Отолитовые рефлексы. Тонические лабиринтные рефлексы
Добавил пользователь Евгений Кузнецов Обновлено: 27.01.2026
При опущенной голове повышается тонус мышц-сгибателей верхних и мышц-разгибателей нижних конечностей. Руки согнуты в локтевых суставах, правая кисть сжата в кулак, голени разведены в стороны, стопы находятся в состоянии подошвенного сгибания за счет повышения тонуса мышц-разгибателей стопы.
При сохранности обоих тонических рефлексов (лабиринтного и шейного) чаще всего имеют место негрубые сгибательные контрактуры в тазобедренных, коленных суставах и разгибательные контрактуры (подошвенное разгибание стоп) в голеностопных суставах. У ребенка формируется «конская стопа». Передвижение возможно только на полусогнутых ногах с упором на пальцы стоп.
В результате того, что лабиринтный установочный рефлекс с головы на шею отсутствует, а тонический лабиринтный рефлекс остается активным, голова постоянно опущена на грудь. Плечо приведено к туловищу, предплечье пронировано (повернуто ладонью вниз), повышен тонус сгибательной мускулатуры рук.

Патологическая установка конечностей, создающаяся под влиянием тонического лабиринтного и симметричного шейного тонического рефлексов
Асимметричный шейный тонический рефлекс наблюдается у здоровых детей до 14 месяцев. Возникает в результате растяжения мышц шеи, связок и суставов шейного отдела позвоночного столба. При этом поворот головы в сторону сопровождается повышением тонуса мышц-разгибателей тех конечностей, в сторону которых обращено лицо, а в противоположных конечностях повышается тонус мышц-сгибателей. Реакция рук отчетливее чем реакция ног.
Сохранение этого рефлекса у ребенка после 14 месяцев указывает на патологию, тормозит развитие активных движений головы и конечностей, мешает фиксации взора на предметах и удержании предметов в руках. У детей, больных ДЦП, возникает характерная поза, носящая название «поза фехтовальщика»: голова повернута влево, повышен тонус мышц-разгибателей левых и тонус мышц-сгибателей правых конечностей.

Патологическая поза ребенка, больного ДЦП, обусловленная асимметричным шейным тоническим рефлексом
Тонические рефлексы у здорового ребенка постепенно угасают. Начиная со 2-го месяца жизни развиваются установочные рефлексы, определяющие возможности статики, локомоции и, в значительной степени, произвольных движений. Особенно сложной и в то же время важной в плане становления установочных рефлексов является деятельность структур головного мозга, ответственных за преодоление силы земного притяжения. Процесс установки тела человека в вертикальное положение контролируется сложной системой различных структур головного и спинного мозга, развитие которых происходит постепенно на протяжении первых двух лет жизни.
На первых этапах становления антигравитационных механизмов основную роль играют структуры вестибулярного аппарата. Последний называется вполне сформированным уже к моменту рождения. Первым, наиболее примитивным, проявлением деятельности вестибулярного аппарата является тонический лабиринтный рефлекс, описанный выше. Более сложный установочный рефлекс, контролируемый деятельностью лабиринтов, — лабиринтный выпрямляющий рефлекс на голову.
Ребенок с появлением этого рефлекса начинает отрывать голову от поверхности, на которой лежит, приподнимать верхнюю половину туловища и, опираясь на предплечья, удерживать ее приподнятой. Данный рефлекс стимулирует развитие цепных симметричных рефлексов, направленных на приспособление туловища к вертикальному положению (шейного симметричного цепного установочного рефлекса и шейного асимметричного цепного установочного рефлекса).
В дальнейшем, по мере развития мозга, появляется еще ряд выпрямляющих реакций и рефлексов.
Так, шейная выпрямляющая реакция, заключающаяся в том, что вслед за пассивным или активным поворотом головы в сторону происходит ротация всего туловища, позволяет ребенку к 4 месяцам поворачиваться со спины на бок.
Выпрямляющий рефлекс, действующий на голову, способствует выпрямлению головы относительно других частей тела.
Выпрямляющий рефлекс, действующий с тела на тело, к 6-8 месяцам видоизменяет примитивную шейную выпрямляющую реакцию, вводя ротацию туловища между плечами и тазом. Во втором полугодии ребенок может поворачивать сначала голову, затем плечевой пояс и, наконец, таз. Ротация в пределах оси тела дает ребенку возможность переворачиваться со спины на живот и наоборот, садиться, вставать на четвереньки и принимать вертикальную позу.
Выпрямляющие реакции, сочетаясь друг с другом, образуют ряд последовательных поз и движений, направленных на приспособление головы и туловища к вертикальному положению. Они достигают согласованности в возрасте 10-15 месяцев и затем совершенствуются.
У детей раннего возраста наблюдаются защитная реакция рук и рефлекс Ландау. Они не относятся к истинным выпрямляющим рефлексам, но на определенных стадиях способствуют развитию двигательных актов.
Защитная разгибательная реакция рук возникает в ответ на внезапное перемещение тела вперед, в сторону, назад и способствует тому, что ребенок в положении сидя может поддерживать свою массу тела руками, вытянутыми вперед.
Рефлекс Ландау комбинируется с выпрямляющими рефлексами и является их частью. Он возникает в возрасте 5-6 месяцев (отдельные элементы раньше) и на втором году жизни начинает угасать. Рефлекс Ландау состоит из двух фаз. Первая обеспечивает разгибание шеи, верхних конечностей и верхней половины туловища у здорового ребенка 5-6 месяцев, положенного на край стола так, чтобы его грудь не касалась поверхности стола. Он может удержаться в таком положении 1-2 минуты.

Рефлекс Ландау у здорового ребенка:
а — первая фаза; б — вторая фаза; в — отрицательный рефлекс Ландау у ребенка, больного ДЦП
У ребенка, больного ДЦП, рефлекс Ландау чаще всего оказывается отрицательным, т.е. разгибания туловища и конечностей при указанном положении тела не происходит, руки и ноги свисают. Такое состояние носит название симптома «свешенного белья». Реакции равновесия — весьма сложные и разнообразные автоматические реакции, необходимые для становления вертикальной позы у ребенка.
Они обеспечивают сохранение равновесия при сидении, стоянии, ходьбе и осуществляются в результате взаимодействия вестибулярного аппарата, базальных ганглиев, ядер субталамической области, мозжечка и коры головного мозга. Это самая высокая форма развития автоматических двигательных реакций, которая развивается и совершенствуется в определенной последовательности до 5-6 лет.
При церебральных параличах в результате поражения мозга в период его интенсивного роста и дифференциации последовательность двигательного развития нарушается. Наряду с замедленным становлением нормальных постуральных механизмов (обеспечивающих вертикальную позу) активизируются тонические рефлексы, которые сосуществуют с патологическим мышечным тонусом (спастичностью, ригидностью, тоническими спазмами или, наоборот, гипотонией) и усугубляют дефекты локомоции.
С января 2017г. в нашем центре ввелась новая процедура на гашение патологических рефлексов, с использованием укладок, лечение положением.
\u042f \u0441\u043e\u0433\u043b\u0430\u0441\u0435\u043d\/\u0441\u043e\u0433\u043b\u0430\u0441\u043d\u0430 \u043d\u0430 \u043e\u0431\u0440\u0430\u0431\u043e\u0442\u043a\u0443 \u0441\u0432\u043e\u0438\u0445 \u043f\u0435\u0440\u0441\u043e\u043d\u0430\u043b\u044c\u043d\u044b\u0445 \u0434\u0430\u043d\u043d\u044b\u0445:
\u0418\u043d\u0444\u043e\u0440\u043c\u0430\u0446\u0438\u044f \u043e \u043f\u0435\u0440\u0441\u043e\u043d\u0430\u043b\u044c\u043d\u044b\u0445 \u0434\u0430\u043d\u043d\u044b\u0445 \u0430\u0432\u0442\u043e\u0440\u043e\u0432 \u043e\u0431\u0440\u0430\u0449\u0435\u043d\u0438\u0439, \u043d\u0430\u043f\u0440\u0430\u0432\u043b\u0435\u043d\u043d\u044b\u0445 \u0432 \u044d\u043b\u0435\u043a\u0442\u0440\u043e\u043d\u043d\u043e\u043c \u0432\u0438\u0434\u0435, \u0445\u0440\u0430\u043d\u0438\u0442\u0441\u044f \u0438 \u043e\u0431\u0440\u0430\u0431\u0430\u0442\u044b\u0432\u0430\u0435\u0442\u0441\u044f \u0441 \u0441\u043e\u0431\u043b\u044e\u0434\u0435\u043d\u0438\u0435\u043c \u0442\u0440\u0435\u0431\u043e\u0432\u0430\u043d\u0438\u0439 \u0440\u043e\u0441\u0441\u0438\u0439\u0441\u043a\u043e\u0433\u043e \u0437\u0430\u043a\u043e\u043d\u043e\u0434\u0430\u0442\u0435\u043b\u044c\u0441\u0442\u0432\u0430 \u043e \u043f\u0435\u0440\u0441\u043e\u043d\u0430\u043b\u044c\u043d\u044b\u0445 \u0434\u0430\u043d\u043d\u044b\u0445: \",\"variants\":[],\"params_value\":\"\",\"validator\":\"\",\"change\":false>>>","nativeStyle":true,"showTitleAfterSend":false,"disabledNativeStyleSelect":false>>'>
Отолитовые рефлексы. Тонические лабиринтные рефлексы
Отолитовые рефлексы. Тонические лабиринтные рефлексы
Тонические лабиринтные рефлексы, относящиеся к отолитовым рефлексам, дают возможность животному привести отдельные части своего тела в гармоническое соответствие друг с другом и поддерживать это соответствие. Лабиринт имеет прямое влияние не только на мышцы конечностей, но также на мышцы шеи и туловища. Кроме первичного влияния лабиринта на шейные мышцы, существует еще вторичное влияние шейных мышц на мышцы конечностей. Это влияние известно под названием шейного рефлекса. Тонический лабиринтный в сочетании с шейным рефлексом обеспечивает соответствие между любым положением головы и остальной части тела.
Кроме тонических лабиринтных рефлексов существует еще лабиринтный восстановительный рефлекс, при помощи которого животное может восстановить прежнее нормальное положение головы и туловища, если оно из него было выведено. Этот рефлекс выпадает, если удалены лабиринты или ствол мозга перерезан каудально (ниже) от красных ядер. Следовательно, для этого рефлекса необходима целость лабиринтов и красных ядер.
При более углубленном изучении отолитовых рефлексов в первую очередь привлекает к себе внимание тонический лабиринтный рефлекс, легче всего наблюдаемый на децеребрированном животном, у которого, следовательно, сняты произвольные движения. Помещая такое животное в разные положения с тем, чтобы его голова перемещалась по отношению к горизонтальной плоскости, можно выявить тонические рефлексы со стороны отолитового аппарата на мышцы конечностей. Так, например, если положить животное на спину, чтобы его темя было направлено вниз, то немедленно возникает напряжение разгибателей всех четырех конечностей и головы; если же положить это животное на живот, чтобы темя было направлено вверх, то тонус этих мышц уменьшается до минимума. Следовательно, наибольшее действие на тонус разгибателей головы и конечностей оказывает свисание утрикулярного отолита, а наименьшее — давление его на подлежащие волоски утрикулярного пятнышка.
После одностороннего удаления лабиринта эти изменения тонуса выпрямителей в зависимости от положения головы в пространстве сохраняются. Следовательно, каждый лабиринт влияет на мышцы конечностей с обеих сторон, чего нельзя сказать про влияние лабиринта на шейные мышцы. Влияние лабиринта на шейные мышцы имеет односторонний характер, так как после удаления одного лабиринта голова животного делает такой поворот (так называемый основной поворот), при котором оперированное ухо оказывается внизу, а здоровое наверху.
У кролика, перенесшего одностороннюю экстирпацию лабиринта, лабиринтный восстановительный рефлекс проявляется с наибольшей силой тогда, когда голова находится в боковом положении с больным ухом наверху (максимальное положение). Рефлекс проявляется минимальным образом, если повернуть голову на 180° и поместить, следовательно, здоровое ухо наверху (минимальное положение). Некоторые авторы пытались связать этот экспериментальный факт с горизонтальным расположением при этих положениях головы главной части саккулярного отолита, причем, как это не трудно представить себе стереометрически, отолитовая мембрана свисает в максимальном положении, а в минимальном—давит на подлежащую саккулярную макулу. На этом основании они приписывали саккулярному отолиту функцию восстановительного рефлекса, имеющего целью восстановить положение головы из латерального положения в нормальное. Однако эта заманчивая гипотеза не подтвердилась, и функция саккулярного отолита до сих пор остается не вполне выясненной.
Отолитовый аппарат реагирует на положение лабиринта в пространстве. Тонические импульсы, возникающие в каждом лабиринте, действуют на шейные мышцы и, следовательно, влияют на положение головы таким образом, чтобы данный лабиринт оказался в наиболее высоком положении по сравнению с положением лабиринта противоположной стороны.
Следовательно, при здоровых лабиринтах каждый из них, если так можно выразиться, имеет тенденцию подняться кверху. Поэтому при ненормальном положении головы более сильные импульсы, идущие из ниже расположенного лабиринта, действуют таким образом на шейную мускулатуру, чтобы перемещение головы подняло этот лабиринт повыше. Как только оба лабиринта оказываются на одном уровне, возникающие в них тонические импульсы на шейные мышцы становятся равными и голова поддерживается в нормальном положении.
Когда положение головы в пространстве меняется, то одновременно меняется взаимоотношение между головой и туловищем и, следовательно, шейные мышцы одной стороны становятся более растянутыми, чем на другой стороне. Это обстоятельство вызывает раздражение заложенных в шейных мышцах нервных окончаний, являющееся причиной возникновения шейного рефлекса, действующего на мышцы конечностей совместно с обычными лабиринтными тоническими рефлексами. При удалении обоих лабиринтов, можно наблюдать шейные рефлексы на конечности в чистом виде. Если поднять голову животного вверх, то напряжение разгибателей передних конечностей увеличивается, а задних уменьшается. Если опустить голову животного вниз, то, наоборот, разгибатели передних конечностей ослабевают, а задних напрягаются.
