Рейсснерова мембрана улитки. Спиральная связка и сосудистая полоска улитки
Добавил пользователь Alex Обновлено: 21.01.2026
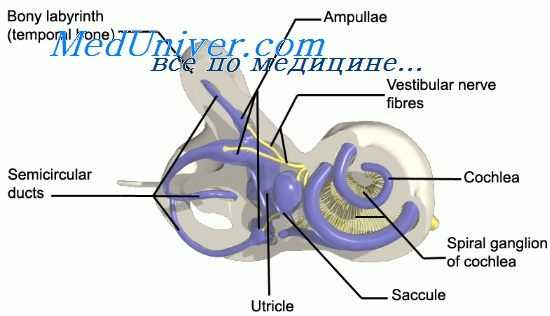
, maculae [[staticae]]. Содержат рецепторные клетки, воспринимающие информацию о положении головы в пространстве. Рис. А, Рис. Б.
Пятно эллиптического мешочка
, macula utriculi. Горизонтально ориентированная область дну эллиптического мешочка размером 2,3 - 3 мм. Рис. Б.
Пятно сферического мешочка
, macula sacculi. Ориентированная в вертикальной плоскости, дугообразно изогнутая рецепторная область шириной около 1,5 мм на медиальной стенке сферического мешочка. Рис. Б.
Статоконии
, statoconia. Кальцийсодержащие частицы размером до 15 мкм, погруженные вместе с волокнами рецепторных клеток в желевидное вещество мембраны статоконий. Рис. А.
Мембрана статоконий
, membrana statoconiorum. Расположена в области пятен эллиптического и сферического мешочков и состоит из желеобразного основного вещества, в которое погружены статоконии и отростки рецепторных клеток, имеющие вид щетинок. Рис. А.
Нейроэпителий
, neuroepithelium]]. Однослойный, призматический эпителий, покрывающий пятна эллиптического и сферического мешочков, состоящий из опорных и рецепторных клеток. Рецепторные клетки имеют волоски длинной 20 - 25 мкм, проникающие в мембрану статоконий. Рис. А.
Улитковый лабиринт
Перилимфатическое пространство
, spatium perilymphaticum. Разделяется на две лестницы. Пронизано соединительнотканными перегородками и содержит перилимфу. Рис. А, Рис. Б.
Лестница преддверия
, scala vestibuli. Расположена над костной спиральной пластинкой и улитковым протоком. Продолжается до верхушки улитки (геликотрема). Рис. В.
Барабанная лестница
, scala tympani. Часть перилимфатического пространства под костной спиральной и базилярной пластинками. Рис. В.
Водопровод улитки
, aquaeductus cochleae [[ductus perilymphaticus]]Соединяет перилимфатическое пространство с подпаутинным. Рис. Б.
Наружное отверстие водопровода улитки
, apertura externa aquaeductus cochleae. Открывается вблизи канальца, через который проходит барабанный нерв. См. стр. 14.22
Улитковый проток
, ductus cochlearis. Трубка, имеющая треугольную форму на поперечном срезе, которая закручивается на 2 1/2 оборота вокруг костной оси (стержня) и слепо заканчивается у верхушки улитки. Покрыта изнутри рецепторным эпителием. Рис. Б, Рис. В, Рис. Г, Рис. Д.
Слепое выпячивание купола
Слепое преддверное выпячивание
Барабанная стенка улиткового протока (спиральная мембрана)
, paries tympanicus ductus cochlearis (membrana spiralis). Нижняя стенка улиткового протока над барабанной лестницей. Рис. Д.
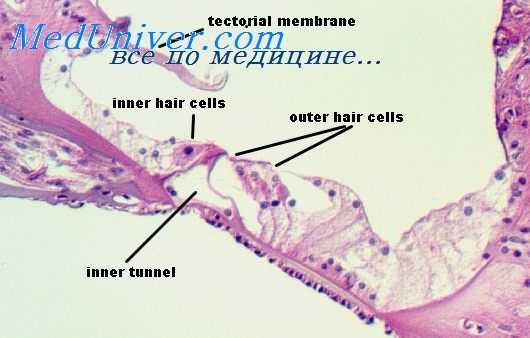
Спиральный [[кортиев]] орган
, organum spirale [[Corti]]. Расположен на базилярной пластинке. Трансформирует механические колебания эндолимфы и базилярной мембраны в нервные импульсы. Рис. Г.
Базилярная пластинка
, lamina basilaris. Состоит из соединительной ткани и формирует нижнюю стенку улиткового протока. Натянута между барабанной губой костной спиральной пластинки и спиральным гребнем. Рис. Д.
Спиральный гребень (спиральная связка)
, crista spiralis (lig. spirale). Полоска утолщенной надкостницы спирального канала улитки, которая имеет треугольную форму на поперечном срезе и продолжается в базилярную пластинку. Рис. Д.
Отверстия нервов
, foramina nervosa. Находятся в базилярной пластинке и содержат волокна улиткового нерва, направляющиеся от волосковых клеток к спиральному узлу. Рис. Г.
Край костной спиральной пластинки
, limbus laminae spiralis osseae. Утолщение эндоста верхнего слоя костной спиральной пластинки, на которой снаружи имеется внутренняя спиральная борозда. Рис. Д.
Губа преддверного края
, labium limbi vestibulare. Верхний, более короткий выступ костной спиральной пластинки. Место прикрепления покровной мембраны. Рис. Д.
Губа барабанного края
, labium limbi tympanicum. Нижний, более длинный выступ края костной спиральной пластинки, который продолжается в базилярную мембрану. Рис. Г, Рис. Д.
Покровная мембрана
, membrana tectoria. Расположена над спиральным органом в виде узкой полоски, которая прикрепляется к губе преддверного края. Ее свободный край заканчивается на уровне ряда наружных волосковых клеток. Рис. Г, Рис. Д.
Слуховые зубчики
, dentes acustici. Ряды клеток, выступающие наподобие гребней на поверхности губы преддверного края. Место прикрепления покровной мембраны. Рис. Г.
Внутренняя спиральная борозда
Наружная спиральная борозда
, sulcus spiralis externus. Находится на барабанной стенке улиткового хода между спиральным гребнем и кортиевым органом. Рис. Д.
Рейсснерова мембрана улитки. Спиральная связка и сосудистая полоска улитки
Кохлеарный или слуховой нерв. Афферентные волокна внутреннего уха
N. cochlearis толще вестибулярного нерва [он состоит, по Расмуссену (Rasmussen), из 31 000 волокон]. Часть пучков кохлеарного нерва проходит через tractus spiralis foraminosus непосредственно к основному завитку улитки, вторая часть вступает в modiolus. На своем пути из центрального канала modiolus к костной спиральной пластинке, через которую последние пучки волокон доходят до основной мембраны, они прерываются в клетках мощного узла в саnalis spiralis modioli s. Rosenthali.
Этот узел—gangl. spirale—является нервной округлой пластинкой, идущей спирально от основного завитка до вершины улитки. Клетки спирального узла биполярны. От них идет периферический дендрит к кортиеву органу и центральный неврит к центрально расположенным слуховым ядрам. Проходя через основную мембрану, нервные волокна теряют миелиновую оболочку и уже как безмякотные волокна (осевые цилиндры) подходят к кортиеву органу. Здесь указанные волокна разветвляются в спиральном и радиальном направлении, образуя пучки нервных волокон, и в виде тончайших волоконец подходят к чувствительным (волосатым) наружным и внутренним клеткам, образуя вокруг их основания нежные бокаловидные сплетения.
По данным Анджеевского, нервные волокна оплетают не только волосатые клетки, но и дейтерсовские клетки, проникая в их цитоплазму. Это тесная связь дейтерсовских клеток с нервной тканью говорит за то, что эти клетки, по-видимому, играют не только поддерживающую роль. Следует отметить, что в нервных рецепторах преддверия (макулы мешочков) и полукружных каналов (ампулярные гребни) концевые нервные волокна разветвляются и снабжают не одну, а несколько чувствительных клеток.
В то же время к каждой чувствительной клетке подходит не одно, а несколько нервных волокон. Гибель любого из этих волокон не влечет за собой смерть клетки, так как она иннервируется еще и другими волокнами. В кортиевом же органе к каждой чувствительной клетке подходит только одно концевое нервное волокно, не дающее ответвлений к соседним клеткам. Дегенерация нервного волокна сопровождается гибелью соответствующей клетки.
Исследования последних лет показали, что кортиев орган получает двойную иннервацию: 1) система волокон с превалирующим радиальным направлением—афферентная, или чувствительная; 2) система волокон с доминирующим спиральным направлением—эфферентная, центрифугальная.
Афферентные волокна давно известны. Они образуют ствол кохлеарного нерва, их клетки находятся в спиральном ганглии. Дендриты этих клеток, идущие в радиальном направлении по отношению к оси modiolus, направляются к кортиеву органу, как описано выше, и образуют plexus spiralis internus и plexus spirales externi. Внутреннее спиральное сплетение дает нервное окончание к внутренним волосатым клеткам. Часть дендритов проходит тоннель кортиева органа и затем, принимая спиральное направление, перпендикулярное к их первичному направлению, идет под наружными волосатыми клетками, образуя наружные спиральные сплетения, предназначенные для трех рядов наружных волосатых клеток. Небольшая часть дендритов проходит отрезок пути не радиально, а спирально, но затем снова принимает радиальное направление и вступает в кортиев орган ближе к геликотреме, чем к чувствительным клеткам.
- Вернуться в оглавление раздела "Анатомия человека."
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Основная мембрана состоит из трех слоев. Средний (волокнистый) построен из радиальных волокон, напоминающих коллагеновые. Этих волокон (слуховых струн) насчитывается около 24 000. Они наиболее длинные и толстые у верхушки улитки и наиболее короткие и тонкие у ее основания.
Верхний, вестибулярный, слой, на котором располагается кортиев орган, очень тонкий и состоит из светлого гомогенного вещества. Нижний, обкладочный, тимпанальный слой богат клетками. Благодаря технике и методике прижизненного наблюдения через проделанное окно в улитке вибраций механических структур внутреннего уха во время акустического раздражения (Бекеши) установлено, что основная мембрана имеет варьирующий импеданс по всей ее длине.
Импеданс—сопротивление вибрационному движению структуры, подвергающейся воздействию приводящей ее в движение силы,—результирует из трех основных свойств: инерции, обязанпой массе, крепости, обязанной эластичности, сопротивления, обязанного трению; эффект этих трех свойств математически комбинируется в единицу, известную как механический импеданс структуры по отношению к вибрации.
Структуры улиткового хода оказывают относительно постоянный импеданс по всей их длине по отношению к силам, приводящим их в движение, в перилимфе, за исключением основной мембраны. Импеданс последней самый высокий у основания и низкий у верхушки. Постоянно верьирующий механический импеданс мембраны результирует с постоянно варьирующей скоростью волны деформации, вызванной движением пластинки стремени. Бекеши установил, что при очень низких частотах движение основной мембраны совпадает в фазе с движением пластинки стремени, а при повышении частоты отставание в фазе между движениями стремени и мембраны увеличивается.
Таким образом, звуковое раздражение распространяется по мембране, как бегущая волна, становящаяся короче при повышении частоты.
Рейсснерова мембрана начинается на вестибулярной поверхности костной спиральной пластинки и прикрепляется к верхней части спиральной связки. Она представляет тонкую соединительнотканную пластинку с небольшим количеством эластических волокон, лишенную сосудов. На вестибулярной стороне она покрыта слоем эндотелиальных клеток, а на стороне улиткового хода—однородным плоским эпителием.
Эндост, выстилающий изнутри перилимфатическое пространство улитки, переходя на наружную сторону завитка улитки, значительно утолщается в соединительнотканный слой, снабженный многочисленными сосудами и имеющий на поперечном разрезе форму серпа—lig. spirale. Спиральная связка на всем протяжении улитки является наружной границей обеих лестниц и улиткового хода (ductus cochlearis). В центре внутренней поверхности спиральной связки имеется выступ, переходящий непосредственно в основную мембрану.
Спиральная связка состоит из нежных коллагеновых волокон и небольшого количества звездчатых соединительнотканных клеток. Со стороны улиткового хода она выстлана так называемой stria vascularis (сосудистая полоска). Сосудистая полоска образована слоем высокого эпителия, под которым находится еще несколько слоев эпителиальных клеток, самый глубокий из которых построен из более низких клеток. Между слоями эпителия проходит большое количество капилляров и прекапилляров.
Высокий эпителий переходит с одной стороны в плоский эпителий райсснеровой мембраны, с другой— в клетки Клаудиуса. В глубоких слоях эпителия сосудистой полоски проходят многочисленные капилляры в два слоя, находящиеся в тесном контакте с эпителиальными клетками, без прослойки соединительной ткани. Таким образом, здесь имеется эпителий, содержащий сосуды, истинный сосудистый эпителий. Большинство клеточных элементов сосудистой полоски относится к хромофильной группе клеток. Гистохимическое исследование показало, что пигмент сосудистой полоски образуется трансформацией плазмы и ядра. Между наличием пигмента и внутриклеточным аппаратом Гольджи имеется определенная связь. В базальной части клеток и всегда внутри от аппарата Гольджи продуцируется эндолимфа [Фиандт и Саксен (Н. Fieandt и A. Saxen)].
По мнению последних авторов, повышение эндолимфатического давления во внутреннем ухе улавливается поддерживающими волокнами хромофильных клеток и кровеносными сосудами. Механическое сдавление кровеносных сосудов сопровождается уменьшением секреции эндолимфы, что ведет к восстановлению нормального давления в улитковом ходе. При понижении давления имеет место противоположный механизм: расширение сосудов с повышением секреции и восстановлением эндолабиринтного давления.
Физиология сосудистой полоски внутреннего уха
Сосудистая полоска играет ключевую роль в гомеостазе улитки, генерируя внутриулитковый потенциал и сохраняя специфический ионный состав эндолимфы. Сосудистая полоска является высоко васкуляризированной многослойной тканью, входящей в состав латеральной стенки улиткового протока. Она включает три различных типа клеток (краевые, промежуточные и базальные), каждый из которых необходим для ее функционирования.
Щелевые соединения обеспечивают ионный барьер, который отделяет сосудистую полоску, одни находятся на уровне маргинальных клеток, а другие на уровне базальных клеток. Внеклеточное пространство между этими двумя барьерами называется интрастриарным (внутриполосковым) пространством. Как показано на рисунке ниже, маргинальные клетки отделяют улитковый проток, наполненный эндолимфой, от интрастриарного пространства, которое наполнено интрастриарной жидкостью, в то время как базальные клетки отделяют интрастриарное пространство от перелимфы, окружающей фиброциты спиральной связки.
Промежуточные клетки, так же как и кровеносные сосуды входят в состав интрастриарного пространства. Щелевые межклеточные промежутки соединяют базальные клетки с промежуточными клетками и с фиброцитами спиральной связки, обеспечивая электрическую передачу, а также обмен ионов и небольших молекул. Система регуляции гомеостаза кохлеарной жидкости также включает эндолимфатический мешок, который отвечает за изменения объема эндолимфы и, возможно, изменяет гомеостаз при нарушении его функционирования.
Нарушения гомеостаза улитковой жидкости, вызванные изменениями внутриулиткового потенциала, ионного состава или механизмов его регуляции, ведут к различным формам тугоухости у людей и животных.
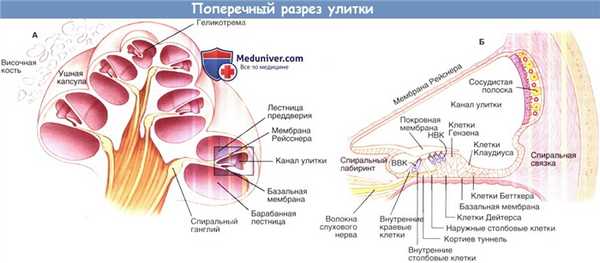
Поперечный разрез улитки. А, схематичное изображение улитки в разрезе.
Выделенная область, показанная на рисунке Б, изображает улитковый проток и окружающие структуры, такие как кортиев орган и сосудистую полоску в разрезе.
ВВК — внутренние волосковые клетки; НВК — наружные волосковые клетки.
а) Внутриулитковый потенциал и гомеостаз калия. Эффективная механоэлектрическая трансдукция (передача) волосковых клеток обеспечивается благодаря большой движущей силе катионов, входящих в цитоплазму клеток из улиткового протока (медиальной лестницы). Внутриулитковый потенциал эндолимфы, составляющий приблизительно +85мВт, и химический градиент К + являются основными компонентами движущей силы, которая достигает 130 мВт, так как в покое потенциал волосковой клетки составляет -45 мВт. При снижении внутриулиткового потенциала на 1 мВт пороги слышимости увеличиваются примерно на 1 дБ.
К + , являющийся основным катионом эндолимфы, несет в себе большую часть электрического заряда, который генерирует внутриулитковый потенциал. Поэтому важно понимать, каким образом К + движется через улитку. К + может входить в волосковые клетки через каналы механоэлектрической передачи и высвобождаться через базолатеральные мембраны волосковых клеток в перелимфатическое внеклеточное пространство. Предполагается, что К + может входить в поддерживающие клетки и двигаться по направлению к спиральной связке через широкие межклеточные пространства.
Кроме того, К + может диффундировать внеклеточно через перелимфатическое пространство.
Фиброциты I и II типов спиральной связки захватывают К + и обеспечивают его внутриклеточное перемещение в базальные и промежуточные клетки сосудистой полоски. К + выпускается промежуточными клетками через KCNJ10 каналы в интрастриарное пространство, из которого он активно засасывается и ко-транспортируется в маргинальные клетки. Маргинальные клетки выделяют К + в улитковый проток. В целом циркуляция К + на самом деле не является истинной циркуляцией, так как перелимфа и интрастриарное пространство не образуют закрытую петлю, поскольку эти отсеки связаны с другими внеклеточными пространствами и с кровотоком.
Сбои в работе некоторых К + каналов ведут к нарушению улиткового К + гомеостаза, приводя к ухудшению слуха. У мышей потеря гена KCNE1, который кодирует субъединицу К + канала, экспрессируемую маргинальными клетками, формирует очень схожий с кардиоаудиторным синдромом фенотип, характеризующийся снижением слуха и аритмией. Это наблюдение привело к выявлению двух генов человека, KNCE1 и KCNQ1, мутации которых вызывают кардиоаудиторныйсиндром.Вполне возможно, что KCNQ1 и KNCE1 формируют каналы, которые обеспечивают секрецию калия из маргинальных клеток в улитковый проток.
Другой член семейства KCNQ калиевых каналов, KCNQ4, скорее всего, участвует и базолатеральной секреции К + волосковыми клетками. Мутация у человека гена KCNQ4 приводит к несиндромной глухоте. Другие известные генетические нарушения включают транспортирующие ионы белки с локализацией на базолатеральной мембране краевых клеток. Вероятно, наиболее известные гены, вовлеченные в калиевый гомеостаз улитки — гены, кодирующие белки-коннексины. Коннексины формируют субъединицы щелевых контактов каналов, которые лежат в основе циркуляции К + , описанной для поддерживающих клеток кортиева органа, спиральной связки и сосудистой полоски. Мутация генов, кодирующих коннексины человека 26, 30, 31 и 43, отвечает за большинство несиндромных наследственных потерь слуха.
Сосудистая полоска и циркуляция ионов K + .
Схема тока ионов К + от улиткового протока через волосковые клетки в перелимфатическое пространство,
а также через межклеточные пространства к спиральной связке.
К + от спиральной связки транспортируется посредством сосудистой полоски в улитковый проток.
Не показаны другие пути тока К + из улиткового протока, а именно через наружные клетки борозды и мембрану Рейсснера.
Прямоугольной скобкой выделено пространство, более детально показанное на рисунке ниже. Ток К + через кортиев орган и сосудистую полоску.
А. К + входит в волосковые клетки посредством каналов механической передачи.
В базально-латеральных отделах калий выводится в перелимфатическое пространство посредством K + -каналов, таких как канал KCNQ4.
К + может перемещаться по направлению к спиральной связке через перелимфатическое пространство и внутриклеточно, через сеть межклеточных пространств.
Фиброциты II типа спиральной связки выводят К + и обеспечивают его передвижение к сосудистой полоске через сеть пространств в соединительной ткани.
Б. К + входит в базальные и промежуточные клетки через пространства между фиброцитами I и II типов.
К + -канал KCNJ10 является важным звеном в высвобождении калия в пространство внутри сосудистой полоски.
Ген, кодирующий KCNJ10, необходим для правильной генерации внутриулиткового потенциала.
К + эффективно выводится из пространства внутри сосудистой полоски краевыми клетками, которые активно выводят К + посредством NKCC1 (Na + /К + /2Cl) ко-транспортеров и Na + /К + -АТФаз.
В итоге, маргинальные клетки секретируют К + в улитковый проток посредством KCNQ1/KCNE1 К + -каналов поддерживая высокую концентрацию К + в эндолимфе, необходимую для механоэлектрической передачи.
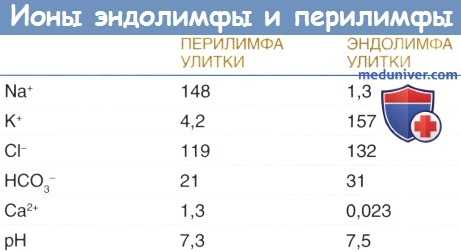
б) Гомеостаз жидкостей улитки. Перелимфа, эндолимфа и жидкость, располагающаяся внутри сосудистой полоски (интрастриарная жидкость), являются тремя разными жидкостями улитки, которые можно рассматривать как систему метаболической поддержки. Для генерации и поддержания внутриулиткового потенциала необходим сбалансированный ионный состав этих трех жидкостей. Перелимфа и интрастриарная жидкость характеризуются высоким содержанием Na + и низким содержанием К + , как и другие внеклеточные жидкости.
Эндолимфа не только имеет высокую концентрацию К + и низкую концентрацию Na + , но также имеет необычайно низкую концентрацию ионов Са 2+ по сравнению с другими внеклеточными жидкостями. Гомеостаз ионов Са 2+ в улитке контролируется ионными каналами и переносчиками, находящихся в плазматических мембранах клеток, как было описано выше для калия.
В сосудистой полоске поступление ионов Na + сопряжено с выведением ионов К + из интрастриарного пространства в маргинальные клетки. Переносчики NKCC1 используют высокие градиенты натрия для переноса ионов Na + , К + и 2Cl - в маргинальные клетки. Na + /K + -АТФ-аза контролирует работу этого градиента посредством нагнетания Na + в интрастриарное пространство в обмен на К + .
Наконец, К + покидает маргинальные клетки и выводится в эндолимфатическое пространство под воздействием высокого положительного потенциала покоя маргинальных клеток. Этот сложный процесс поддерживает высокую концентрацию ионов Na + и низкую концентрацию К + в интрастриарной жидкости, что облегчает пополнение калием интрастриарного пространства. Ионы хлора переносятся обратно к интрастиарному пространству при помощи С1С-К/барттиновых каналов. Ингибирование NKCC1 и Na + /К + -АТФ-азы петлевым диуретиком фуросемидом и уабаином приводит к уменьшению внутриулиткового потенциала.
Мутация гена, кодирующего барттин, или мутация обеих субъединиц базолатеральных Cl - каналов ClC-Ка и ClC-Kb приводит к синдрому Барттера 4 типа, который характеризуется глухотой и потерей почкой солей. Натрий реабсорбируется из эндолимфы наружной бороздой и клетками Рейснеровой мембраны, которые играют роль в сохранении низкой концентрации Na + в улитковом протоке.
Регулирование концентрации ионов Са 2+ в эндолимфе крайне важно. Изучение волосковых клеток показало, что при низких концентрациях Са 2+ происходит разрыв концевых связей, а при высоких концентрациях Са 2+ блокируются каналы механоэлектрической передачи. Кроме того, ионы Са 2+ частично обеспечивают работу потоковой передачи и играют важную роль в адаптации и, возможно, в улитковой амплификации. Са 2+ -проницаемые каналы, кальциевая АТФ-аза, так же как и Na + /Ca 2+ -обмен обнаруживаются в волосковых клетках многих типов и могут участвовать в регуляции притока и оттока Са 2+ из эндолимфы, однако точные механизмы этого еще неизвестны.
Регуляция объема жидкостей улитки также важна для функционирования улитки, описано множество различных механизмов, поддерживающих эту регуляцию. Первоначально, в качестве основных принципов были предположены продольные и радиальные потоки. Продольный ток эндолимфы описывался как ее секреция на протяжении перепончатого лабиринта с реабсорбцией в эндолимфатическом протоке и мешке, в то время как радиальный ток формировался локальной секрецией и реабсорбцией, в частности, посредством сосудистой полоски.
За патологические состояния, такие как увеличение или уменьшение объема эндолимфы, может быть ответственен продольный ток. Было обнаружено, что экспериментальное увеличение эндолимфы вызывало продольный ток эндолимфы по направлению к основанию улитки в эндолимфатический мешок, снижая как объем жидкости, так и концентрацию электролитов в улитковом протоке. С другой стороны, экспериментальное уменьшение объема эндолимфы приводило к току, направленному к верхушке, увеличению объема жидкости и концентрации электролитов. Теория радиального тока никогда не была подтверждена экспериментально.
В настоящее время преобладает мнение, что в физиологических условиях не существует значимого тока эндолимфы. Эксперименты на животных показали, что меченые ионы, помещенные в эндолимфу без нарушения ее объема, двигались исключительно посредством диффузии. Таким образом, ионы эндолимфы циркулировали без формирования объемного тока. Схожие регуляторные механизмы предложены для объяснения гомеостаза перелимфы. Медленный объемный ток внутри улитки играет роль при внутриулитковом введении препаратов, где в физиологических условиях диффузия в отдельных наполненных жидкостью отсеках, по всей видимости, равномерно распределяет концентрации препаратов от основания до верхушки.
На клеточном уровне трансмембранное движение воды во многом зависит от напоминающих поры водопроницаемых каналов, таких как аквапорины. Некоторое количество аквапоринов было обнаружено во внутреннем ухе, но лишь немногие из них локализовались в выстилающем эдролимфу эпителии. Снижение аквапорина-4 у мыши приводило к нарушениям слуха. Мыши с другими модифицированными генами аквапорина либо имели неизвестный на сегодняшний день фенотип, либо погибали в эмбриональном периоде, как и мыши с модифицированным геном аквапорина-2. Тем не менее, аквапорин-2 интересен тем, что он обнаруживается в эпителии, выстилающем эндолимфатический мешок, и регулируется гормоном вазопрессином.
Модели животных показали, что патологически повышенный уровень вазопрессина приводил к выраженному отеку, морфологически характерному для болезни Меньера. Помимо потенциальной роли в патогенезе болезни Меньера данный факт дает основания говорить о гормональном влиянии на регуляцию объема жидкостей внутреннего уха. В частности, вазопрессин, как было показано, не только влияет на мембранную экспрессию аквапорина-2, но также увеличивает активность эпителиальных натриевых каналов и NKCC1 переносчиков, обнаруживаемых в маргинальных клетках сосудистой полоски и фиброцитах II типа спиральной связки. Модуляция этих каналов ведет к увеличению секреции калия в эндолимфу, а впоследствии к изменению осмотического объема, приводя к типичному гидропсу. Сходные данные были получены для другого гормона, альдостерона, который увеличивает активность как эпителиальных натриевых каналов, и Na + /К + -АТФ-азы. Однако, другие гормоны имеют противоположный вазопрессину эффект.
Глюкокортикоиды, в частности, как было показано, подавляют симптомы, возникающие при болезни Меньера, возможно, из-за снижения выработки вазопрессина и модуляции мембранной экспрессии определенных аквапоринов.
Гомеостаз жидкостей улитки, ионный транспорт и внутриулитковый потенциал необходимы для правильного функционирования улитки. Метаболическая блокада или специфическое угнетение ионного транспорта, в частности уабаином или фуросемидом, быстро поражает эту микросреду и нарушает слуховую функцию, снижая заряд улитки, то есть внутриулитковый потенциал. Есть предположение, что старение влияет на внутриулитковый потенциал и возникающее вслед за этим снижение метаболизма сосудистой полоски может играть роль в возрастной потере слуха.

Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Органы чувств
, membrana reticularis. Формируется фаланговыми отростками наружных волосковых клеток и клеток Дейтерса. В нее заходят волоски рецепторных клеток. Рис. Б.
Спиральный сосуд
, vas spiralе. Небольшой кровеносный сосуд в базилярной пластинке под туннелем спирального органа. Рис. А.
Преддверная стенка улиткового протока (преддверная мембрана) [[Рейснера]]
, paries vestibularis ductus cochlearis (membrana vestibularis) [[Reissner]]. Верхняя стенка улиткового протока толщиной около 3 мкм. Рис. А.
Наружная стенка улиткового протока
Базилярный гребешок
, crista basilaris. Возвышение на спиральной связке, которое продолжается в базилярную пластинку. Рис. А.
Спиральный выступ
, prominentia spiralis. Расположен над наружной спиральной бороздой и состоит из соединительной ткани, в которой проходит выступающий сосуд. Рис. А.
Выступающий сосуд
Сосудистая полоска
Спиральный узел улитки
, ganglion spirale cochlearis. Cостоит из биполярных нейронов, которые расположены в спиральном канале улитки. Их периферические отростки берут начало от волосковых клеток, центральные - формируют улитковую часть преддверно-улиткового нерва. Рис. В.
Сосуды внутреннего уха
Артерия лабиринта
, a. labyrinthina. Начинается от базилярной артерии спереди от места отхождения передней нижней мозжечковой артерии, через внутренний слуховой проход, вместе с преддверно-улитковым нервом проникает в пирамиду височной кости,где разветвляется на ветви, кровоснабжающие внутреннее ухо. Рис. В.
Преддверные ветви
, rami vestibulares. Кровоснабжают стенки полукружных протоков, сферического и эллиптического мешочков, а также нижнюю 1/3 первого завитка улиткового протока. Рис. В.
Улитковая ветвь
, ramus cochlearis. Проникает в стержень улитки и кровоснабжает спиральный узел, а также улитковый проток с его содержимым за исключением нижней трети первого завитка. Рис. В.
Артериальные клубочки улитки
, glomeruli arteriosi cochleae. Спиральная артериальная сеть, которая сопровождает спиральную вену стержня. Рис. В.
Вены лабиринта
, vv. labyrinthinae. Сопровождают одноименную артерию, проходят через внутренний слуховой проход и открываются в нижний каменистый синус или во внутреннюю яремную вену. Рис. В.
Спиральная вена стержня
Преддверные вены
, vv. vestibulares. Начинаются от стенок полукружных протоков, эллиптического и сферического мешочков. Впадают в вены лабиринта и вену водопровода преддверия. Рис. В.
Вена водопровода преддверия
, v. aquaeductus vestibuli. Сопровождает эндолимфатический проток и впадает в нижний каменистый синус. Рис. В.
Вена водопровода улитки
, v. aquaeductus cochleae. Проходит в канальце улитки и несет кровь от первого завитка улитки к внутренней яремной вене. Рис. В.
Костный лабиринт
Преддверие
, vestibulum. Часть костного лабиринта, которая содержит эллиптический и сферический мешочки. Рис. Г.
Сферическое углубление
, recessus sphericus. Находится на медиальной стенке преддверия, в котором находится сферический мешочек. Рис. Г.
Эллиптическое углубление
, recessus ellipticus. Находится на медиальной стенке преддверия и содержит часть эллиптического мешочка, расположенную между задней ампулой и общей ножкой. Рис. Г.
Читайте также: