Синтез гастроинтестинальных пептидов и их катаболизм
Добавил пользователь Евгений Кузнецов Обновлено: 09.01.2026
Регуляторные пептиды пищеварительного тракта влияют на пищеварительные функции. Так называемые общие эффекты особенно выражены в изменении обмена" веществ, деятельности сердеч- но-сосудистой и эндокринной систем организма.
. Гастрин усиливает высвобождение гистамина, инсулина, кальцитонина, липолиз в жировой ткани, выделение почками воды, калия, натрия. Соматостатин тормозит высвобождение гастроинтестинальных гормонов, соматотропина, подавляет гликогенолиз, изменяет пищевое по-ведение. ВИП снижает тонус кровеносных сосудов и бронхов. Секретин усиливает липолиз и гликолиз, тормозит реабсорбцию бикарбонатов в поч-ках, увеличивает диурез, ренальное выделение натрия и калия, повышает сердечный выброс. Нейротензин усиливает высвобождение глюкагона, соматостатина, вазопрессина, гистамина, лютропина и фоллитропина, тормозит высвобождение инсулина, усиливает теплопродукцию. Высвобождение регуляторных пептидов и аминов пищ тракта рег-ся гормонами эндокринных желез.
Слюна содержит лизоцим (мурамидаза), которая обладает антибактериальной активностью, участвует в реакциях местного иммунитета, увеличивая продукцию антител, фагоцитов, повышает межклеточную проницаемость, свертываемость крови. Из слюны выделен белок, обладающий свойствами антианемического фактора. Ферменты слюны влияют на микрофлору полости рта, на ее тро-фику. Слюнные железы принимают участие в обеспечении гомеостаза ферментов и гормонов в крови, выделяя их из крови и в кровь. В слюне и железе обнаружен паротин. Он влияет на обмен белков, Са, липидов, гемопоэза, пролиферация хрящей ткани. Желудок влияет на многие непищевар-е ф-ции. Его сок обладает высокой бактерицидностью, содержит антианемический фактор Кастла, про-,антикоагулянты и фибринолитики.
Поджелуд жел-за принимает уч-е в рег-ции микрофлоры кишечника, трофики его слиз-й и скор-ти обновления ее эпителиоцитов.
Слиз об-ка тонкой кишки обладает тромбопластической, антигепариновойи фибринолитической активностью.
ОБМЕН ВЕШЕСТВ И ЭНЕРГИИ
Понятие об обмене в-в.
Ххарактер признака жизни. В результате обмена веществ непрерывно образуются, обновляются и разрушаются клеточные структуры, синтезируются и разрушаются различные химические соединения. В организме динамически уравновешены процессы анаболизма (ассимиляция) — биосинтеза органических веществ, компонентов клеток и тканей, и катаболизма (диссимиляция) — расщепления сложных молекул компонентов клеток. Для возмещения энергозатрат организма, сохранения массы тела и удовлетворения потребностей роста необходимо поступление из внешней среды белков, липидов, углеводов, витаминов, минеральных солей и воды. Это достигается путем питания.
Белкизанимают ведущее место среди органических элементов, на их долю приходится более 50 % сухой массы клетки. Они выполняют ряд важнейших биологических функций. Вся совокупность обмена веществ в организме (дыхание, пищеварение, выделение) обеспечивается деятельностью ферментов, которые являются: белками. Все двигательные функции организма обеспечиваются взаимодействием сократительных белков — актина и миозина. Поступающий с пищей из внешней среды белок служит пластической и энергетической целям. Пластическое значение белка состоит в восполнении и новообразовании различных структурных компонентов клетки. Энергетическое значение заключается в обеспечении организма энергией, образующейся при расщеплении белков. Белки, содержащие весь необходимый набор аминокислот в таких соотношениях, которые обеспечивают нормальные процессы синтеза, являются биологически полноценными.
Азотистый баланс. Это соотношение количества азота, поступившего в организм с пищей и выделенного из него.
Количество азота, поступившего с пищей, всегда больше количества усвоенного азота, так как часть его теряется с калом.
Зная количество усвоенного азота, легко вычислить общее количество усвоенного организмом белка, так как в белке содержится в среднем 16 % азота (1 г азота содержит 6,25 г белка). Следовательно, умножив найденное количество азота на 6,25, можно определить количество усвоенного белка.
Чтобы установить количество разрушенного белка, необходимо знать общее количество азота, выведенного из организма.
У взрослого человека при адекватном питании, как правило, количество введенного в организм азота равно количеству азота, выведенного из организма. Это состояние получило название азотистого равновесия.
Азотистое равновесие может устанавливаться при значительных колебаниях содержания белка в пище.
В случаях, когда поступление азота превышает его выделение, говорят о положительном азотистом балансе. При этом синтез белка преобладает над его распадом.
Когда количество выведенного из организма азота превышает количество поступившего азота, говорят об отрицательном азотистом балансе.-Отрицательный азотистый баланс отмечается при белковом голодании.
Регуляция обмена белков.Нейроэндокринная регуляция обмена белков
осуществляется рядом гормонов.
Соматотропный гормон гипофиза во время роста организма стимулирует увеличение массы всех органов и тканей. Повышается проницаемость клеточных мембран для аминокислот, усиления синтеза РНК в ядре клетки и подавления синтеза катепсинов — внутриклеточных протеолитических ферментов.
Гормоны щитовидной железы — тироксин и трийодтиронин. Стимулируют синтез белка и благодаря этому активизировать рост, развитие и дифференциацию тканей и органов. Гормоны коры надпочечников — глюкокортикоиды (гидрокортизон, кортикостсрон) усиливают распад белков в тканях, особенно в мышечной и лимфоидной.
24. Роль гастроинтестинальных пептидов и аминов
В управлении пищеварительными функциями принимают участие пептиды, которые продуцируются эндокринными клетками самого пищеварительного тракта. Они рассеяны в слизистой оболочке и пищеварительных железах, поэтому их совокупность названа диффузной эндокринной системой. Продукты ее деятельности называют гастроинтестинальными гормонами, регуляторными пептидами желудочно-кишечного тракта.В пищ тракте открыто более 30 рег-х пептидов: 1)гастрин – усил-е секреции желудка(HCl и пепсиногена) и поджелуд жел-ps 2)секретин – усил-е секреции воды и бикарбонатов поджелуд железой 3)Холецистокини(ХЦК) – усил-е моторики желчного пузыря и секреции ферментов поджел-й жел-й 4)Желудочноый(гастральный) ингибирующий пептид(ГИП) – глюкоззависимое усил-е восвобождения роджел-й железой инсулина; тормож-есекреции и моторики желудка 5)панкреатический пептид(ПП) – торм-е секреции ферментов и бикарбонатов поджел-й жел-й,антагонист ХЦК 6)Энтероглюкагон – мобилизация УВ.
Обмен веществ
1. Понятие об обмене в-в.
Ххарактер признака жизни. В результате обмена веществ непрерывно образуются, обновляются и разрушаются клеточные структуры, синтезируются и разрушаются различные химические соединения. В организме динамически уравновешены процессы анаболизма (ассимиляция) — биосинтеза органических веществ, компонентов клеток и тканей, и катаболизма (диссимиляция) — расщепления сложных молекул компонентов клеток. Для возмещения энергозатрат организма, сохранения массы тела и удовлетворения потребностей роста необходимо поступление из внешней среды белков, липидов, углеводов, витаминов, минеральных солей и воды. Это достигается путем питания.
Белки занимают ведущее место среди органических элементов, на их долю приходится более 50 % сухой массы клетки. Они выполняют ряд важнейших биологических функций. Вся совокупность обмена веществ в организме (дыхание, пищеварение, выделение) обеспечивается деятельностью ферментов, которые являются: белками. Все двигательные функции организма обеспечиваются взаимодействием сократительных белков — актина и миозина. Поступающий с пищей из внешней среды белок служит пластической и энергетической целям. Пластическое значение белка состоит в восполнении и новообразовании различных структурных компонентов клетки. Энергетическое значение заключается в обеспечении организма энергией, образующейся при расщеплении белков. Белки, содержащие весь необходимый набор аминокислот в таких соотношениях, которые обеспечивают нормальные процессы синтеза, являются биологически полноценными.
Азотистый баланс. Это соотношение количества азота, поступившего в организм с пищей и выделенного из него.
Количество азота, поступившего с пищей, всегда больше количества усвоенного азота, так как часть его теряется с калом.
Зная количество усвоенного азота, легко вычислить общее количество усвоенного организмом белка, так как в белке содержится в среднем 16 % азота (1 г азота содержит 6,25 г белка). Следовательно, умножив найденное количество азота на 6,25, можно определить количество усвоенного белка.
Чтобы установить количество разрушенного белка, необходимо знать общее количество азота, выведенного из организма.
У взрослого человека при адекватном питании, как правило, количество введенного в организм азота равно количеству азота, выведенного из организма. Это состояние получило название азотистого равновесия.
Азотистое равновесие может устанавливаться при значительных колебаниях содержания белка в пище.
В случаях, когда поступление азота превышает его выделение, говорят о положительном азотистом балансе. При этом синтез белка преобладает над его распадом.
Когда количество выведенного из организма азота превышает количество поступившего азота, говорят об отрицательном азотистом балансе.-Отрицательный азотистый баланс отмечается при белковом голодании.
Регуляция обмена белков. Нейроэндокринная регуляция обмена белков
осуществляется рядом гормонов.
Соматотропный гормон гипофиза во время роста организма стимулирует увеличение массы всех органов и тканей. Повышается проницаемость клеточных мембран для аминокислот, усиления синтеза РНК в ядре клетки и подавления синтеза катепсинов — внутриклеточных протеолитических ферментов.
Гормоны щитовидной железы — тироксин и трийодтиронин. Стимулируют синтез белка и благодаря этому активизировать рост, развитие и дифференциацию тканей и органов. Гормоны коры надпочечников — глюкокортикоиды (гидрокортизон, кортикостсрон) усиливают распад белков в тканях, особенно в мышечной и лимфоидной.
Синтез гастроинтестинальных пептидов и их катаболизм
Гастроинтестинальные пептиды: регуляторные и гормональные
Регуляторные и гормональные гастроинтестинальные пептиды — не синонимы. Первое из этих понятии шире второго, ибо все гормональные пептиды относятся к регуляторным, но не все регуляторные действуют после всасывания в кровь, т. е. гормональным путем. Известны также паракринный способ влияния некоторых регуляторных пептидов (непосредственно в зоне выделения их на расположенные вблизи клеточные группы разных органов пищеварения) и пептидергический (выделение регуляторных пептидов нервными окончаниями пептидергической иннервации, как правило — в том пли ином органе системы пищеварения). Для целого ряда гастроинтестинальных регуляторных пептидов характерен только гормональный тип влияния, они синтезируются исключительно в эндокринных клетках органов пищеварения — гастрин, секретин, мотилин, гастроингибиторный пептид, глюкагон и энтероглюкагон, панкреатический полипептид.
Общим принципом всех эндокринных клеток органов пищеварения является биосинтез в каждой из специфических инкреторных клеток только одного вида гормональноактивного пептида. Исключением являются G — клетки антральной слизистой оболочки, образующие, помимо гастрина, также незначительное количество АКТГ (конечно, удельный вес гастринообразования и синтеза АКТГ этими клетками просто несопоставимы).
Во всех случаях (опять-таки за исключением G-клеток желудка, расположенных в зоне действия соляной кислоты) апикальные поверхности эндокринных клеток пищеварительного тракта состоят из ворсинок, обращенных в просвет его. Это способствует возможности прямого действия пищевых продуктов и пищеварительных секретов на кишечные эндокринные клетки, для которых такое прямое действие b является основным физиологическим стимулом к выделению в кровь гормонов.
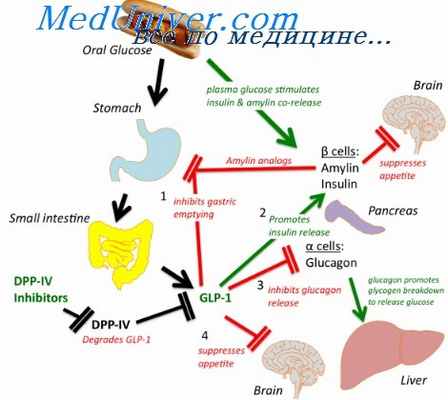
В дополнение к диффузности локализации кишечных инкреторных клеток такой способ побуждения их к функциональной активности отличает эту гормональную систему от классической эндокринной системы. Для отдельных гастропнтестинальных регуляторных пептидов доказано поступление их после биосинтеза в эндокринных клетках не только в кровь (с гормональным эффектом), по и в просвет пищеварительного тракта с пищеварительными секретами — гастрина в составе желудочного сока, соматостатина — с панкреатическим секретом. Сочетание гормонального и паракринного типа действия известно в отношении соматостатина.
Для гастрина, холецистокинина-панкреозимина, соматостатнна множественные молекулярные формы гормонального пептида доказаны с такой определенностью, что эти формы уже выделены и синтезированы. Применительно к другим интестинальным гормональным пептидам множественность структурных форм, наличие молекул-предшественников (прогормоиов) выявлены хроматографнческими исследованиями.
Радиоиммунологические исследования позволяют сейчас уточнить удельный вес каждой из молекулярных форм, циркулирующих в крови, только в отношении отдельных гормонов (гастрина, холецистокинина-панкреозимина). Иммуноцнтохнмнческпе определения дают возможность локализовать биосинтез конкретных гастроннтестпнальных пептидов в той или иной эндокринной клетке, по не дают аргументации для суждении о молекулярных формах образующегося гормона (для этого нужны хроматографические исследования тканевых экстрактов).
Общность молекулярной структуры в определенной мере установлена для гастрина и холецпетокпнина-панкреозимина, но она сама по себе еще не определяет обязательную общность физиологического действия но всем параметрам (панкреотропный эффект холецистокинина-панкреозимина и гастрина принципиально сходен, хотя и различается по силе, а па желудочную секрецию действие обоих гормонов качественно неоднотппно). Предполагается наличие общего прогормона (прегормона) для «семейства» секретина (самого секретина, а также глюкагона и вазоактпвного интестинального пептида). Гастронигибиторный пептид был выделен в процессе очистки холецистокинина-панкреозимина, вазоактивный интестинальный пептид — при очистке секретина. Ведутся поиски информационной РНК применительно к каждому желудочно-кишечному гормональному пептиду, поскольку информационная РНК направляет биосинтез прогормонов, и с ней связаны многие дальнейшие перспективы развития теоретической и практической эндокринологии системы пищеварения.
Синтез пептида прошел на двух цепях РНК без участия рибосомы
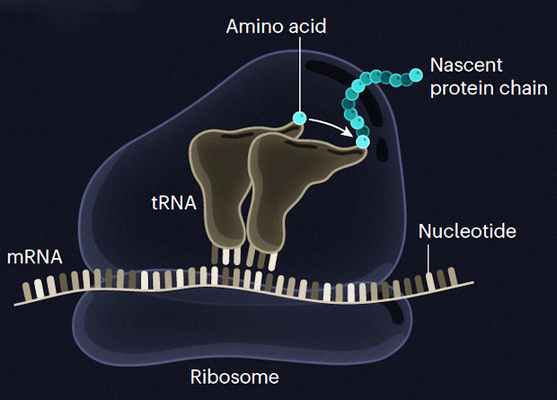
Химики из Германии показали, как мог проходить синтез пептидов до возникновения жизни на Земле. Они выяснили, что на фрагментах транспортных РНК с неканоническими азотистыми основаниями, содержащими остатки аминокислот, могут получаться пептиды без участия рибосом. Исследование опубликовано в журнале Nature.
Молекулы РНК — хранители генетической информации в живых организмах. Они состоят из нуклеотидов, которые содержат азотистые основания — аденин (A), урацил (U), гуанин (G) и цитозин (C). На основе генетического кода, который хранят молекулы РНК, происходит биосинтез белков. При этом для биосинтеза необходима рибосома, именно с ее помощью образуются пептидные связи между аминокислотами, а в результате получается белок.
Так как за счет считывания информации с РНК происходит биосинтез белка, многие биологи считают, что молекулы РНК существовали до появления белков и катализировали синтез сложных органических молекул в пребиотических условиях (гипотеза мира РНК). Но как именно молекулы РНК могли ускорять синтез белков и пептидов, ученым до сих пор неизвестно.
Химики под руководством Томаса Карелла (Thomas Carell) из Мюнхенского университета Людвига и Максимилиана нашли один из возможных механизмов этого процесса. Им было известно, что в состав молекул транспортных РНК, помимо четырех основных азотистых оснований, могут входить несколько неканонических оснований. Причем некоторые из них содержат остатки аминокислот. Поэтому ученые предположили, что из этих аминокислотных остатков в пребиотических условиях могли получаться пептиды.
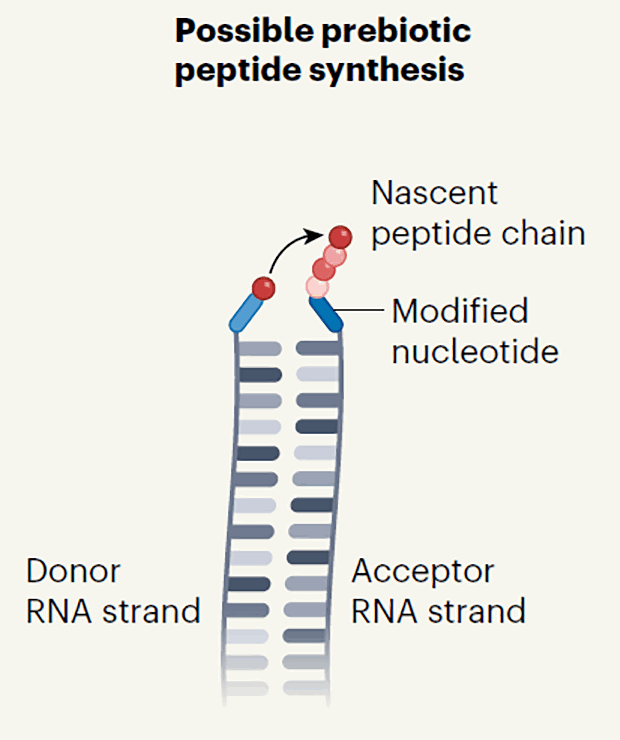
Синтез полипептида на комплементарных цепях РНК без участия рибосомы. Цепи содержат неканонические нуклеотиды
C. Bonfio / Nature, 2022
Чтобы проверить свою гипотезу, химики синтезировали два комплементарных набора цепей тРНК, содержащих неканонические азотистые основания. В одном из наборов неканонические нуклеотиды содержали фрагменты аминокислот со свободной карбоксильной группой, а в другом — свободную аминогруппу. Когда ученые смешали две комплементарные цепи тРНК из двух наборов в водном растворе, между ними образовались водородные связи, а фрагменты аминокислот стали ближе друг к другу.
Затем химики добавили в смесь карбодиимид — активатор карбоксильной группы — и между двумя аминокислотными фрагментами образовалась пептидная связь. В результате дальнейшего нагревания раствора при температуре 90 градусов Цельсия произошел гидролиз, а фрагмент аминокислоты оказался на комплементарной цепи тРНК. Таким образом ученые выяснили, что комплементарные РНК с неканоническими основаниями могут передавать друг другу аминокислотные фрагменты с образованием пептидов.
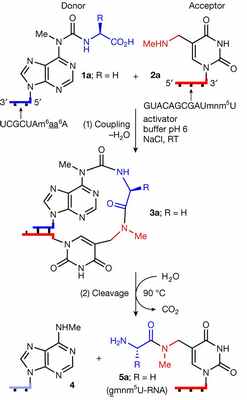
Образование пептидной связи и последующий гидролиз с переносом фрагмента аминокислоты на комплементарную цепь РНК
T. Carell et al. / Nature, 2022
Когда химики попробовали провести эту же цепочку превращений несколько раз, им удалось получить полипептид. Но реакция работала не всегда, а только в случае, если исходные РНК содержали как минимум три нуклеотида и были комплементарны. Интересно, что в обычном биосинтезе белков на рибосомах каждую аминокислоту также кодируют три нуклеотида РНК.
В результате химики показали, что полипептидные цепочки могут образовываться при участии тРНК без рибосом. При этом синтез контролируется комплементарностью молекул тРНК, как и в случае обычного биосинтеза на рибосоме. Реакции образования пептидной связи протекали с выходом около 50 процентов, а выход реакции гидролиза в большинстве случаев не превышал 10 процентов: при нагревании до 90 градусов часть цепей тРНК разрушалась.
Синтез гастроинтестинальных пептидов и их катаболизм
Гормоны. Гидрофильные гормоны
Метаболизм пептидных гормонов
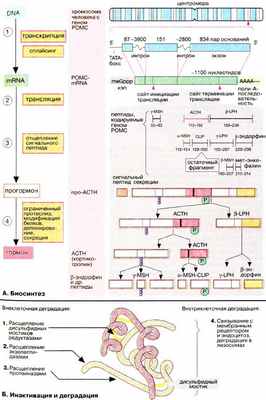
А. Биосинтез
Биосинтез пептидных и белковых гормонов и их секреция находятся под контролем иерархической системы гормональной регуляции (см. с. 360). В этой системе в качестве вторичного мессенджера принимают участие ионы кальция; увеличение концентрации кальция стимулирует синтез и секрецию гормонов.
Анализ гормональных генов показывает, что иногда многие совершенно разные пептиды и белки кодируются одним и тем же геном. Одним из наиболее изученных является ген проопиомеланокортина [ ПОМК (POMC)]. Наряду с нуклеотидной последовательностью, соответствующей кортикотропину [адренокортикотропный гормон, АКТГ (АСТН)], этот ген включает перекрывающиеся последовательности, кодирующие ряд небольших пептидных гормонов, а именно α-, β- и γ- ме ланотропинов [МСГ (MSH)], β- и γ- липотро пинов (ЛПГ (LPH)], β- эндорфина и мет-энке фалина (см. рис. 343). Последний гормон может также образовываться из β-эндорфина. Прогормоном для этого семейства является так называемый полипротеин. Сигнал о том, какой пептид должен быть получен и секретирован, поступает из системы регуляции после завершения синтеза препропептида. Наиболее важным секретируемым продуктом, полученным из гипофизарного полипротеина кодируемого геном ПОМК, является гормон кортикотропин (АКТГ), стимулирующий секрецию кортизола корой надпочечников. Биологические функции других пептидов до конца не выяснены.
Б. Инактивация и деградация
Деградация пептидных гормонов часто начинается уже в крови или на стенках кровеносных сосудов, особенно интенсивно этот процесс идет в почках. Некоторые пептиды, содержащие дисульфидные мостики, например инсулин, могут инактивироваться за счет восстановления остатков цистина ( 1 ), Другие белково-пептидные гормоны гидролизуются протеиназами, а именно экзо- ( 2 ) (по концам цепи) и эндопептидазами ( 3 ). Протеолиз приводит к образованию множества фрагментов, некоторые из которых могут проявлять биологическую активность. Многие белково-пептидные гормоны удаляются из системы циркуляции за счет связывания с мембранным рецептором (см. с. 372) и последующего эндоцитоза гормон-рецепторного комплекса. Деградация таких комплексов происходит в лизосомах, конечным продуктом деградации являются аминокислоты, которые вновь используются в качестве субстратов в анаболических и катаболических процессах.
Липофильные и гидрофильные гормоны имеют различный полупериод существования в системе циркуляции (точнее биохимический полупериод , t 1/2 ). По сравнению с гидрофильными гормонами (t 1/2 несколько минут или часов) липофильные гормоны живут существенно дольше (t 1/2 составляет несколько часов или дней). Биохимический полупериод гормонов зависит от активности системы деградации. Воздействие на систему деградации лекарственными препаратами или повреждение тканей может вызвать изменение скорости распада, а следовательно, и концентрации гормонов.
Читайте также: