниках различают наружное корковое вещество, составляющее примерно 2/3 всей массы надпочечника , и внутреннее мозговое вещество. У новорожденных масса надпочечника в среднем 3,5 г. С возрастом она увеличивается, постепенно разграничиваются корковое и мозговое вещество, отдельные зоны коркового вещества.
Кровоснабжение надпочечников осуществляют три группы надпочечниковых артерий: верхняя, средняя, нижняя, проникающие в паренхиму в виде многочисленных капилляров, которые широко анастамозируют между собой и образуют в мозговом веществе синусоиды - расширения. Отток крови от надпочечников происходит через центральную и многочисленные поверхностные вены, впадающие в венозную сеть окружающих органов и тканей. Параллельно кровеносным расположены лимфатические капилляры, отводящие лимфу. Иннервируются надпочечники симпатическими (преимущественно) и парасимпатическими волокнами чревного, блуждающего и диафрагмального нервов . Корковое вещество надпочечника делится на клубочковую, пучковую и сетчатую зоны. Клубочковая зона, прилегающая тонким слоем к соединительнотканной капсуле, состоит из клеток неправильной формы. В пучковой, средней, наиболее широкой зоне группируются железистые клетки, расположенные радиальными колонками, а в сетчатой, внутренней, зоне группы клеток имеют вид неправильной сетки. Корковое вещество надпочечника богато липидами, окрашивающими его в желтый цвет. Гормоны коркового вещества кортикостероиды (глюко- и минералокортикоиды) синтезируются в митохондриях секреторных клеток из холестерина. Многообразное влияние кортикостероидов на все виды обмена веществ, сосудистый тонус, иммунитет и др. делает корковое вещество надпочечников важнейшим участком жизнеобеспечения человека в обычных условиях и в условиях адаптации к различным стрессам. В клубочковой зоне коркового вещества синтезируется альдостерон - основной минералокортикоид, участвующий в регуляции водно-солевого обмена. В пучковой зоне синтезируется преимущественно кортизол - глюкокортикоид, влияющий на белковый, жировой и углеводный обмен и на обмен нуклеиновых кислот, и кортикостерон, обладающий свойствами глюко- и минералокортикоида. В сетчатой зоне образуются половые гормоны, главным образом андрогены. Синтез кортикостероидов, прежде всего глюкокортикостероидов, регулируются адренокортикотропным гормоном. В глубине надпочечника находится мозговое вещество. Железистые клетки мозгового вещества получили название хромаффинных, или феохромных, т.к. избирательно окрашиваются солями хрома в желто-бурый цвет. Помимо железистых клеток в мозговом веществе надпочечника много нервных волокон и нервных клеток. Скопления хромаффинных нервных клеток, так называемых параганглиев, обнаруживают также по ходу легочного ствола и восходящей части аорты, в средостении находится поясничный аортальный параганглий и другие. Хромаффинные клетки секретируют три гормона (адреналин, норадреналин и дофамин), объединенных под общим названием катехоламины. Биосинтетическими предшественниками этих гормонов является аминокислота тирозин. Адреналин синтезируется только в надпочечниках; норадреналин и дофамин образуются также в параганглиях и многочисленных нейронах симпатической нервной системы. Все ткани, продуцирующие катехоламины, составляют адреналиновую систему. Активность хромаффинных клеток стимулируют различные воздействия окружающей и внутренней среды: эмоции, артериальная гипотензия, гипогликемия, физическая нагрузка, охлаждение и др.
Механизм дивергенции и конвергенции внутриклеточных сигналов
• Сигнальные сети состоят из групп биохимических реакций, которые напоминают функции математической логики, интегрирующие информацию
• Для обработки информации на более высоком уровне комбинации таких логических функций объединяются в сигнальные сети
Как отмечалось в статьях на сайте, функции передачи сигналов, используемые для интеграции информации и ее направления к клеточным мишеням, удивительно напоминают функции математической логики, которые используются для проектирования индивидуальных цепей электронных компьютеров.
Действительно, существуют биологические эквиваленты практически всех функциональных компонентов, которые хорошо известны компьютерщикам и инженерам, занимающимся конструированием компьютеров и электронных контрольных приборов. Поэтому для того, чтобы понять, как функционируют пути передачи сигнала, полезно рассмотреть отдельные группы реакций в составе процесса как составляющие логической цепи, похожей на компьютерную.
Простейшим примером служит конвергенция двух процессов, ведущих к стимуляции. Если каждый из них генерирует сигнал, достаточный для того, чтобы вызвать ответ, то конвергенция представит функцию «ИЛИ». Если ни на одном из входов сигнал недостаточен, но комбинация обоих вызывает ответ, то конвергирующий путь создаст функцию «И». Цепи типа И также рассматриваются как детекторы совпадений, т. е. ответ генерируется только в том случае, когда одновременно активируются два пути стимуляции.
Функция И возникает в результате комбинации двух близких, но в количественном отношении неадекватных входных сигналов. Наоборот, для генерации ответа могут потребоваться оба механистически разных входных сигнала. Примером служит белок-мишень, который аллостерически активируется только при фосфорилировании, или же он активируется при фосфорилировании, но начинает функционировать только после своей транслокации в специфические внутриклеточные сайты.
Противоположность функции «И» представляет собой функция «НЕ». Она характеризуется тем, что один путь блокирует стимулирующий эффект другого. Во многих точках клеточных сигнальных путей существуют простые логические шлюзы.
Можно также предполагать существование конвергентного пути передачи сигнала в количественном, а не в булевом смысле, рассматривая аддитивность входных сигналов как отдельный процесс. Функция «ИЛИ», относящаяся к верхнему рисунку, может рассматриваться как аддитивный положительный входной сигнал от двух путей. Такая аддитивность может отражать способность двух рецепторов стимулировать пул особого G-белка, или способность двух протеинкиназ фосфорилировать один субстрат.
Аддитивность может приводить к положительному сигналу, как в примере выше, или к отрицательному, когда объединяются два ингибиторных входных сигнала. Ингибирование и стимуляция могут также объединяться аддитивно, давая на выходе алгебраически сбалансированный сигнал. Наоборот, множественные входные сигналы способны объединяться, давая суммарный сигнал больше или меньше аддитивного. Обсуждавшаяся выше Функция НЕТ аналогична описывающей блокаду стимуляции.Функция И описывает синергизм, при котором один входной сигнал потенцирует другой, но сам по себе дает небольшой эффект.
Даже для простой сигнальной сети характерна сложная схема обработки информации. Хорошим примером служит создание «памяти»: преобразование эффекта временного сигнала в более или менее постоянный. Системы передачи располагают множеством механизмов формирования памяти и забывания сигнала. Один из механизмов, общий для протеинкиназного пути, представляет собой цикл положительной обратной связи, и представлен в верхней части рисунка ниже. В этом цикле сигнал на входе стимулирует передатчик (Т), который, в свою очередь, активирует эффекторный белок (Е), формирующий выходной сигнал.
Если эффектор также может стимулировать передатчик, то часть начального сигнала может передаваться на передатчик. При этом передатчик может обеспечивать появление на выходе полного сигнала эффектора, даже при отсутствии входного сигнала. Как показано на рисунке ниже справа, для такой системы обычно характерно существование порогового эффекта.
Положительная опережающая связь может генерировать память другого типа, которая отмечает продолжительность входного сигнала. В таких цепях эффектору необходимы одновременные входные сигналы от рецептора и от промежуточного передатчика. Если путь передачи сигнала от рецептора через передатчик относительно медленный или же необходимо накопление значительных количеств передатчика, ответ включится только при продолжительном сигнале. Это показано на временной диграмме выходного сигнала справа.
Третий путь запоминания представляет собой разрешение одному входному сигналу контролировать обратимость второго регуляторного события. Белок WASP, который инициирует полимеризацию актина, обеспечивающую подвижность клетки и изменение ее формы, активируется при фосфорилировании и при связывании с Cdc42, который представляет собой небольшой ГТФ-связывающий белок (G). Однако фосфорилированный сайт на WASP открывается только после его связывания с Cdc42. Таким образом, для фосфорилирования требуется активированный Cdc42 и активированная протеинкиназа.
Если Cdc42 диссоциирует, фосфорилированное состояние WASP поддерживается до тех пор, пока с ним опять не свяжется другая сигнальная молекула, природа которой неизвестна. При этом снова откроется сайт для протеинфосфатазы. Как представлено на графике временной заисимости, связывание с Cdc42 приводит к активирующему эффекту, а связывание только с киназой не оказывает такого действия. Если присутствует Cdc42, то киназа может активировать WASP. Фосфо-WASP относительно устойчив к одной протеинфосфатазе (Р), однако он дефосфорилируется, если Cdc42 или другой G-белок связывается с ним и открывает сайт для фосфатазы.
Для обработки поступающей информации сигнальные сети используют простые логические функции.
Булевы функции «ИЛИ», «И» и «НЕ» (слева) соответствуют количественным взаимоотношениям между конвергирующими сигналами, представленными справа. Относительно сложный процесс обработки сигнала осуществляется с помощью простых мультибелковых модулей.
На рисунке представлены три типа сигнальных модулей (слева) и их реакция на присутствие агонистов (справа), (сверху)
В модуле положительной обратной связи белок-передатчик (Т) стимулирует эффектор (Е), который продуцирует сигнал на выходе.
Однако эффектор также стимулирует активность передатчика. В результате образуется выключатель типа «все или ничего».
При этом входной сигнал, величина которого не превышает определенного порогового значения, оказывается малоэффективным.
Однако эффективность сигнала резко возрастает, когда обратной связи с эффектором достаточно для поддержания активности передатчика, даже в отсутствие генерации постоянного входного сигнала от рецептора. (в центре)
В положительном опережающем модуле эффектор принимает сигналы от передатчика и от элементов, расположенных перед ним.
Когда стимуляция носит кратковременный характер (короткая горизонтальная полоса под графиком справа), накопления существенных количеств активного передатчика не происходит, и сигнал на выходе достигает минимальной величины.
При более продолжительной стимуляции (длинная горизонтальная полоса) на выходе получается более сильный сигнал, (внизу).
В некоторых модулях переключения двойного контроля, связывание одного регулятора (G) активирует эффектор и демаскирует другой регуляторный сайт, показанный на рис. как сайт Ser субстрата (-ОН) протеинкиназы.
Эффектор может фосфорилироваться или дефосфорилироваться только в том случае, когда он связан с G-белком.
Поэтому, как показано на рисунке справа, добавление одного G будет оказывать активирующий эффект, однако активации одной киназы (К) наблюдаться не будет.
Если киназа активна, пока она связана с белком G, то фосфорилированная форма устойчива к действию фосфатазы, если только G не связался повторно и не произошло демаскирования фосфосеринового остатка (показан на графике справа буквой Р).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Дивергенция и конвергенция сигналов.
Часто стимулы, поступающие к нервному волокну по одному, возбуждают гораздо большее число нервных волокон. Этот феномен называют дивергенцией. Благодаря дивергенции одна и та же нервная клетка может принимать участие в организации различных реакций и контролировать большое количество нейронов. Одновременно каждый нейрон может обеспечивать широкое перераспределение импульсов, что ведет к иррадиации возбуждения(активное распространение возбуждения в ЦНС, особенно при сильном и длительном раздражении). Конвергенция и дивергенция взаимно связаны.Существуют два основных типа дивергенции, функциональная роль которых различна.
Расходящийся тип дивергенции, при котором происходит распространение действия входящего сигнала на все большее количество нейронов по мере того, как сигнал проходит через последовательный ряд нейронов. Такой тип дивергенции характерен для кортикоспинального тракта, управляющего скелетными мышцами. При этом одна большая пирамидная клетка в двигательной области коры большого мозга при чрезвычайно облегченных условиях способна возбудить до 10000 мышечных волокон.Второй тип дивергенции, приводит к расхождению нервных путей с образованием многочисленных трактов. В данном случае сигнал передается в двух направлениях от пула(несколько мотонейронов могут иннервировать одну мышцу, в этом случае они образуют так называемый мотонейронный пул). Например, информация, передаваемая в восходящем направлении по задним столбам спинного мозга, в нижней части головного мозга направляется двумя разными путями: (1) в мозжечок; (2) через нижние области головного мозга к таламусу и коре большого мозга. Аналогично в таламусе почти вся сенсорная информация передается одновременно в еще более глубокие структуры самого таламуса и различные области коры большого мозга.
На каждом из нейронов ЦНС конвергирует (сходятся) различные афферентные волокна.Таких афферентных входов для большинства нейронов много десятков и даже тысяч. Так, на мотонейронах заканчиваются в среднем 6000 коллатералей аксонов, которые поступают от периферических рецепторов и различных структур мозга, образуя возбуждающие и тормозные синапсы. Это такое универсальное явление, можно говорить о принципе конвергенции в нейронах и их связях. Благодаря этому явлению в один и тот же нейрон одновременно поступают многочисленные и разнообразные потоки возбуждений, которые затем подлежат сложной обработке и перекодируются и формируются в единое возбуждение - аксону, что идет к следующему звену нервной сетки. Конвергенция возбуждения на нейроне является универсальным фактором его интегративной деятельности.
Конвергенция означает объединение сигналов множественных входов на одном нейроне. Это значит, что на одном нейроне заканчиваются многочисленныетерминали нервных волокон одиночного тракта. Этот тип конвергенции важен, поскольку нейроны почти никогда не возбуждаются потенциалом действия одной входящей терминали. Но потенциалы действия многих терминалей, конвергирующих на нейроне, обеспечивают достаточную пространственную суммацию, чтобы сдвинуть мембранный потенциал нейрона до порогового уровня,необходимого для его возбуждения.
Возможна также конвергенция сигналов (возбуждающих или тормозящих), исходящих из многих источников, как показано на рисунке. Например, на вставочных нейронах спинного мозга конвергируют сигналы от: (1) периферических нервных волокон, входящих в спинной мозг; (2) проприоспинальных волокон, идущих от одного сегмента спинного мозга к другому; (3) кортикоспинальных волокон из коры большого мозга; (4) нескольких других длинных нисходящих путей из головного в спинной мозг. Затем сигналы от вставочных нейронов сходятся на мотонейронах спинного мозга, непосредственно управляющих функцией скелетных мышц.
Относительная конвергенция - в спинном и стволовом мозге - конвергенция импульсов от различных рецепторных полей одного и того же рефлекса. Абсолютная конвергенция - в коре головного мозга имеются полимодальные (полисенсорные) нейроны, к ним сходятся импульсы от различных рецепторов. Принцип конвергенции обеспечивается за счет изменения частотных характеристик нервных импульсов, идущих от объединяющего нейрона.
Различают мультисенсорную, мультибиологическую и сенсорно-биологическую формы конвергенции. В первом случае на нейрон поступают сигналы различной сенсорной модальности (зрительные, слуховые, болевые и др.), во втором - потоки возбуждений различной биологической модальности (пищевые, половые и др.), в третьем - сигнализация (зрительная, пищевая) и другие.
Такая конвергенция позволяет осуществлять суммацию информации из различных источников, а ответная реакция нейрона является результатом интеграции всей этой информации. Конвергенция — один из важных способов, с помощью которых центральная нервная система коррелирует, интегрирует и сортирует различные типы информации.
• Чувствительность путей передачи сигналов регулируется таким образом, который позволяет изменять ответ клетки при изменении силы сигнала в широких пределах
• Во всех путях передачи сигналов эта функция регулируется с помощью механизмов обратной связи
• Большинство систем передачи сигналов содержат много адаптивных циклов обратной связи, которые управляют сигналами различной силы и продолжительности
Универсальным свойством систем внутриклеточной передачи сигналов является их адаптируемость к входному сигналу. Клетка постоянно регулирует свою чувствительность к сигналам с тем, чтобы поддерживать у себя способность улавливать изменения входного сигнала. Обычно, когда в клетку поступает новый сигнал, в ней начинается процесс десенсибилизации, который заглушает ее ответ до размеров нового плато, лежащего ниже пика начального ответа.
При удалении сигнала, состояние десенсибилизации сохраняется, и чувствительность клетки медленно возвращается к обычному уровню. Аналогичным образом, удаление тонического сигнала может привести к гиперчувствительности сигнальной системы.
Адаптируемость сигнальных систем представляет собой один из лучших примеров биологического гомеостаза. Эта адаптируемость оказывается впечатляющей. Обычно клетки способны регулировать свою чувствительность к физиологическим сигналам более чем в 100-кратном диапазоне, и зрительная реакция у млекопитающих может адаптироваться к изменению интенсивности источника света в 10 7 раз.
Такая невероятная способность позволяет фоторецепторной клетке реагировать только на один фотон и дает возможность читать как в сумерках, так и при ярком солнечном свете. Способность к адаптации характерна для бактерий, растений, грибов и животных. Многие процессы адаптации достаточно консервативны с точки зрения эволюции, хотя наиболее сложные адаптационные механизмы существуют у животных. Общим адаптационным механизмом является цикл отрицательной обратной связи, который отбирает определенный биохимический сигнал и контролирует процесс адаптации.
Адаптация меняется в зависимости от интенсивности и продолжительности входного сигнала. Вероятно, более сильный или продолжительный входной сигнал вызывает выраженные адаптивные изменения, и часто продолжительную адаптацию. Подобным образом клетки могут модулировать адаптацию, поскольку она определяется последовательными независимыми механизмами, каждый из которых обладает собственной чувствительностью и кинетическими параметрами.
Прекрасным примером адаптивного механизма является G-белок. На рисунке ниже показано, что самым ранним этапом в цепи процессов адаптации служит фосфорилирование рецептора, которое катализируется киназой рецептора, сопряженной с G-белком (GRK). Киназа избирательно узнает активную конформацию комплекса лиганд-рецептор. Фосфорилирование блокирует способность рецептора активировать G-белок, а также усиливает связывание аррестина, белка, который еще больше блокирует активацию. Более того, связывание аррестина делает рецепторы чувствительными к эндоцитозу, в результате чего они удаляются с поверхности клеток. Эндоцитоз также служит первым шагом на пути протеолиза рецепторов.
Наряду с этими прямыми эффектами, многие гены рецепторов регулируют свою транскрипцию по механизму обратной связи так, что сигнал, исходящий от рецептора, приводит к снижению его экспрессии.
Таким образом, стимуляция вызывает множество адаптивных процессов, которые происходят либо сразу же (фосфорилирование, связывание аррестина), либо наступление их задерживается (регуляция транскрипции) и которые носят обратимый и необратимый характер. Этот набор адаптивных реакций существует для многих рецепторов, сопряженных с G-белками, и многие клетки могут использовать весь набор с тем, чтобы контролировать выход сигнала с одного рецептора. При этом скорость и степень адаптации, а также ее обратимость выбираются в соответствии с программой развития клетки.
В клетке адаптивные процессы могут изменяться в количественном и качественном отношении при изменении точек приложения действия механизмов обратной связи, которые расположены на пути передачи сигнала. В линейной цепи событий передачи сигналов изменения этих точек приводят к изменениям кинетики или степени адаптации. В разветвленной системе изменения этих точек определяют, является ли адаптация уникальной к одному входному сигналу или развивается по отношению к нескольким близким входным сигналам.
Если при активации рецептора происходит его прямая десенсибилизация или она запускается каким-то дальнейшим событием на неразветвленном пути, то изменяться будут только те сигналы, которые генерируются этим рецептором. Адаптация, селективная в отношении определенного рецептора, называется гомологичной адаптацией.
Наоборот, контроль с помощью обратной связи может затрагивать конвергентные пути, отдаленные от нескольких рецепторов. Таким образом, регулируется активность как инициирующего, так и и других рецепторов. При такой гетерологичной адаптации в данной контрольной точке регулируются все возможные входные сигналы. Общим примером служит фосфорилирование рецепторов, сопряженных с G-белком под действием или протеинкиназы А, или протеинкиназы С. Эти киназы активируются цАМФ или Са2+ плюс липид диацилглицерин соответственно. Так же как GRK, киназы ослабляют активность рецептора и способствуют связыванию аррестина.
Реакция клетки на входящий сигнал может меняться в зависимости от состояния ее геомеостаза. В данном случае под гомеостазом понимают фазу цикла, метаболический статус и другие аспекты физиологического состояния клетки. Опять-таки все адаптивные процессы могут проявляться в большей или меньшей степени в разных клетках, для различных внутриклеточных процессов, или в различные периоды их существования.
Наверху: при получении сигнала чувствительность путей его передачи регулируется таким образом,
чтобы клетка могла приспособиться к новому уровню входного сигнала.
Таким образом, после начальной стимуляции, ответ клетки становится более слабым. Если для восстановления системы не прошло достаточно времени,
второй подобный сигнал вызовет слабый ответ.
Внизу: для некоторых адаптационных механизмов обратная связь эффективна только по отношению к рецептору, который активируется, и не влияет на параллельные пути передачи сигналов.
Такие механизмы называются гомологичными.
Слева: агониста рецептора R1 может инициировать одно из двух событий обратной связи, которые приводят к десенсибилизации R1.
В остальных случаях получение клеткой сигнала будет также вызывать десенсибилизацию параллельных или близких систем их передачи.
Справа: агонист а инициирует десенсибилизацию R1 и R2. Ответ на агонист b, который связывается с R2, также десенсибилизирован.
Такая гетерологичная десенсибилизация представляет собой обычное явление. При получении клеткой сигнала в ней запускаются многочисленные процессы адаптации.
Как правило, их компоненты существуют в клетке. Обычно они запускаются в определенной последовательности, в соответствии с продолжительностью и интенсивностью сигнала.
Для GPCR известно, по крайней мере, пять механизмов десенсибилизации; другие механизмы действуют на уровне G-белка и эффекторов.
Читайте также:
- Галактоземия. Врожденные нарушения обмена аминокислот
- Острое облучение радиоактивным йодом. Влияние радиоактивного йода на щитовидную железу
- Рентгенограмма, КТ, МРТ при остеопорозе позвоночника
- Влияние этаминал-натрия, скополамина на условные рефлексы. Рефлекторные эффекты диазепама
- Пример отогенного синустромбоза. Отогенный сепсис