Поворот головы вокруг продольной оси тела вправо так, что правый глаз оказался бы ниже левого, уменьшает тонус разгибателей в правых конечностях и увеличивает в левых. Поворот же головы к правому или левому плечу вызывает противоположный эффект. Таким образом, поворот головы вокруг продольной оси тела вызывает увеличение тонуса конечностей, к которым направлена челюсть. Это изменение мышечного тонуса держится все время, пока голова находится в повернутом положении. Проявление шейного рефлекса может иметь место и при наличии лабиринтов, лишь бы голова не изменила своего положения по отношению к горизонтальной плоскости.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Установочные рефлексы мозгового ствола
Даже таламическое животное (т.е. животное, у которого удалены большие полушария) способно принимать и поддерживать нормальное положение тела. Это обеспечивается совокупным действием ряда рефлексов мозгового ствола. Примеры этих рефлексов приведены ниже. Лабиринтный установочный рефлекс на голову состоит в том, что при любом положении тела голова животного принимает нормальное положение с параллельно расположенной ротовой щелью благодаря рефлексу с вестибулярного аппарата. Тонические лабиринтные рефлексы на глаза. Компенсаторные движения глаз при поворотах головы и туловища можно наблюдать у всех позвоночных.
Однако у животных с боковым положением глаз (например, у кролика) они выражены более отчетливо. Под действием этих рефлексов глаз максимально перемещается назад, когда голова животного занимает положение мордой вниз.
Максимальный поворот глаза верхним полюсом роговицы вверх происходит при положении головы мордой вертикально вверх. Благодаря этим рефлексам каждому положению головы в пространстве соответствует определенное положение глазного яблока в орбите.
При этом не имеет значения, с какой стороны достигалось положение головы. Всегда устанавливается одно и то же закономерное положение глаз.
У животных с фронтальным расположением глаз (например, у хищных, приматов) при наклоне головы вперед глаза отклоняются вверх. Тонические реакции глаз рефлекторно возникают в результате раздражения отолитового аппарата.
При повороте головы кролика вокруг сагиттальной плоскости также возникают тонические лабиринтные рефлексы на глаза. При этом отклонения обоих глаз при разных положениях головы в пространстве происходят в противоположных направлениях.
Если вращать голову от нормального положения (ротовая щель расположена горизонтально), то глаз, находящийся внизу, отклонится в орбите вверх.
Отклонение достигает максимума при положении головы на боку. Контралатеральный глаз при этом отклонится вниз. При вращении через положение на затылке реакция глаз быстро изменяется на обратную.
Таким образом, существуют два независимых компонента тонического лабиринтного рефлекса на глаза, которые образуют все возможные комбинации при каждом положении головы в пространстве.
Быстрые движения глаз, поскольку они вызываются с полукружных каналов, не участвуют в этих статических рефлексах. Активирование полукружных каналов (вращением головы) вызывает вестибулоокулярный рефлекс.
3. Вестибулоокулярный рефлекс (ВОР) – один из древнейших; он сопровождает позвоночных животных на протяжении всей их эволюции.
Рефлекс проявляется в том, что при вращении головы глаза в орбитах противовращаются. Внешне это проявляется в виде нистагма колебательных движений глазных яблок в орбитах.
Различают медленную компоненту нистагма, при которой глаза вращаются в сторону вращения головы примерно с той же скоростью, и быструю фазу нистагма, при которой глаза в орбитах после достижения своего крайнего положения скачком (саккадой) возвращаются в исходное положение.
Этот рефлекс можно наблюдать как в полной темноте, так и на свету. Биологическая роль этого рефлекса – предотвращать сползание сетчаточного изображения при вращении головы.
Этот афферентный сигнал после обработки выходит на эффекторное звено экстраокулярные мышцы. Главная проблема для такой системы состоит в калибровке: как отрегулировать скорость противовращения глазных яблок, чтобы оно эффективно компенсировало вращение головы.
В процессе калибровки скорости противовращения глазных яблок в орбитах относительно направления вращения головы участвует мозжечок.
Это символически показано одной клеткой коры мозжечка (клеткой Пуркинье), а именно – самой задней частью мозжечка (флоккулонодулярная), которую часто называют вестибулярным мозжечком.
Гипотеза об участии флоккулюса мозжечка в пластичности ВОР была высказана на основе тесных связей вестибулярного мозжечка (flocculus и nodulus) как с вестибулярной, так и со зрительной системой.
В пользу этого предположения говорят также результаты по удалению вестибулярного мозжечка.
Оказалось, после удаления вестибулярного мозжечка (флоккулюса и парафлоккулюса), а также нижней половины VIII дольки и нижних 3-4 долек парамедианной доли усиление ВОР устанавливалось на уровне 1,063 (диапазон 0,95-1,173) и исчезала способность ВОР к пластическим изменениям коэффициента усиления.
Предполагают, что сигналом для изменения усиления ВОР является смещение сетчаточного изображения.
Далее этот сигнал по дополнительной зрительной системе поступает в претектальную область (ядро оптического тракта), затем к нижней оливе и через лазающие волокна из нижней оливы к клеткам Пуркинье коры вестибулярного мозжечка (флоккулюсу).
Сигнал скорости вращения головы передается от верхнего вестибулярного ядра по мшистым волокнам к клеткам-зернам и далее через параллельные волокна поступает также на клетки Пуркинье.
Таким образом, клетки Пуркинье коры вестибулярного мозжечка выполняют оль компараторов (сравнителей), благодаря «работе» которых может модифицироваться коэффициент усиления ВОР.
У человека ВОР участвует в реакции установки взора. Эта реакция возникает при появлении сбоку в поле зрения неожиданного зрительного стимула. В первый момент человек скачкообразно переводит глаза в сторону стимула и «захватывает» зрением этот стимул.
Слово «захватывает» означает, что стимул помещается в область сетчатки с наилучшим разрешением (зрительная ямка, fovea). С небольшим отставанием (это связано с инерцией) в эту же сторону начинает перемещаться голова.
Чтобы изображение не сползло с сетчатки (другими словами, чтобы человек не потерял изображение), включается ВОР, который в данном случае запускается благодаря вращению головы и, следовательно, возбуждению рецепторов полукружных каналов. При этом скорость противовращения глаз в орбитах равна скорости вращения головы. Благодаря этому взор (алгебраическая сумма скоростей вращения глаз и головы) остается неподвижным в пространстве.
ВОР обеспечивает движение глаза в противофазе с движениями головы в диапазоне 0,01-6,5 Гц. На более низкой частоте в стабилизации сетчаточного изображения участвуют также оптомоторный рефлекс и рефлекс прослеживания.
Под «стабилизацией» будем понимать то, что положение глаза относительно зрительной цели не меняется, несмотря на вращение головы. Это достигаете механизмом ВОР: глаза противовращаются в орбитах со скоростью, равной скорости вращения головы.
Для успешной стабилизации сетчаточного изображения при вращениях головы необходимо регулировать коэффициент усиления ВОР.
Нетрудно себе представить ситуации, в которых необходимо изменять усиление ВОР. Например, при росте организма меняются размеры головы и глазных яблок, в силу разных обстоятельств (травмы, процессы старения и пр.) меняется упругость глазодвигательных мышц и многое другое. Коэффициент усиления ВОР регулируется в коре задней части мозжечка.
Оптомоторный рефлекс проявляется в том, что при движении в поле зрения человека структурированного зрительного поля происходит непроизвольное движение глазных яблок в сторону движения стимулов.
Когда движение глазных яблок в орбитах исчерпывает свой диапазон (по 45 угл. градуса от среднего положения), глаза скачкообразно возвращаются в исходное состояние.
Совместная работа трех систем (вестибулоокулярной, оптомоторной и прослеживания) обеспечивает латентный период реакции глазного яблока, не превышающий 80 мс. Другими словами, человек способен следить за объектами, движущимися со скоростью около 300 угл. град. в секунду.
Шейные рефлексы на глаза. Если у животного (например, у кролика) удалить лабиринты, а затем поворачивать его голову, то можно получить все те перемещения глаз, которые описаны выше.
Но в данном случае движения глаз осуществляются исключительно за счет шейных рефлексов. Положение глаз, вызванное этими рефлексами (при изменении положения головы или туловища), в конечном итоге приводит к тому, что изображение на сетчатках обоих глаз остается стабильным.
В норме это достигается только совместным действием лабиринтных и шейных рефлексов. Шейноокулярный рефлекс довольно плохо выражен у интактных взрослых млекопитающих и практически отсутствует в норме у человека.
Шейный рефлекс на конечности. Если поднимать или опускать голову животного в атлантоокципитальном суставе, то вызываемые при этом рефлексы на передние и задние конечности действуют противоположным образом: при опускании головы активируются флексоры передних конечностей и экстензоры задних, при поднимании головы активируются экстензоры передних конечностей и флексоры задних.
Сравнивая тонические рефлексы, можно видеть, что шейные и лабиринтные рефлексы на передние конечности действуют как синергисты, а на задние – как антагонисты.
Рефлекс перераспределения тонуса конечностей при поворотах головы. Если голову децеребрированной кошки (удалены большие полушария), лежащей на животе, поворачивать, то повышается тонус экстензорной мускулатуры в «челюстных» конечностях (конечности той половины туловища, к которым поворачивается нос), уменьшается и увеличивается – в «черепных» (конечности, к которым в этом случае поворачивается череп).
Такое перераспределение тонуса для четвероногих животных оправдано соответствующим перенесением центра тяжести, и для того чтобы выдержать увеличенный вес в соответствующих конечностях, увеличивается их разгибательный (экстензорный) тонус.
Можно получать все описанные шейные тонические рефлексы, если фиксировать позвоночный столб и перемещать только переднюю часть шеи.
У здорового взрослого человека шейные и лабиринтные рефлексы не проявляются, но они отчетливо выступают в патологических состояниях (гидроцефалия, опухоли мозга, тяжелые мозговые травмы и пр.). Например, в этих условиях обнаруживаются шейные тонические рефлексы.
Если поворачивать голову влево, чтобы нос приближался к левому плечу, то левые конечности становятся «челюстными», а правые – «черепными», поэтому повышается разгибательный тонус в обеих конечностях левой половины тела и уменьшается-справа.
Оптический установочный рефлекс играет особенно важную роль в правильной установке головы у высших млекопитающих.
Например, обезьяна с разрушенным лабиринтом может правильно устанавливать голову, но если надеть ей колпак, закрывающий глаза, т.е. лишить зрения, она уже не сможет правильно устанавливать голову в пространстве.
Таким образом, в организме животных взаимодействует ряд установочных рефлексов, благодаря которым обеспечивается нормальное положение головы и тела. Описанные установочные рефлексы осуществляются нервными центрами на уровне не выше среднего мозга.
Лифтная реакция представляет собой рефлекс на прямолинейное ускорение.
Рефлекс состоит в том, что при поднятии площадки, на которой находится животное, тонус экстензоров снижается, и конечности подгибаются; при опускании площадки развивается противоположная реакция: экстензоры активируются и конечности разгибаются. Эта реакция легко вызывается и у человека при подъеме или опускании в скоростном лифте.
Описанные выше установочные рефлексы легко наблюдать на интактной кошке, которая свободно падает из положения на спине. Прежде всего, активируются лабиринтные рефлексы на голову, благодаря которым она поворачивается по направлению к нормальному положению.
К этой реакции присоединяется шейный установочный рефлекс, вследствие чего за головой следует тело, сначала грудной отдел и только затем таз. Происходит своего рода штопорообразное движение животного в пространстве, начинающееся с головы.
При этом передние конечности разгибаются. Дальнейшее прямолинейное смещение головы в пространстве при свободном падении вызывает разгибание задних конечностей. Последнее можно объяснить лабиринтным рефлексом на падение (линейное ускорение).
Благодаря совокупному действию этих рефлексов голова и тело кошки принимают нормальное положение, конечности тонически разгибаются и готовы принять вес тела, когда животное достигнет земли.
Таким образом, положение тела и головы в пространстве и по отношению друг к другу принадлежит к наиболее застрахованным и автоматически выполняемым реакциям ЦНС.
Благодаря взаимной деятельности большого количества разнородных рефлексов осуществляется единая конечная цель – правильная установка головы и тела в пространстве.
Статокинетические и статические реакции взаимно дополняют друг друга: благодаря кинетическим реакциям выполняется движение, приводящее отдельные части тела в такое положение, в котором они потом удерживаются статическими рефлексами.
Взаимодействие вестибулярного и шейного рефлексов
Вестибулярные (лабиринтные) и шейные позотонические рефлексы описал Магнус (Haltungsreflexe). Описал - мягко сказано, работа для 20х годов совершенно грандиозная.
Проблемы есть не столько с его описанием, сколько с последующими интерпретациями. Во-первых, принято считать, что Магнус описал шейный рефлекс ассиметричным, а лабиринтный - симметричным относительно конечностей. Ниже можно увидеть, что они оба одинаково ассиметричны, но противоположны.
Во-вторых, в учебниках часто можно увидеть примерно такую мысль, c пиететом приписываемую Магнусу(*)
Это утверждение довольно странно, если не сказать безграмотно. Такая "прямая" работа вестибулярного рефлекса могла бы быть полезна мифическому животному - колобку, но у человека и котов вестибулярный аппарат расположен в голове, а она на гибкой шее. Однако именно такая концепция, вслед за Магнусом, устоялась весь XX век - что лабиринтные и шейные позотонические рефлексы "распределяют" тонус между мышечными группами.
Содержание
Шейное взаимодействие
Координатная трансформация
Вместо концепции "распределения тонуса" на основе лабиринтных ощущений, и отдельного "распределения" на основе шейных, на эту проблему можно посмотреть иначе.
Вестбулярный поток ощущений был бы очень полезен для позного контроля, но он отражает движения головы, а не центра масс тела. Для использования в позных задачах в этом потоке надо учесть движение шеи, как минимум. Фактически (шея более подвижна, чем тело), необходимо вычесть из движения головы (вестибуляр) движения шеи (проприоцепция шеи).
Это вычитание является по сути преобразованием координат - из системы, связанной с головой, в систему туловища.
Можно, конечно, сказать, что рефлекс не обязан быть таким умным, что он подавляется и направляется высшими структурами и задача с таким сложным названием должна решаться где-то там. Но оказывается, такое преобразование координат прекрасно выполняют именно рефлексы, описанные Магнусом, взаимодействуя друг с другом на уровне ствола (возможно мозжечок участвует). Речь идет о лабиринтном рефлексе положения и АШТР.
Это успешно, и, похоже, независимо, продемонстрировали шотландец Tristan DM Roberts, воспроизведя на уровне технологий 1970х годов работы Магнуса, и немец Kornhuber. Оба указывают, что Магнус некорректно описал лабиринтные рефлексы положения. Они ровно настолько же ассиметричны, как и АШТР, но противоположны по знаку. Фактически можно говорить об ассиметричном лабиринтно тоническом рефлекса - АЛТР. А сам принцип преобразования координат на основе взаимодействия шейных и лабиринтных рефлексов впервые описали von Holst и Mittelstaedt в своем Das Reafferenzprinzip в 1950 (как ни странно, ни тот ни другой на них не ссылаются).
Более того, есть почти что прямые наблюдения именно такой работы нейронов вестибулярных ядер, и спинного мозга. И есть практические наблюдения (неопубликованные) что АЛТР наблюдаем у тяжелых детей в явном виде.
Ниже я привожу перевод выдержек из статьи TDM Roberts в Nature.
Ассиметричный (!) Лабиринтный рефлекс и Ассиметричный Шейный Тонический Рефлекс
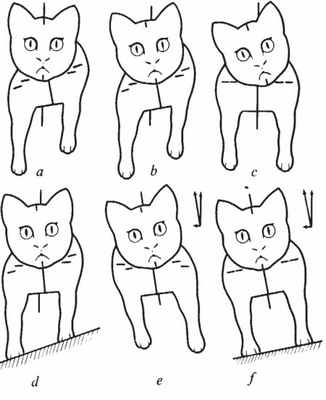
a, Шейные рефлексы отдельно. Тело наклонено, голова прямо, лапы со стороны подбородка разгибаются. b. Лабиринтные рефлексы отдельно. Голова и тело отклонены, шея прямая - нижние лапы разгибаются. c. Отклонение головы отдельно. Лапы симметричны - не разгибаются и не сгибаются, не реагируют на поворот вообще (ВМ). d. Неровная опора. тело отклонено, лапы в компенсирующей позе, голова свободна. e. Постоянное боковое ускорение. Лапы ассиметрично соответствуют отклонению тела относительно вектора опоры. f. Постоянное боковое ускорение. Лапы симметричны на адекватно наклоненной опоре рисунок из статьи TDM Roberts, подробнее см. статью
Успех поддержания вертикальной позы обычно приписывают рефлексам, инициируемым рецепторами лабиринтов внутреннего уха. Традиционные описания работ этих рефлексов, однако, не объясняют наблюдаемую стабильность. Согласно Магнусу, изменение положения головы изменяет тонус разгибателей всех четырех конечностей животного симметричным образом. В противовес этому, тонические шейные рефлексы описаны как ассиметричные в своей реакции на конечности, и лапы, на стороне куда поворачивают челюсть выпрямляются, а с другой стороны - сгибаются.
Соответственно, Робертс занялся повторным исследованием рефлексов, вызываемых отклонением головы, используя котов, децеребрированных несколько выше межколликулярного уровня во избежания чрезмерной ригидности, используя аппарат, в котором независимо можно поддерживать и поворачивать тело, шею и голову кота (описание см. Lindsay, TDM Roberts & Rosenberg 1976), включая устрашающую возможность поворачивать шейные позвонки относительно неподвижных туловища и головы.
Лабиринтные рефлексы в ответ на отклонение головы обнаружены неизменно ассиметричными и пригодными для функции стабилизации, в отличии от симметричной схемы Магнуса.
Их можно описать принципом "нижние лапы разгибаются, верхние лапы сгибаются"
Когда поворачивается шея, "лапы со стороны подбородка разгибаются", в полном соответствии со схемой Магнуса и Клейна.
Однако ответ на шейные рефлексы противоположен ответам на лабиринтные рефлексы при аналогичном повороте шеи. Действуя одновременно, эти рефлексы суммируются, и взаимодействие этих двух наборов рефлексов приводит к стабилизации туловища, независимой от поворота головы.
Что из этого взаимодействия получается

Далее Робертс начинает расписывать алгебраические уравнения, но принцип суммирования этих рефлексов (точнее вычитания - они же противоположны, антагонистичны по действию) можно описать проще (я для этого воспользуюсь картинкой из работы Kornhuber, они, как видно, близнецы-братья):
- При стабильном положении тела поворот головы вызывает лабиринтную реакцию (АЛТР), которая полностью компенсируется АШТР - суммарный эффект на конечности нулевой.
- Однако если наклоняется все тело, вместе с головой - лабиринтная реакция (АЛТР) будет больше, чем АШТР, и суммарный рефлекторный ответ будет компенсировать отклонение.
- Если же тело "выскальзывает" из под стабильной головы, то АШТР будет больше, чем лабиринтная реакция (АЛТР), и суммарный рефлекторный ответ снова будет компенсировать отклонение
Суммарный эффект - получается такой, что
- голову можно вращать как угодно (и потребно для задач зрения, например)
- суммарная реакция на конечности получается такой, как если бы вестибулярный "сенсор" находился в туловище.
Задача преобразования координат успешно решена!
Кто ее решает? Есть основания полагать, что процесс "вычитания" осуществляется определенной подгруппой нейронов в вестибулярных ядрах. Однако аналогичные "вычитающие" нейроны найдены и в interpositus nucleus мозжечка (теми же авторами, см. Luan&Gdowski), и в черве мозжечка (см. Manzoni, Pompeano, Andre). В силу наличия прямых связей между всеми этими зонами, сложно сказать, кто из них первичен, несмотря на то, что Kornhuber утверждает, что "вычитание" от мозжечка не зависит. Более аккуратные эксперименты итальянцев в 1998г показывают, что зависит.
Эффект и "голого рефлекса" и "рефлекса с трансформацией координат", похоже, можно наблюдать как Short latency и Medium latency VSR у человека. См. там же о роли мозжечка в этих трансформациях.
Отмечу так же, (см. Manzoni, Pompeano, Andre), что для прямостоящего человека важно не только положение шеи, но и взаимная ориентация каждого из сегментов оси. Общая картина гораздо сложнее чем "АЛТР минус АШТР", но принцип работы, видимо, именно такой. См. также ниже про поясничные рефлексы.
Corollary discharge/ принцип реафферентации
Первое упоминание описанного вычитания не случайно появляется именно в Das Reafferenzprinzip. При движении головы (неважно, активном или пассивном) вестибулярный ответ является известным, предсказуемым сенсорным последствием, или Reafference которое следует вычесть из общего сенсорного потока- тогда останется только Exafference, которое будет описывать движение тела вместе с головой и шеей.
То есть неважно, как это называть - преобразованием координат или эффектом corollary discharge, это описывает одно и то же явление в данном случае.
Почему АШТР может проявляться у младенцев?
Описанные выше эксперименты выполняются на децеребрированных котах (и других животных), что делает рефлексы видимыми. Проявление АШТР же вообще считается признаком патологии, и во всяком случае ожидается, что он должны исчезнуть с возрастом. Однако даже у взрослой нормы рефлекторные цепи вполне присутствуют и активны, хотя для их выявления требуется более тонкие измерения (измерять ЭМГ или Проприоцептивные рефлексы), или же они вылезают наружу в виде движения/позы в ситуациях большой нагрузки, например в спорте.
Отсутствие видимых рефлексов в норме в данном случае почти наверняка означает, что лабиринтные и шейные рефлексы настолько хорошо синхронизированы между собой, что внешне не проявляются, компенсируя друг друга. Координатная трансформация, которую они осуществляют, однако, представляется слишком полезной))
Можно предположить, что проявление АШТР является следствием незрелости или отклонения в развитии нервной системы, когда уже созревшая нервная цепь рефлекса не получает еще необходимой регулировки от мозжечка, или же это просто этап в этой самой регулировке, когда несогласованное действие АШТР и лабиринтных рефлексов создает ненужный "моторный шум". Этот шум, вероятно, должен быть обнаружен в Inferior Olive и привести к мозжечковой регулировке силы рефлексов до их полного согласования. Или же, отсутствие шума и проблем с ним должно вести к успеху решения первых моторных задач и появлению сигнала подкрепления со стороны базальных ядер. Так или иначе, можно предположить, что наблюдение АШТР у младенцев или пациентов с ДЦП является проявлением задержки этого этапа.
В норме АШТР и лабиринтные рефлексы являются частью единой системы. Их нет смысла разделять, когда мы говорим о нормальной функции. А если ребенок проявляет ассиметричный шейно-тонический "рефлекс" - это означает, что эта система дает сбой (слабость лабиринтного рефлекса, или слабость регулирующих механизмов).
У совсем тяжелых детей ЛМ Зельдин иногда наблюдает и реакцию, противоположную по построению АШТР — иными словами Ассиметричный Лабиринтный Тонический Рефлекс - АЛТР.
Известно также, что симптомы анестезии или повреждения задних корешков шейных отделов С1-С3, нарушающих проприоцепцию шеи, приводит к нистагму, атаксии и ощущениям падения или наклона - что чрезвычайно напоминает симптомы лабирентэктомии Wilson&Peterson
Шейное Головокружение
Существует - весьма спорный - диагноз, "шейное головокружение" - cervical vertigo, спорный потому, что это диагноз исключения, и список исключений там длинный. Подробный хороший обзор на русском языке можно найти в посте laesus-de-liro , где приводится удачное определение этого состояния - «неспецифическое ощущение нарушения ориентации в пространстве и равновесия, обусловленное патологической афферентной импульсацией из области шеи».
Фактически, это нарушение того самого взаимодействия, которое обсуждается в настоящей статье.
Установка тела

По всеобщему мировому признанию труд известного голландского физиолога Р. Магнуса «Установка тела» оценивается как классическое произведение, в котором изложена одна из существенных систем рефлекторной деятельности животного организма. Имя знаменитого ученого ставится рядом с тремя другими основоположниками современной разработки законов деятельности центральной нервной системы - И.П. Павлова, Н.Е. Введенского, Ч.С. Шеррингтона. Рудольф Магнус принадлежит к той плеяде великих учителей физиологии XIX и начала XX в., жизнь которых неотделима от создания целых областей науки.
Лучшая подборка с книгой
Вещий КритиК (AleksSar) Вещий КритиК создал в подборках

Список 100 важных книг по версии Александра Невзорова








8 декабря 2020 г.
Год издания: 1962
Подборки
Вещий КритиК (AleksSar) Вещий КритиК создал в подборках

Список 100 важных книг по версии Александра Невзорова
8 декабря 2020 г.
Похожие книги
Вы можете посоветовать похожие книги по сюжету, жанру, стилю или настроению. Предложенные вами книги другие пользователи увидят здесь, в блоке «Похожие книги». Посоветовать книгу
Читайте также: