Нуклеосома как структурная единица хромосомы
Добавил пользователь Владимир З. Обновлено: 07.01.2026
В прокариотических клетках наследственный материал представлен только молекулой ДНК, чаще кольцевой. В эукариотических клетках ДНК в хромосоме существует в комплексе с другими химическими компонентами и в первую очередь с белками. Этот комплекс получил название хроматин. Большая часть хроматина представлена белками (до 65%), ДНК составляет около 35%. Кроме белков и ДНК в хроматин входит: РНК, липиды, углеводы, ионы металлов (Ca, Mg) и некоторые другие компоненты, но эти вещества находятся в хроматине в малом количестве.
Таким образом, хроматин – это сложная многокомпонентная система. Важную роль в хроматине играют белки. Среди белков хроматина выделяют негистоновые белки и белки гистоны, которые составляют до 80% всех белков хроматина. Гистоны небольшие белки с очень высоким содержанием положительно заряженных аминокислот (лизина и аргинина). Суммарный положительный заряд позволяет гистонам прочно связываться с ДНК. Выделяют 5 типов гистонов: H1; H2A; H2B; H3; H4. Особенностью генов этих белков является отсутствие в их последовательностях интронов. Белки гистоны не только выполняют структурную функцию в хроматине, но и участвуют в механизмах функционирования наследственного материала. Негистоновые белки выполняют структурную, регуляторную и ферментативную функцию в хроматине.
Таким образом, хроматин следует рассматривать как вещество хромосом. Отдельные хромосомы представляют собой хроматин с одной молекулой ДНК и определенной структурной организацией.
Структурная организация эукариотической хромосомы достаточно сложная. Эта сложность связана с тем, что длина молекулы ДНК в составе хромосомы во много раз превышает размер самой хромосомы. В среднем одна метафазная хромосома человека размером 5 мкм содержит молекулу ДНК длинной 5 см., то есть ДНК превышает размер хромосомы примерно в 10 тысяч раз. Чтобы молекула ДНК могла уместиться в хромосоме она должна компактно укладываться в ней, поэтому при формировании хромосомы имеет место компактизация ДНК в ней. Выделяют пять уровней упаковки (компактизации) ДНК в хромосоме или пять уровней структурной организации хромосом.
Первый уровень – это формирование нуклеосом. Формирование нуклеосом можно представить путём накручивания спирально молекул ДНК на белковый остов, состоящий из 8 молекул гистонов четырёх типов: H2A; H2B; H3; H4 (гистоновый октамер, белковый кор). При этом молекула ДНК делает около двух витков вокруг белкового остова и переходит на другой остов. Так формируется повторяющаяся структурная единица хромосомы – нуклеосома (рис. 6).
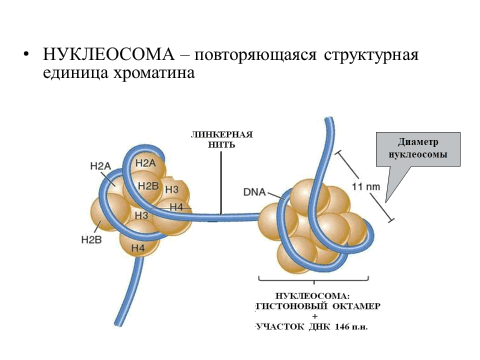
Рис. 6. Нуклеосома
Участки ДНК, расположенные между нуклеосомами, называют линкерной (связывающей) ДНК. Расположенные рядом друг с другом нуклеосомы образуют довольно толстую нить, которая напоминает «бусины на нитке». При этом происходит 7-кратное укорочение длины молекулы ДНК. Нуклеосома рассматривается как универсальная структурная единица хроматина (хромосомы).
На втором уровне компактизации ДНК нуклеосомная нить закручивается в спираль с образованием соленоидо-подобной структуры (фибриллы). Стабильность этого уровня организации хромосомы обеспечивается гистоном H1, который «сшивает» витки ДНК соленоида. Диаметр хроматиновой фибриллы составляет 30 нм. При этом длина нити ДНК сокращается в 50 раз от исходной (рис. 7).

Третий уровень компактизации ДНК – это формирование хроматиновых петель (рис. 8). Хроматиновые соленоидо-подобные фибриллы укладываются в виде петель, диаметр фибриллы увеличивается до 300 нм. Степень компактизации при этом составляет 10 3 , первоначальная длина молекулы ДНК сокращается в 1000 раз. Таким образом, в результате спирализации ДНК и взаимодействия с белками образуется три уровня структурной организации хромосом. Все эти три уровня компактизации ДНК в хромосомах имеют место в интерфазу митотического цикла.
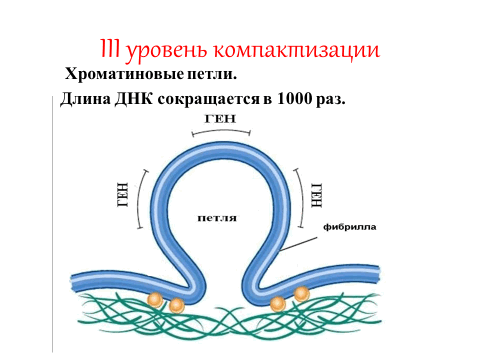
Рис. 8. Хроматиновая петля
На следующем четвёртом уровне компактизации ДНК фибриллы с хроматиновыми петлями многократно складываются и превращаются в хроматиды диаметром 700 нм. Длина молекулы ДНК в хроматиде сокращается в 2-3 тысячи раз от исходной.
На пятом уровне наблюдается максимальная степень компактизации ДНК с образованием метафазных хромосом. Благодаря такой суперспирализации достигается плотная упаковка наследственного материала, что важно при расхождении хромосом в митозе. При такой упаковке происходит укорочение метафазных хромосом по сравнению с размерами заключённой в них молекулы ДНК примерно в 10 тысяч раз.
В метафазу хромосомы имеют вид цилиндрических телец, которые интенсивно окрашиваются основными красителями. В метафазной хромосоме можно выделить центромеру (первичную перетяжку), которая делит хромосому на два плеча (рис. 9).
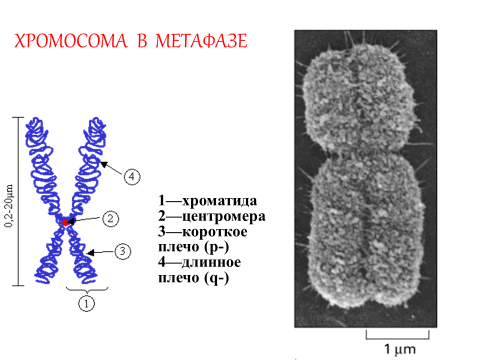
Рис. 9. Метафазная хромосома
Центромеры, которые разделяют хромосомы на плечи, выполняют важные функции. Они обеспечивают: удержание хромосом; правильность выстраивания хромосом по экватору в метафазу; прикрепление к веретену деления; отвечают за контроль наступления анафазы. В центромерах локализованы повторяющиеся последовательности нуклеотидов генома.
В зависимости от положения центромеры хромосомы человека делятся на четыре типа: 1) метацентрические хромосомы, обладающие плечами примерно равной длины (равноплечные); 2) субметацентрические хромосомы с плечами неравной длины (умеренно неравноплечные); 3) очень субметацентрические хромосомы (выражено неравноплечные); 4) акроцентрические хромосомы с очень коротким вторым плечом. Короткое плечо хромосомы принято обозначать p-, длинное плечо q-.
Некоторые хромосомы содержат вторичные перетяжки. Часть хромосомы, которая отделяется вторичной перетяжкой получила название – спутник. Хромосомы, которые имеют спутник называют спутничными (рис. 10).
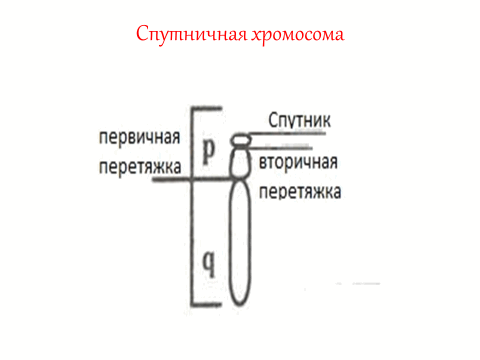
Рис. 10. Спутничная хромосома
Спутничными хромосомами у человека являются 13-15, 21-22 хромосомы. В коротких плечах спутничных хромосом локализованы ядрышко-образующие районы хромосом (ЯОР). На препаратах метафазных хромосом активные ЯО-районы выявляются как вторичные перетяжки и окрашиваются азотнокислым серебром. В ЯО-районах хромосом образуются ядрышки.
Особое значение имеют концевые участки хромосом, получившие название теломеры (рис. 11). Они расположены на обоих концах линейной хромосомы. В теломерных районах хромосом локализованы в основном двунитевые некодирующие повторы (ТТАГГГ)n, заканчивающиеся 3′-однонитевым участком. Основными функциями теломер являются: сохранение целостности хромосомы; обеспечение её полной репликации. Кроме того, теломеры: а) участвуют в прикреплении хромосом к ядерному матриксу; б) способствуют конъюгации хромосом и кроссинговеру в профазу мейоза I; в) способствуют удержанию сестринских хроматид в метафазу митоза.

Рис. 11. Теломерные участки хромосом
В интерфазных хромосомах сохраняются только три первых уровня компактизации хроматина, поэтому интерфазный хроматин гораздо более растянут по сравнению с хроматином, который имеется во время митоза. Это связано с некоторой деспирализацией, менее плотной упаковкой хромосом в интерфазу. Нитевидные хромосомы в интерфазу свободно располагаются в ядерном соке, переплетаются между собой и различить каждую хромосому становится невозможным. Однако степень спирализации, уплотнения как отдельных хромосом, так и отдельных участков хромосом не одинакова. Выделяют гетерохроматиновые (гетерохроматин) и эухроматиновые (эухроматин) области хромосом. Гетерохроматин – плотные, компактные (конденсированные) районы хромосом, интенсивно окрашиваются и могут быть видны в световой микроскоп. Выделяют два вида гетерохроматина:
1) структурный (конститутивный) гетерохроматин; 2) факультативный гетерохроматин. Структурный гетерохроматин стабильно выявляется в хромосомах в зоне центромеры. Он в целом генетически инертный, так как не содержит активно транскрибируемых генов.
Факультативный гетерохроматин – это гетерохроматизированные эухроматические районы хромосом. Примером факультативного гетерохроматина является тельце полового хроматина (тельце Барра), которое образуется в клетках гомогаметного пола (у женщин). При этом одна из двух Х-хромосом сворачивается, плотно укладывается и хорошо прокрашивается.
Эухроматин – это менее конденсированные районы хромосом. Обычно эухроматин не виден в световой микроскоп, либо виден в виде слабоокрашенных структур. Эухроматин транскрипционно активен, содержит функционирующие гены.
Итак, у эукариот ДНК сосредоточена в хромосомах. В ядре клеток человека 46 хромосом, из них 22 пары аутосом и пара половых хромосом. Хромосомы, которые относятся к одной паре называют гомологичными. Гомологичные хромосомы абсолютно аналогичны. Число хромосом в клетках организма – это видовой признак. Кроме количества хромосомы каждого вида характеризуются определенным размером, формой, деталями строения. Хромосомный набор клеток данного вида организмов с характерным числом, морфологическими особенностями отдельных хромосом называется кариотипом.
Хромосомы человека не очень легко отличить одну от другой. С целью унификации методов идентификации хромосом человека в 1960 г на конференции цитогенетиков в г. Денвере (США) была принята классификация, учитывающая величину хромосом и расположение центромер. В том же году К. Патау дополнил эту классификацию и предложил разделить 23 пары хромосом человека на 7 групп от А до G (табл. 1).
Нуклеосома как структурная единица хромосомы
Нуклеосома как структурная единица хромосомы
• При действии микрококковой нуклеазы из хроматина высвобождаются индивидуальные нуклеосомы, представляющие собой частицы размером 11S
• Нуклеосома содержит примерно 200 пн ДНК, по две копии каждой из молекул гистонов Н2А, Н2В, Н3 и Н4, а также одну копию молекулы гистона Н1
• ДНК обернута по поверхности октамера белков
Хроматин и хромосомы состоят из дезоксирибонуклеопротеиновой фибриллы, которая имеет несколько уровней организации. Структура митотических хромосом с их характерной дифференциальной окраской соответствует наиболее сложному, полностью конденсированному состоянию этой фибриллы. У всех эукариот основная структурная единица хроматина одинакова — это нуклеосома, содержащая около 200 пн ДНК и гистоны. Негистоновые белки принимают участие в скручивании цепочки нуклеосом в структуру более высокого порядка.
Когда интерфазные ядра суспендируют в растворе низкой ионной силы, они набухают и разрушаются. При этом высвобождаются фибриллы хроматина. На рисунке ниже представлен лизат ядер, содержащий вышедший из них хроматин. Отдельные участки состоят из плотно упакованных фибрилл, однако видно, что области, содержащие релаксированный хроматин, содержат дискретные частицы. Это нуклеосомы. В полностью деспирализованных участках отдельные нуклеосомы соединены между собой тонкой нитью двойной цепи ДНК. Эта непрерывная двойная цепь проходит через группу частиц.
Индивидуальные нуклеосомы можно получить, обрабатывая хроматин одной из эндонуклеаз — микрококковой нуклеазой. Она разрезает нити ДНК между нуклеосомами. Вначале высвобождаются группы нуклеосом, а затем отдельные нуклеосомы. Изолированные нуклеосомы выглядят как компактные частицы. Они обладают коэффициентом седиментации ~11S.

При переваривании хроматина с помощью микрококковой нуклеазы
высвобождаются индивидуальные нуклеосомы.
Нуклеосома содержит 200 пн ДНК, связанной с октамером гистонов, который состоит из двух копий молекулы каждого гистона Н2А, Н2В, Н3, и Н4. Эти гистоны называются коровыми (англ, core — сердцевина). Расположение их в нуклеосоме схематически показано на рисунке ниже. Эта модель объясняет стехиометрию коровых гистонов в хроматине: Н2А, Н2В, Н3, и Н4 присутствуют в эквимолекулярных количествах, по 2 молекулы каждого белка на 200 пн ДНК.
Гистоны Н3 и Н4 принадлежат к числу наиболее консервативных из известных белков. Это позволяет предполагать, что они обладают одинаковыми функциями в клетках всех эукариот. Гистоны Н2А и Н2В обнаружены во всех эукариотических клетках, однако отмечены существенные видоспецифические различия в их первичной структуре.
Гистон Н1 включает группу близких по структуре белков, которые показывают выраженную тканевую и видовую вариабельность (и не обнаружены у дрожжей). Функция гистона Н1 отличается от функции коровых гистонов. Этот гистон присутствует в хроматине в половинном от коровых гистонов количестве и может быть удален из него наиболее полно (обычно с помощью экстракции разбавленным (0,5 М) солевым раствором). Удаление гистона Н1 не нарушает структуру нуклеосомы. Это позволяет предполагать, что гистон располагается вне нуклеосомной частицы.
По форме нуклеосома соответствует плоскому диску или цилиндру диаметром 11 нм и высотой 6 нм. Длина ДНК примерно в два раза превышает ~34 нм окружность нуклеосомы. ДНК проделывает симметричный путь вокруг октамера. На рисунке ниже схематически представлен путь ДНК в виде спирали, делающей два оборота вокруг цилиндрического октамера. Заметьте, что ДНК «входит» на поверхность нуклеосомы и «покидает» ее в точках, расположенных близко друг к другу. В этой области может располагаться гистон Н1.
На поперечном сечении предложенной модели мы видим, что витки двух оборотов нити ДНК лежат вплотную друг к другу. Высота цилиндра 6 нм, из них 4 нм занято двумя витками ДНК (диаметр нити ДНК составляет 2 нм).
Наличие на нуклеосоме двух витков ДНК, возможно, имеет функциональное значение. Поскольку один виток вокруг нуклеосомы составляет 80 пн ДНК, две точки, разделенные расстоянием 80 пн в свободной двойной спирали, в действительности, на поверхности нуклеосомы могут располагаться близко друг к другу, как это показано на рисунке ниже.
Хроматин, который высвобождается при лизисе ядер,
состоит из компактно организованных дискретных частиц. Нуклеосомы состоят из примерно одинаковых количеств ДНК и гистонов (включая гистон Н1).
Рассчетная маса нуклеосомы составляет 262 кДа Два витка ДНК на нуклеосоме расположены близко друг к другу. Нуклеосома может представлять собой цилиндр,
снаружи которого располагаются два витка ДНК. Последовательности ДНК, локализованные на разных витках,
на нуклеосоме могут располагаться близко друг к другу.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Научная электронная библиотека

Геном человека – термин, используемый при описании всей генетической информации, закодированной в виде ДНК клеток человека. Клетка человека имеет два генома: сложный ядерный геном (хромосомная ДНК), содержащий более 99,9995 % всей генетической информации, и простой митохондриальный геном, в составе которого находится менее 0,0005 % ДНК. Ядерный геном распределён между 24-мя различными двуспиральными молекулами ДНК, которые в комплексе с различными гистоновыми и негистоновыми белками формируют хромосомы человека. С молекулярной точки зрения каждая хромосома клетки является сложно организованной структурой. Диплоидная клетка человека с интерфазным ядром при диаметре 5–10 мкм содержит около двух метров последовательностей молекул ДНК, которые образуют 23 пары хромосом. Гаплоидный геном человека, который характеризует половые клетки с хромосомами Х или Y, включает 23 различные хромосомы – 22 аутосомы и одну половую хромосому (гоносому) в зависимости от пола: Х – женский или Y – мужской пол. Индивидуальные хромосомы, как было сказано выше, различаются своими морфологическими характеристиками. В среднем, хромосома человека содержит около 130 млн пн, однако содержание ДНК в разных хромосомах варьирует от 40 до 260 млн пн.
Как уже сказано выше, данные о структуре, размерах и количестве митотических хромосом в клеточном ядре определяются как кариотип. Морфологическое строение хромосом на всех уровнях организации (молекулярном, микроскопическом и субмикроскопическом) определяется упаковкой нитей ДНК, организующих хромосому. На микроскопическом уровне нити ДНК формируют спирализованные хромонемы митотических хромосом. Упаковка хромонем по длине хромосом неравномерна. В них закономерно дифференцируются участки с резко выраженной и резко сниженной степенью спирализации хромонем. Последние формируют хромосомные перетяжки. Важным элементом структуры хромосом является первичная перетяжка, в участке которой расположена центромера. Она делит хромосому на два плеча – короткое (р) и длинное (q).
Структура хромосом претерпевает значительные изменения в ходе клеточного деления (клеточного цикла) и, следовательно, не является постоянной. Хромосомы интерфазных ядер (интерфазные хромосомы) в отличие от метафазных хромосом (см раздел, посвященный делению клетки) представляют собой более расправленные и диффузные клеточные структуры. Интерфазные хромосомы содержат одну хроматиду, в составе которой имеется одна двуспиральная молекула ДНК, в то время как метафазные хромосомы образованы двумя хроматидами и двумя молекулами ДНК. Хромосома, как функционирующая клеточная органелла, должна содержать минимум три типа последовательностей ДНК, формирующих её структурные компоненты: центромеру, теломеры и участок начала репликации ДНК.
Индивидуальные хромосомы содержат одну протяжённую двуспиральную молекулу ДНК диаметром 2 нм, которая претерпевает несколько уровней упаковки, формируя хроматин или ДНК-белковый комплекс (дезоксирибонуклеопротеид – ДНП). Элементарной дискретной единицей упаковки хроматина является нуклеосома. Каждая нуклеосома имеет вид глобулы и состоит из четырех пар гистоновых белков Н2А, Н2В, Н3 и Н4, содержащих по 102–135 аминокислотных остатков. Вокруг гистоновой глобулы диаметром 10 нм (октамера) располагается молекула двуспиральной ДНК длиной 146 пн (или 68 нм), образуя виток в 1,75 оборота. Соседние нуклеосомы соединяются так называемой «спейсерной» ДНК длиной около 20 нм, образуя структуры в виде «бус на нитке». На следующем этапе компактизации 10-нанометровые нуклеосомы образуют фибриллы хроматина диаметром 30 нм (в каждом витке примерно 6 нуклеосом), которые образуют большие петли. На стадии метафазы степень компактизации достигает 1 на 10000 от первоначальной длины хромосомной ДНК. Петли 30-нанометровых фибрилл хроматина содержат 20–100 тысяч пн. ДНК на каждой петле прикрепляется к осевой структуре хромосомы (рис. 4). Осевые компоненты метафазной хромосомы состоят из негистоновых белков.
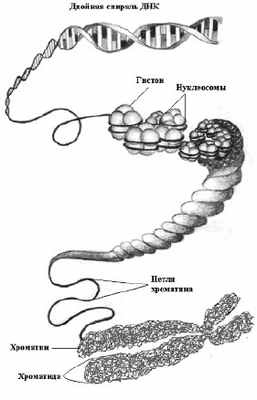
Рис. 4. Организация ДНК
Как уже отмечалось, одним из структурных элементов хромосомы является центромера – место присоединения кинетохоров (макромолекулярных структур на белковой основе, участвующих в процессе расхождения хромосом при митотическом делении клетки), и слипания сестринских хроматид. Они также регулируют присоединение хромосом к веретену деления и направляют движение хромосом в ходе мейотического и митотического деления клетки. Хромосомы без центромеры (ацентрические фрагменты) не могут присоединяться к нитям веретена деления и теряются в ходе клеточного деления. У человека центромеры содержат особые типы повторяющихся ДНК, а также специфические «центромерные» белки, обеспечивающие сегрегацию хромосом в ходе митотического и мейотического деления клеток. В зависимости от положения центромеры в хромосоме, как уже сказано выше, она может быть метацентрической, субметацентрической и акроцентрической. В акроцентрических хромосомах довольно часто на коротких плечах видны небольшого размера хроматические тельца, так называемые спутники, и спутничные нити (рис. 5). Они содержат ядрышковый организатор – значимую структуру генома человека. Плечи некоторых хромосом человека содержат вторичные перетяжки.
Рис. 5. Схематическое изображение метафазных хромосом:
1 – метацентрическая хромосома; 2 – субметацентрическая хромосома; 3 – акроцентрическая хромосома. Видны: а – центромеры; б – длинные плечи (q); в – короткие плечи (p); г – теломерные участки; д – спутники; е – спутничные нити
Другими структурными элементами хромосом являются теломеры. Это специализированные структуры, содержащие особые типы ДНК и белки, которые образуют концевые участки хромосом. Теломеры выполняют несколько функций:
1) поддержание структурной целостности хромосомы;
2) обеспечение полной репликации концевых участков хромосомы;
3) поддержание организации хромосом в интерфазном ядре.
Теломеры хромосом человека представляют собой повторы нуклеотидной последовательности ТТАГГГ, общий размер которой варьирует от 3 до 20 тысяч пн. Стабильность теломер поддерживается с помощью фермента теломеразы, нарушение функциональной активности которой приводит к нарушению структуры хромосом и, как следствие, к клеточной гибели. Укорочение последовательностей теломер, которому противодействует теломераза, связано с процессами старения и малигнизации.
ДНК хромосом реплицируется в ходе периода синтеза ДНК клеточного цикла. Каждая хромосома содержит множество элементарных единиц репликации – репликонов, представляющих собой участки автономной репликации. Каждый репликон имеет одну точку инициации репликации, с которой начинается двунаправленный синтез ДНК (см предыдущий раздел, посвященный синтезу ДНК). Размеры репликонов могут различаться друг от друга. Репликоны у человека могут иметь большие размеры, достигая более чем 1 млн пн.
Нуклеосома как структурная единица хромосомы
преподаватель - д.б.н., профессор И.И. Киреев
Ядерные компоненты бактерий: нуклеоид, его химический состав, структура нуклеоида, петлевые домены, единица репликации - репликон, механизм расхождения нуклеоидов после репликации.
Ядро эукариот: компоненты, структурные типы интерфазных ядер (диффузные, хромоцентрические, хромонемные, политенные).
Хроматин. ДНК хроматина, линейность, гетерогенность, кинетика реассоциации ДНК, изохоры, уникальные последовательности нуклеотидов, умеренно и высокоповторяющиеся последовательности нуклеотидов, сателллитные ДНК, функциональные элементы хромосомных ДНК: участки независимой (автономной) репликации, центромерные ДНК, теломерные ДНК, роль теломеразы, MAR, SAR- участки ДНК, связанные с ядерным белковым матриксом. Феномен синтении.
Гетерохроматин и эухроматин, их функциональное значение и структуризация.
Синтез ДНК хроматина: полирепликонность, кластеры репликации ДНК (реплисомы), асинхронность репликации по длине хромосомы, репликация участков уникальных последовательностей и повторов, репликация гетерохроматина и эухроматина, причины блокады повторной репликации.
Белки хроматина: гистоны и негистоновые белки (ферменты, факторы, HMG- , белки ядерного матрикса). Гистоны: общая характеристика и свойства, консервативность состава, характер ассоциации с ДНК, типы гистонов, их функциональная и структурная роль, модификации гистонов в связи с активацией хроматина.
Уровни компактизации ДНК.
Нуклеосомный уровень. Характеристика нуклеосомы, спейсеры и их величина, динамика построения нуклеосомы, поведение нуклеосом при репликации и транскрипции. Модификации N-концевых участков молекул гистонов, их значение.
30 нм- фибрилла - основной нативный компонент хроматина, общая характеристика, суперсоленоидная модель, нуклеомерная модель, состав нуклеомера, его характеристики, роль гистона H1 в поддержании структуры 30 нм - фибриллы.
Третий уровень компактизации ДНК- петлевые домены. Получение “нуклеоидов” интерфазных ядер, величина петель ДНК “нуклеоидов”, модели петлевых доменов, “розетки” в составе хроматина при его декомпактизации, размер петель в “розетках” и количество в них ДНК, розетки в составе интерфазных ядер, политенных и митотических хромосом, понятие “хромомер”, примеры их обнаружения в естественных условиях. Роль негистоновых белков в поддержании структуры петлевых доменов.
Хромонемный уровень укладки фибрилл хроматина. Хромонема в интерфазных ядрах, хромонема в профазе и телофазе митоза и при искусственной декомпактизации хромосом.
Структура митотических хромосом. Фазы митоза, хромосомный цикл.
Продольная неоднородность хромосом: G(Q), R,C бэнды, их химическая природа, методы дифференциальной окраски хромосом, химические особенности различных бэндов; искусственная дифференцированная деконденсация хромосом – причина дифференциальной окраски, этапы деконденсации хромосом, обратимость и стабилизация деконденсированного состояния хромосом.
Уровни структурной организации митотических хромосом: петлевые домены, белковый матрикс митотических хромосом, иерархия уровней компактизации ДНК.
Локализация хромосом в интерфазном ядре: Хромосомные территории. Тельца Барра, локализация центромер и теломер, локализация по Раблю, распределение меченного тимидина в ряду клеточных делений, трехмерная локализация в ядрах политенных хромосом, метод флуоресцентной in situ гибридизации (FISH)..
Продукты ядерной активности. Типы синтезированных РНК, коротко и долго живущие РНК, синтез РНК, транскрипционная единица, типы РНК-полимераз. Малые ядерные РНК и РНП, их синтез, функции и значение для клетки.
Особенности синтеза и-РНК: структура гена, интроны и экзоны, синтез гетерогенных ядерных РНК, сплайсинг, сплайсосомы, малые ядерные РНП, интерхроматиновые гранулы, перихроматиновые гранулы, информофоры и информосомы, морфология транскрипции и-РНК, синтез и-РНК в кольцах Бальбиани политенных хромосом, разные типы активности транскрипционных единиц.
Синтез т-РНК: полицистронность участков синтеза т-РНК, предшественник, процессинг и образование зрелых т-РНК.
Синтез р-РНК: структура р-гена, консервативность состава р-РНК, полицистронность р-генов, их кластерность, локализация в районах ядрышковых организаторов, строение транскрипционных единиц, синтез предшественника, его процессинг, образование четырех типов р-РНК и их участие в структуре субъединиц рибосом, образование рибосом.
Ядрышко – хромосомный локус синтеза р-РНК и рибосом: ядрышковый организатор, число ядрышек в ядре, амплификация ядрышек. Строение ядрышек: фибриллярные центры, плотный фибриллярный компонент, гранулярный компонент; количество и структура этих компонентов в зависимости от функциональной нагрузки ядрышка, ферменты и ядрышковые белки при синтезе рибосом; судьба ядрышковых компонентов при митозе; периферический хромосомный материал; пред-ядрышковые структуры, локализация белков ядрышка и их судьба во время митоза.
Ядрышко как мультифункциональный компартмент, нетрадиционные функции ядрышка. Малые ядрышковые РНП и их функции.
Специальные ядерные компартменты: тельца Кахаля, PLM- структуры, околоядрышковый хроматиновый компартмент, их функции.
Ядерный белковый матрикс. Способы выявления ядерного белкового матрикса (ЯБМ), его компоненты и биохимический состав; ламина и ламины, их участие в связывании с периферическим хроматином; ДНК в составе ЯБМ, РНК в составе ЯБМ; белки ЯБМ; роль его в синтезе ДНК и РНК, распределение компонентов ЯБМ во время митоза.
Ядерная оболочка. Строение и состав: внешняя ядерная мембрана, перинуклеарное пространство, внутренняя ядерная мембрана, ламина, комплексы ядерных пор; ламины и их роль в связывании фибрилл хроматина с ламиной, гранулярный периферический слой хроматина, механизм связи хроматина с ядерной оболочкой.
Ядерные поры: комплекс ядерной поры (КЯП), строение, химический состав, число ядерных пор; участие КЯП в ядерно-цитоплазматических связях, импорт белковых молекул, рецепторы импортинов, роль ПЯЛ в транспорте нуклеофильных белков, механизмы транслокации через ядерную пору; ядерный экспорт: экспортины и контроль за выходом из ядра белков, РНП и рибосом, особенности транспорта и-РНП; судьба ядерной оболочки во время митоза, модификация ламинов, образование микроядер.
Ядерная технология: получение гетерокарионов, получение преждевременно конденсированных хромосом, получение микроядер, микрохирургические и лучевые манипуляции с ядерными компонентами, пересадки ядер, молекулярная гибридизация in situ.
1.Функциональная архитектура ядра. Основные функциональные компартменты ядра эукариотической клетки. Сайты репликации, их динамика в S-периоде. Ядрышко, сплайсосомы, тельца Кахаля, PML- тельца и др. Характеристики их белкового состава, функции
2.Хромосомные территории и регуляция активности генов. Способы визуализации хромосомных территорий. Различные теории взаимодействия хрс территорий в ядре.
Зависимость топологии отдельных хромосом в ядре от активности содержащихся в них генов.
3. Гистоны и их модифицированные формы. Общая архитектура строения нуклеосомы. Способы и сайты ковалентных модификаций N-концевых участков различных гистонов. Роль модифицированных гистонов в эпигенетической регуляции транскрипции и др. процессов.. Гистоновый код.
4. Многофункциональное ядрышко. Канонические и неканонические функции ядрышка. Ядрышко в митозе, поведение ядрышкового организатора. Ядрышко в интерфазе, процессинг различных РНК. Белки ядрышка и их функции. Взаимодействие ядрышка с другими функциональными компартментами ядра.
5.Ламина и ее связь со структурой и функцией хроматина. Структура ламины, белки, заякоривающие ее структуры во внутренней ядерной мембране. LBR- комплекс, белки группы LAP. Взаимодействие ламины с хроматином. Регуляторные аспекты взаимодействия ламины с пристеночным гетерохроматином.
6.Комплекс ядерной поры. Морфология комплекса ядерной поры (КЯП) по данным, полученным различными методами. Белки КЯП, их взаимодействие с ламиной и оболочкой ядра. Функционирование КЯП, его участие в сегрегации и активном транспорте макромолекул. Белки, обеспечивающие эти процессы.
7.Хроматин и транскрипция. Отличительные черты транскрипционно активного хроматина, модификации гистонов в этом процессе. Ремоделлинг хроматина, сопровождающий транскрипцию. Поведение нуклеосом в процессе транскрипции. Транскрипция и высшие уровни упаковки хроматина.
8.Малые ядерные РНК и их функции. Виды малых ядерных РНК, их размеры. Приуроченность отдельных видов мяРНК к определенным типам клеток. Происхождение мяРНК в клетке – метаболические пути их образования. Транскрипция прицентромерного гетерохроматина. Явление РНК-интерференции. Функции мяРНК, регуляторная роль и их значение при внутриклеточной инактивации вирусной агрессии.
9. Центромера. Центромера, как специфический локус хромосом высших организмов. История изучения. Локализация на хромосоме, состав ДНК, специфические белки. Варианты строения ЦМ у разных организмов. Консервативность строения. Взаимодействие ЦМ с веретеном деления и поведение ее в митозе. Образование и элиминация ЦМ, неоцентромеры.
10 Теломера. История изучения, проблема концевой репликации. Структура Т, состав ее ДНК, консервативность. Пространственная структура Т по современным данным. Специфические белки Т., их свойства. Теломеразный комплекс, его функции. Теломера, старение и раковый рост.
11.Негистоновые белки хроматина. Группы негистоновых белков хроматина. Общие функции. HMG-белки, конденсины, когезины – функционирование на различных стадиях клеточного цикла.
12.Ядерный белковый матрикс (ЯБМ). История изучения. Способы изучения ЯБМ, его визуализации для ультраструктурного исследования. Структуры, входящие в ЯБМ, белки ЯБМ. Специфические участки ДНК , связанные с ЯБМ. Взаимодействие ЯБМ и хроматина. Структура нативного ЯБМ в живой клетке, его изменения в процессах функционирования хроматина. Соотношение ЯБМ и «скэффолда» митотических хромосом.
13. Уровни упаковки хроматина в интерфазном ядре и митотических хромосомах. Факторы, поддерживающие различные уровни упаковки хроматина, начиная с нуклеосомного уровня до уровня митотической хроматиды. Элементарная 30нм фибрилла хроматина. Возможное участие негистоновых белков в упаковке хроматина на разных уровнях. Хромонема, ее выявление и свойства.
14.Современные модели строения митотической хромосомы. Сопоставление моделей укладки хроматина в митотическую хромосому с данными о структуре и свойствах различных видов хроматина: эухроматин, факультативный гетерохроматин, конститутивный гетерохроматин. Корреляция плотности укладки отдельных районов митотических хромосом с данными дифференциальной окраски хромосом. Причины дифференциальной конденсации хромосом.
Чтение курса «Кариология» проходит параллельно с практическими занятиями по той же теме в рамках большого практикума, где студенты самостоятельно получают препараты и исследуют в световой и электронный микроскопы ядра и хромосомы, полученные из различных объектов. В ходе обсуждения практических заданий и их выполнения проходит промежуточная аттестация студентов по отдельным темам курса.
Итоговая аттестация (вопросы)
1.Функциональная архитектура ядра. Основные функциональные компартменты ядра эукариотической клетки. Сайты репликации, их динамика в S-периоде.
2. Ядрышко, сплайсосомы, тельца Кахаля, PML- тельца и др. Характеристики их белкового состава, функции
3.Хромосомные территории и регуляция активности генов. Способы визуализации хромосомных территорий. Различные теории взаимодействия хрс территорий в ядре.
4. Гистоны и их модифицированные формы. Общая архитектура строения нуклеосомы. Способы и сайты ковалентных модификаций N-концевых участков различных гистонов.
5.Роль модифицированных гистонов в эпигенетической регуляции транскрипции и др. процессов.. Гистоновый код.
6. Канонические и неканонические функции ядрышка. Ядрышко в митозе, поведение ядрышкового организатора.
7.Ядрышко в интерфазе, процессинг различных РНК. Белки ядрышка и их функции. Взаимодействие ядрышка с другими функциональными компартментами ядра.
8.Ламина и ее связь со структурой и функцией хроматина. Структура ламины, белки, заякоривающие ее структуры во внутренней ядерной мембране.
9.Комплекс ядерной поры. Морфология комплекса ядерной поры (КЯП) по данным, полученным различными методами. Белки КЯП, их взаимодействие с ламиной и оболочкой ядра.
10Функционирование комплекса ядерной поры, его участие в сегрегации и активном транспорте макромолекул. Белки, обеспечивающие эти процессы.
11.Хроматин и транскрипция. Отличительные черты транскрипционно активного хроматина, модификации гистонов в этом процессе. Ремоделлинг хроматина, сопровождающий транскрипцию.
12. Малые ядерные РНК и их функции. Виды малых ядерных РНК, их размеры. Приуроченность отдельных видов мяРНК к определенным типам клеток. Происхождение мяРНК в клетке – метаболические пути их образования.
13. Явление РНК-интерференции. Функции мяРНК, регуляторная роль и их значение при внутриклеточной инактивации вирусной агрессии.
14. Центромера, как специфический локус хромосом высших организмов. Локализация на хромосоме, состав ДНК, специфические белки. Варианты строения ЦМ у разных организмов.
15. Взаимодействие центромеры с веретеном деления, ее функции и поведение в митозе. Образование и элиминация ЦМ, неоцентромеры.
16 Теломера. Проблема концевой репликации. Структура Т, состав ее ДНК, консервативность. Пространственная структура Т по современным данным.
17.Специфические белки теломер, их свойства. Теломеразный комплекс, его функции. Теломера, старение и раковый рост.
18.Негистоновые белки хроматина. Группы негистоновых белков хроматина. Общие функции. HMG-белки, конденсины, когезины – функционирование на различных стадиях клеточного цикла.
19.Ядерный белковый матрикс (ЯБМ). Способы изучения ЯБМ, его визуализации для ультраструктурного исследования. Структуры, входящие в ЯБМ, белки ЯБМ. Структура нативного ЯБМ в живой клетке, его изменения в процессах функционирования хроматина.
20. Уровни упаковки хроматина в интерфазном ядре и митотических хромосомах. Факторы, поддерживающие различные уровни упаковки хроматина, начиная с нуклеосомного уровня до уровня митотической хроматиды.
21.Современные модели строения митотической хромосомы. Корреляция плотности укладки отдельных районов митотических хромосом с данными дифференциальной окраски хромосом. Причины дифференциальной конденсации хромосом.
Список учебной литературы
Босток К., Самнер Э. Хромосома эукариотической клетки. “Мир”, 1981.
Жимулев И.Ф. Общая и молекулярная генетика. Изд.2-ое, Новосибирск, 2003
Захаров А.Ф. Хромосомы человека, М., “Медицина”, 1977.
Збарский И.Б. Организация клеточного ядра. М., “Медицина”, 1988.
Збарский И.Б., Кузьмина С.Н. Скелетные структуры клеточного ядра. М., “Наука”,1991.
Коряков Д.Е., Жимулев И.Ф. Хромосомы. Структура и функции. Новосибирск, Изд-во Сибирского отделения РАН, 2009.
Прокофьева- Бельговская А.А. Гетерохроматические районы хромосом. М., «Наука»,1986.
Разин С.В., Быстрицкий А.А. Хроматин: упакованный геном. М., 2009.
Ченцов Ю.С., Поляков В.Ю. Ультраструктура клеточного ядра. М., «Наука», 1974.
Ченцов Ю.С. Введение в клеточную биологию. 4-е изд. М., ИКЦ «Академкнига», 2004.
Alberts B., Johnson A., Lewis J., Raff M., Roberts K., Walter P. Molecular Biology of the Cell. 5-th edition. 2007.
Pollard T., Earnshaw W.C., Lippincott-Schwartz J. Cell Biology. (2-nd edition). 2006.
После прослушивания курса «Кариология» студент должен иметь полное представление о современном состоянии цитологических знаний о структуре и функционировании хроматина и хромосом эукариотической клетки.
Разница между хроматином и нуклеосомой
Видео: Разница между хроматином и нуклеосомой | Сравните разницу между похожими терминами
Содержание:
Ключевое различие - хроматин против нуклеосом
ДНК находится в ядре эукариотических организмов и содержит информацию о наследственности, которая передается следующему поколению. Из-за своей важности ДНК плотно обернута вокруг гистоновых белков и конденсируется в очень стабильную структуру в хромосомах эукариотических клеток, чтобы защитить ее от повреждений. Эта высококонденсированная сложная структура ДНК с гистоновыми белками известна как хроматин. Хроматин состоит из основных структурных единиц, называемых нуклеосомами. Нуклеосому можно определить как небольшую длину ДНК, обернутую вокруг восьми гистоновых белков. Ключевое различие между хроматином и нуклеосомой заключается в том, что хроматин - это целая структура сложной ДНК и белков пока нуклеосома - основная единица хроматина.
СОДЕРЖАНИЕ
1. Обзор и основные отличия
2. Что такое хроматин
3. Что такое нуклеосома
4. Сравнение бок о бок - хроматин против нуклеосом
5. Резюме
Что такое хроматин?
ДНК в ядре не существует в форме свободной линейной цепи. Он связан с белками, называемыми гистонами, и конденсирован в структуру, называемую хроматином. Следовательно, хроматин можно определить как высококонденсированную форму ДНК с гистоновыми белками. Под микроскопом хроматин выглядит как нить, состоящая из гранул, как показано на рисунке 01. Одна гранула известна как нуклеосома, и она является основной структурной единицей хроматина. Хроматин образует хромосомы эукариотических организмов и упакован внутри ядра. Структура хроматина видна только при делении клеток под микроскопом.
Существует две формы хроматина: эухроматин и гетерохроматин. Эухроматин - это менее конденсированная форма хроматина, которая может транскрибироваться в РНК во время экспрессии. Гетерохроматин - это сильно конденсированная форма хроматина, которая обычно не транскрибируется в РНК. Сильно конденсированная суперспиральная ДНК в форме хроматина лучше всего подходит для упаковки внутри ядра, имеющего небольшой объем.
Основная функция хроматина - эффективно упаковывать ДНК внутри ядра, имеющего очень небольшой объем. Хроматины также выполняют дополнительные функции, такие как защита структуры и последовательности ДНК, обеспечивая митоз и мейоз, предотвращая хромосомные разрывы, регулируя экспрессию генов и репликацию ДНК.
Что такое нуклеосома?
Нуклеосома - это небольшой участок хроматина, который обернут вокруг гистонового белка ядра. Это похоже на бусину в ниточке. Гистоновый белок ядра представляет собой октамер, состоящий из восьми гистоновых белков. Две копии каждого гистонового белка находятся в основном октамере. Состав гистонового белка в основном октамере - это H2A, H2B, H3 и H4. Ядро ДНК плотно обвивает глобулярный гистоновый октамер ядра и образует нуклеосому. Затем нуклеосомы выстраиваются в цепочечную структуру и плотно оборачиваются вокруг дополнительных гистоновых белков, чтобы хроматин попал в хромосомы.
Длина основной нити ДНК, которая обвивает октамер гистонов в нуклеосоме, составляет приблизительно 146 пар оснований. Примерный диаметр нуклеосомы составляет 11 нм, а спираль нуклеосом в хроматине (соленоид) имеет диаметр 30 нм. Нуклеосомы поддерживаются дополнительными гистоновыми белками, которые упаковываются в плотно свернутую структуру внутри ядрышка.
В чем разница между хроматином и нуклеосомой?
Хроматин против нуклеосом
Резюме - Хроматин против нуклеосом
Хроматин - это комплекс белков ДНК и гистонов. Он состоит из цепочки нуклеосом, обернутых гистоновыми белками. Нуклеосома - это основная единица хроматина, которая состоит из 147 пар оснований ДНК и восьми гистоновых белков. Цепочка нуклеосом оборачивается гистоновыми белками и конденсируется в высокоорганизованную структуру хроматина, которая является наиболее стабильной формой ДНК, которая упаковывается внутри ядра. В этом разница между хроматином и нуклеосомой.
Ссылки
1. «Структура и функция хроматина: руководство». Структура и функция хроматина: руководство от Abcam. N.p., 20 апреля 2017 г. Web. 20 апреля 2017
2. Люгер, Каролин, Меконнен Л. Дечасса и Дэвид Дж. Треметик. «Новое понимание структуры нуклеосом и хроматина: упорядоченное состояние или беспорядок?» Обзоры природы. Молекулярная клеточная биология. Национальная медицинская библиотека США, 22 июня 2012 г. Интернет. 20 апреля 2017
Изображение предоставлено:
1. «Макроструктура ДНК 0321» от OpenStax (CC BY 4.0) через Commons Wikimedia
2. Предполагается, что "Nucleosome" By Spellcheck. Предполагается, что собственная работа (на основании заявлений об авторском праве) (общественное достояние) через Commons Wikimedia
Читайте также: