Синтез гормонов коры надпочечников. Стероиды - производные холестерола
Добавил пользователь Алексей Ф. Обновлено: 29.01.2026
Являются производными холестерина. Различают половые стероидные гормоны и гормоны коры надпочечников.
Синтез стероидных гормонов происходит путем укорочения боковой цепи холестерина и окисления кольца под действием гидроксилаз с использованием кислорода, витаминов РР и С.
В клубочковой зоне коры надпочечников синтезируется альдостерон, который усиливает реабсорбцию натрия, хлора и воды. Освобождение альдостерона происходит под влиянием ангиотензина-II, который образуется из белка крови ангиотензиногена под влиянием фермента ренина из юкстагломерулярных клеток почек.
Впучковой зонекоры надпочечников образуются глюкокортикосте-роиды (ГКС).
В сетчатой зоне коры образуются слабые андрогены и некоторые эстрогены.
Когда концентрация ГКС снижается, из гипоталамуса освобождаются кортиколиберины, которые приводят к освобождению из гипофиза кортикотропина (АКТГ). АКТГ действует на пучковую и сетчатую зоны коры надпочечников, в результате освобождаются ГКС. Концентрация последних в крови повышается, что прекращает выход кортиколиберинов. В случае, когда гипоталамус не реагирует на высокий уровень ГКС, то есть продолжает выброс кортиколиберинов, возникает болезнь Иценко-Кушинга. В этом случае вся система гипоталамус-гипофиз-кора надпочечников работает чрезмерно. Концентрация ГКС увеличивается. Болезнь Иценко-Кушинга возникает также при опухоли аденогипофиза. АКТГ на клубочковую зону коры влияет мало.
Транспорт ГКС в крови осуществляет белок плазмы транскортин.
Инактивация ГКС происходит в печени путем связывания с глюкуроновой кислотой, а также путем восстановление в неактивные тетрагидропроизводные.
Эффекты ГКС
1. Индуцируют ферменты глюконеогенеза и репрессируют гексокиназу, что приводит к гипергликемии.
2. Усиливают распад белков в мышечной и лимфоидной тканях, тормозят в них синтез.
3. Индуцируют синтез ферментов обмена аминокислот.
4. Усиливают синтез мочевины.
5. Усиливают липолиз, окисление жирных кислот, синтез кетоновых тел и холестерина.
6. Увеличивают прочность сосудов, снижают их проницаемость (снижают активность гиалуронидазы).
7. Увеличивают работоспособность мышц.
8. Усиливают секрецию пищеварительных соков (особенно желудка), но снижают выработку слизи.
9. ГКС необходимы для проявления эффектов катехоламинов.
10.Обладают противоаллергическим действием.
11.Обладают противовоспалительным действием.
Осложнения при стероидной терапии: сахарный диабет, атеросклероз, язвенная болезнь желудка, гипотрофия надпочечников.
1.1. Гормоны - производные полиненасыщенных жирных кислот
Являются производными арахидоновой кислоты. Под действием фосфолипазы происходит расщепление фосфолипидов и освобождение арахидоновой кислоты. Фосфолипазу ингибируют ГКС.
Под действием липооксигеназы в лейкоцитах из арахидоновой кислоты образуются лейкотриены. Это нециклические, окисленные производные арахидоновой кислоты. Лейкотриены действуют через цАМФ.
Эффекты лейкотриенов:
1. Провоспалительное действие.
2. Развитие медленных анафилактических реакций.
3. Освобождение ферментов лизосом.
4. Хемотаксис лейкоцитов.
5. Сокращение гладких мышц.
Под действием циклооксигеназы из арахидоновой кислоты образуются промежуточные биологически активные эндоперикиси ПГG2 и ПГН2. В большинстве тканей из них образуются простагландины Е и F (ПГЕ и ПГF). В стенке сосудов из ПГG2 образуется простациклин (ПЦ), в тромбоцитах – тромбоксаны (ТХ). ПГЕ, ПГF, ПЦ и ТХ являются циклическими, окисленными производными арахидоновой кислоты.
Эффекты ПГЕ и простациклинов:
6. Расслабление гладких мышц.
7. Освобождение гормонов (стероидных, катехоламинов, йодтиронинов, инсулина).
8. Развитие воспаления (провоспалительное действие).
9. Вызывают дезагрегацию тромбоцитов.
10. Вызывают расширение сосудов.
Эффекты ПГF и тромбоксанов:
11. Сокращение гладких мышц.
12. Освобождение гормонов (стероидных, катехоламинов, йодтиронинов, инсулина).
13. Развитие воспаления (провоспалительное действие).
14. Рассасывание желтого тела.
15. Агрегация тромбоцитов.
16. Сужение сосудов.
План изучения темы
Синтез гормонов коры надпочечников. Стероиды - производные холестерола
Синтез гормонов коры надпочечников. Стероиды - производные холестерола
а) Синтез и секреция гормонов коры надпочечников. В коре надпочечников есть три отдельные зоны. На рисунке ниже показано, что кора надпочечников подразделяется на три относительно независимых слоя.
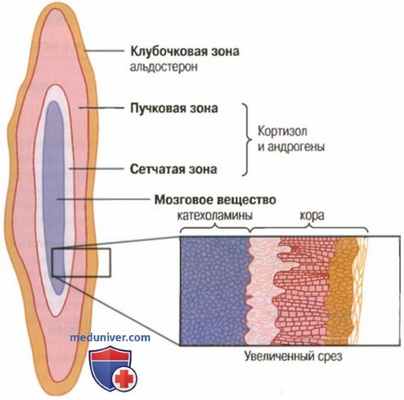
Секреция адренокортикальных гормонов различными зонами коры и секреция катехоламинов мозговым веществом
1. Клубочковая зона — тонкий слой клеток, расположенный непосредственно под капсулой надпочечника, составляющий 15% коры надпочечников. Только клетки этой зоны способны синтезировать существенное количество алъдостерона благодаря присутствию фермента алъдостеронсинтетазы, необходимого для образования альдостерона.
Секреторные процессы этой зоны контролируются главным образом концентрацией ангиотензина II и калия во внеклеточной жидкости, стимулирующих продукцию альдостерона.
2. Пучковая зона — средняя, наиболее широкая зона, которая составляет 75% коры надпочечников и продуцирует глюкокортикоиды, кортизол и кортикостерон наряду с небольшим количеством андрогенов и эстрогенов. Секреция этой зоны коры надпочечников регулируется гипоталамо-гипофизарной системой посредством адренокортикотропного гормона (АКТГ).
3. Сетчатая зона — глубинная зона коры надпочечников, продуцирующая андрогены дегидроэпиандростерон и андростендион наряду с небольшими количествами эстрогенов и некоторых глюкокортикоидов. Регулируют активность этих клеток также АКТГ и другие факторы, например гормон-стимулятор кортикальных андрогенов, выделяемый гипофизом. Механизмы, контролирующие продукцию андрогенов, не так понятны, как механизмы, ответственные за продукцию глюко- и минералокортикоидов.
Продукция альдостерона и кортизола регулируется независимо друг от друга. Так, ангиотензин II, особенно активно стимулирующий выброс альдостерона и благодаря которому наблюдается гипертрофия клубочковой зоны, неэффективен по отношению к двум другим зонам.
Сходным образом АКТГ, повышающий секрецию кортизола и андрогенов надпочечниками, вызывающий гипертрофию пучковой и сетчатой зон, оказался малоэффективным или даже не действенным по отношению к клубочковой зоне.
б) Гормоны коры надпочечников - стероиды, производные холестерола. Стероидные гормоны человека, включая гормоны коры надпочечников, синтезируются из холестерола. Приблизительно 80% холестерола, используемого для синтеза стероидов, обеспечиваются липопротеинами низкой плотности, циркулирующими в плазме крови, хотя клетки коры надпочечников могут заново синтезировать небольшое количество холестерола из ацетата.
ЛПНП, содержащие холестерол в высокой концентрации, диффундируют из плазмы в интерстициальное пространство и прикрепляются к рецепторам, содержащимся в структурах, названных окаймленными ямками, на мембранах адренокортикальных клеток. Затем окаймленные ямки погружаются внутрь путем эндоцитоза, образуя везикулы, сливающиеся в итоге с лизосомами клеток, и высвобождают холестерол, используемый для синтеза стероидных гормонов.
Транспорт холестерола в адреналовые клетки регулируется механизмом обратной связи, который может заметно менять количество имеющегося холестерола для синтеза гормонов. Например АКТГ, который стимулирует синтез стероидных гормонов надпочечников, увеличивает количество рецепторов для ЛПНП наряду с активацией ферментов, высвобождающих холестерол их ЛПНП.
Как только холестерол попадает в клетку, он доставляется в митохондрии, где расщепляется ферментом холестеролдесмолазой, образуя прегненолон; этот этап лимитирует скорость возможного образования стероидов (для облегчения понимания просим вас изучить рисунок ниже).
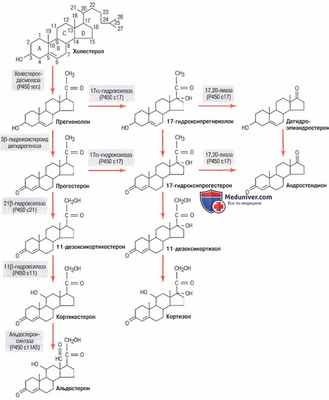
Пути синтеза стероидных гормонов коры надпочечников. Ферменты выделены серым фоном
Во всех трех зонах коры надпочечников этот начальный этап синтеза стероидов стимулируется разными факторами, что определяет в итоге, какие гормоны будут преобладать: альдостерон или кортизол.
Например, как АКТГ, стимулирующий секрецию кортизола, так и ангиотензин II, стимулирующий секрецию альдостерона, ускоряют превращение холестерола в прегненолон.
Видео физиология гормонов надпочечника - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Гормоны гипофизарно-надпочечниковой системы
К гормонам гипофизарно-надпочечниковой системы относят адренокортикотропный гормон (АКТГ), регулирующий активность коркового слоя надпочечников, и глюкокортикоиды (кортикостероиды), синтезируемые в этом корковом слое. Определение количества гормонов этой системы используется для диагностики причин и понимания патогенеза различных заболеваний.
Проопиомеланокортин представляет собой пептид, включающий 254 аминокислоты. При его гидролизе в клетках переднего и промежуточного гипофиза образуется ряд гормонов: α-, β-, γ-меланоцитстимулирующие гормоны, адренокортикотропный гормон, β-, γ-липотропины, эндорфины, мет-энкефалин.
Адренокортикотропный гормон
Строение
Представляет собой пептид, включающий 39 аминокислот.
Регуляция синтеза и секреции
Активируют : кортиколиберин, синтезируемый при стрессе (тревога, страх, боль) под влиянием нервных сигналов от структур головного мозга, вазопрессин, ангиотензин II, катехоламины.
У АКТГ также имеется собственная суточная ритмичность - максимальная концентрация в крови достигается в утренние часы, минимальная в полночь.
Уменьшают : глюкокортикоиды (обратная отрицательная связь).
Механизм действия
Мишени и эффекты
В жировой ткани стимулирует липолиз.
В надпочечниках (главным образом, в сетчатой и пучковой зонах) стимулирует образование белка и нуклеиновых кислот для роста их ткани, активирует синтез холестеролаde novo и его получение из эфиров, усиливает синтез прегненолона.
Патология
Гипофункция
Возможна при гипофизарной недостаточности, сопровождается снижением активности коры надпочечников.
Гиперфункция
Проявляется болезнью Иценко-Кушинга – симптомы гиперкортицизма (см ниже) и специфичные симптомы:
- активациялиполиза,
- увеличение пигментации кожи из-за частичного меланоцитстимулирующего эффекта, благодаря чему появился термин "бронзовая болезнь".
Глюкокортикоиды
Строение
Глюкокортикоиды являются производными холестерола и имеют стероидную природу. Основным гормоном у человека является кортизол.
Строение глюкокортикоидов
Синтез
Осуществляется в сетчатой и пучковой зонах коры надпочечников. Образованный из холестерола прогестерон подвергается окислению 17-гидроксилазой по 17 атому углерода. После этого в действие последовательно вступают еще два значимых фермента: 21-гидроксилаза и 11-гидроксилаза . В конечном итоге образуется кортизол.
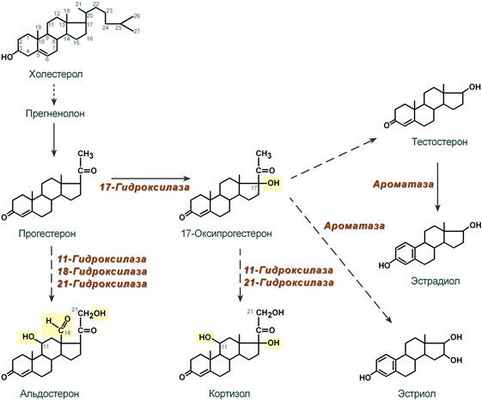
Схема синтеза стероидных гормонов (полная схема)
Регуляция синтеза и секреции
Активируют : АКТГ, обеспечивающий нарастание концентрации кортизола в утренние часы, к концу дня содержание кортизола снова снижается. Кроме этого, имеется нервная стимуляция секреции гормона.
Уменьшают : кортизол по механизму обратной отрицательной связи.
Механизм действия
Мишени и эффекты
Мишенью является лимфоидная, эпителиальная (слизистые оболочки и кожа), жировая, костная и мышечная ткани, печень.
Белковый обмен
- значительное повышение катаболизма белков в лимфоидной, эпителиальной, мышечной, соединительной и костной тканях,
- в печени в целом стимулирует анаболизм белков (например, ферменты трансаминирования и глюконеогенеза),
- стимуляция реакций трансаминирования через синтез аминотрансфераз, обеспечивающих удаление аминогрупп от аминокислот и получение углеродного скелета кетокислот,
Углеводный обмен
В целом вызывают повышение концентрации глюкозы крови:
- усиление мощности глюконеогенеза из кетокислот (полученных из аминокислот) за счет увеличения синтеза фосфоенолпируват-карбоксикиназы,
- увеличение синтеза гликогена в печени за счет активации фосфатаз и дефосфорилирования гликогенсинтазы.
- снижение проницаемости мембран для глюкозы в инсулинзависимых тканях.
Липидный обмен
- стимуляция липолиза в жировой ткани благодаря увеличению синтеза ТАГ-липазы, что усиливает эффект АКТГ, СТГ, глюкагона, катехоламинов, т.е. кортизол оказывает пермиссивное действие (англ. permission - позволение).
Водно-электролитный обмен
- слабый минералокортикоидный эффект на канальцы почек вызывает реабсорбцию натрия и потерю калия,
- потеря воды в результате подавления секреции вазопрессина и излишняя задержка натрия из-за увеличения активности ренин-ангиотензин-альдостероновой системы.
Противовоспалительное и иммунодепрессивное действие
- увеличение перемещения лимфоцитов, моноцитов, эозинофилов и базофилов в лимфоидную ткань,
- повышение уровня лейкоцитов в крови за счет их выброса из костного мозга и тканей,
- подавление функций лейкоцитов и тканевых макрофагов через снижение синтеза эйкозаноидов посредством уменьшения транскрипции ферментов фосфолипазы А2 и циклооксигеназы.
Другие эффекты
Повышает чувствительность бронхов и сосудов к катехоламинам, что обеспечивает нормальное функционирование сердечно-сосудистой и бронхолегочной систем.
Инактивация кортизола
Деактивация кортизола, как и других стероидных гормонов, происходит в печени. Суть реакций заключается
- в восстановлении двойной связи в А-кольце и оксогруппы в 3-м положении,
- в отщеплении радикала от 17-го атома углерода,
- в конъюгации ОН-групп с серной или глюкуроновой кислотами с образованием гидрофильных соединений.
В результате деактивации образуются разнообразные соединения с резко пониженной гормональной активностью или совсем лишенные таковой.
Патология
Гипофункция
Причиной первичной недостаточности ( болезнь Аддисона , бронзовая болезнь ) является разрушение коры надпочечников при кровоизлиянии или травмах, аутоиммунные процессы, туберкулез, гемохроматоз. Гипокортицизм проявляется как:
- гипогликемия,
- повышенная чувствительность к инсулину,
- анорексия и снижение веса,
- слабость,
- гипотензия,
- гипонатриемия и гиперкалиемия,
- усиление пигментации кожи и слизистых (компенсаторное увеличение количества АКТГ, обладающего небольшим меланотропным действием).
Вторичная недостаточность возникает при дефиците АКТГ или снижении его эффекта на надпочечники – возникают все симптомы гипокортицизма, кроме пигментации.
Гиперфункция
Первичная гиперфункция ( синдром Кушинга , синдром гиперкортицизма, стероидный диабет) возникает при доброкачественных или злокачественных опухолях коры надпочечников. Он проявляется как:
- снижение толерантности к глюкозе – аномальная гипергликемия после сахарной нагрузки или после еды,
- гипергликемия из-за активации глюконеогенеза,
- ожирение лица и туловища (связано с повышенным влиянием инсулина при гипергликемии на жировую ткань) – буйволиный горбик, фартучный (лягушачий) живот, лунообразное лицо,
- глюкозурия,
- повышение катаболизма белков и повышение азота крови,
- остеопороз и усиление потерь кальция и фосфатов из костной ткани,
- снижение роста и деления клеток – лейкопения, иммунодефициты, истончение кожи, язвенная болезнь желудка и двенадцатиперстной кишки,
- нарушение синтеза коллагена и гликозаминогликанов,
- гипертония благодаря активации ренин-ангиотензиновой системы.
Вторичная гиперфункция ( болезнь Иценко-Кушинга ) связана с избыточным синтезом АКТГ при микроаденоме гипофиза и проявляется схоже с первичной формой.
Гормоны коры надпочечников
В коре надпочечников синтезируются стероидные гормоны. Это соединения липидной природы, производные циклопентанопергидрофенантрена. Стероидные гормоны синтезируются, кроме надпочечников, в половых железах, однако независимо от места синтеза их общим предшественником является холестерин. Кортикостероиды коры надпочечников разделяют на две группы: глюкокортикоиды и минералокортикоиды. Глюкокортикоиды контролируют многие стороны обмена углеводов, липидов и нуклеиновых кислот. Наибольшей активностью обладают такие представители этой группы, как кортизол и кортикостерон.
Минералокортикоиды оказывают существенное влияние на водно-солевой обмен, причем наиболее активным из них является альлостерон. Гормонам каждой группы свойственна (в меньшей степени) биологическая активность гормонов другой группы (табл. 13.1).
Таблица I3.1. Биологическая активность кортикостероидов (Bagavan N. V., 1978)
Г люкокортикоидная активность
Биосинтез. Образование кортикостероидов осуществляется в несколько стадий, причем общим предшественником их является холестерин (гл. 23). Холестерин синтезируется в надпочечниках или же поступает в них из кровяного русла. В цитоплазме клеток происходит этерификация холестерина и его депонирование. Сигнал на синтез кортикостероидов формируется в гипоталамусе и реализуется в синтезе кортиколиберина. Этот гормон, воздействуя на гипофиз, стимулирует образование адренокортикотропного гормона (АКТГ). Последний, взаимодействуя с мембранными рецепторами клеток надпочечников, через систему вторичных посредников активирует эстеразу холестсрола; при этом освободившийся холестерол транспортируется в митохондрии. Превращение холестерола в прегненолон в митохондриях происходит в результате гидроксилирования и отщепления боковой цепи посредством ферментов десмолазного комплекса, включающего в себя 20- и 22-гидроксилазы, а также С20_22"Л иаз у. В реакциях гидроксилирования принимают участие цитохром Р-450 и НАДФН. В результате образуется С21-стероид, который носит название прегненолон:
Идентифицировано несколько путей дальнейшего превращения прегне- нолона в биологически активные гормоны — кортикостероиды. Один из них связан с превращением прегненолона:
В ином варианте из прогестерона образуется кортизол по схеме:
Синтез минералокортикоидов контролируется ренин-ангиотензиновои системой, основным компонентом которой является ангиотензин-П-октапептид. образующийся из пол и пептидного предшественника.
Возможен путь образования из прегненолона оксикортикостерона, а затем минералокортикоида — альдостерона:
Регуляция биосинтеза. Образование кортикостероидов имеет многоуровневый характер. Прежде всего следует отметить регуляцию, связанную с сигналами, поступающими из гипоталамуса и гипофиза. Далее существенное влияние на этот процесс оказывает содержание холестерола и его транспорт в митохондрии. И наконец, регуляция образования кортикостероидов определяется активностью ферментов гидроксилирования холестерина. Образование прегненолона является лимитирующей стадией всего процесса стероидогене- за. Был обнаружен специальный белок, способствующий взаимодействию холестерина с цитохромом Р-450 и, таким образом, оказывающий существенное влияние на стероидогенез.
Метаболизм. Биотрансформация глюкокортикоидов происходит в печени и заключается в серии реакций окисления и восстановления (причем последние превалируют). Одной из основных реакций инактивации этих гормонов является образование восстановленных дигидро- и тетрагидропроизводных в результате восстановления двойных связей в кольце А в присутствии НАДФН. Биотрансформация включает также конъюгацию с глюкуроновой кислотой и в меньшей степени с сульфатами. Образовавшиеся конъюгаты с желчью поступают в кишечник, где возможна их реадсорбция, попадание в кровяное русло и выведение с мочой.
Минералокортикоиды. Эти гормоны в печени превращаются в тетрагидропроизводные. Например, альдостерон восстанавливается до тетрагидроаль- достсрона, который затем образует конъюгаты с глюкуроновой кислотой и выводится из организма с мочой.
Биохимические функции. Глюкокортикоиды стимулируют катаболиче- ские процессы в организме, преимущественно в мышечной и жировой тканях. Новосинтезированные гормоны быстро секретируются в кровь и связываются со специфическим белком — транскортином. Образованный макромолеку- лярный комплекс переносится к клеткам-мишеням, где происходит его диссоциация и реализация действия гормонов. Глюкокортикоиды усиливают распад белков, повышают содержание аминокислот в крови и аминного азота в моче. Данные гормоны ингибируют синтез нуклеиновых кислот во всех тканях, кроме печени. Их действие на углеводный обмен проявляется прежде всего в увеличении глюкозы в крови за счет активации глюконеогенеза в печени. В липидном обмене глюкокортикоиды стимулируют интенсификацию липолиза, а также ингибируют синтез жирных кислот в печени.
Минералокортикоиды, воздействуя на почки, регулируют водно-солевой обмен в организме. Самым активным в этой группе гормонов является альдостерон, обеспечивающий транспорт Na~ в почечных канальцах. Кроме того, он стимулирует выделение с мочой К + и иона аммония. Механизм действия альдостерона связан с увеличением числа натриевых каналов в мембранах почечных клеток, а также с индукцией синтеза АТФ, необходимого для транспорта ионов.
Практическое применение. Кортикостероиды проявляют антивоспали- тельную, антиаллергическую и иммунодепрессивную активность. Эти фармакологические эффекты обусловливают их применение в качестве лекарственных препаратов. Их применяют для лечения ревматизма, ревматоидных артритов, бронхиальной астмы, лейкозов, аллергических реакций и ряда других заболеваний. Кроме того, кортикостероиды используют в заместительной терапии, например при болезни Аддисона, а также для подавления иммунитета при пересадке органов. Лекарственные формы — ампулы для инъекций, таблетки, мази.
Синтез стероидных гормонов и их функции
Государственный Медицинский Университет г.Семей.
Кафедра биохимии и химических дисциплин.
Дисциплина: Медицинская биохимия.
СРС:
Синтез стероидных
гормонов и их функции.
Выполнил: Есентаев Н.А.
217гр. ОМФ
Проверила: Прозор И.И.
Г.Семей. 2017г.
План:
-Введение;
-Общие сведения о гормонах;
-Классификация гормонов;
-Синтез стероидных гормонов;
-Функции стероидных гормонов;
-Заключение;
-Список литературы.
Введение:
Гормоны - это биологически активные вещества, которые
синтезируются в малых количествах в
специализированнных клетках эндокринной системы и
через циркулирующие жидкости (например, кровь)
доставляются к клеткам-мишеням, где оказывают свое
регулирующее действие.
Особенности гормонов:
1. Синтез гормонов происходит в клетках
эндокринной системы. При этом синтез гормонов
является основной функцией эндокринных клеток.
2. Гормоны секретируются в кровь, чаще в венозную,
иногда в лимфу.
3. Телекринный эффект (или дистантное действие) гормональное действие на клетки-мишени
осуществляется на большом расстоянии от места
синтеза гормонов.
Строение гормонов:
Строение гормонов бывает разным. В настоящее время
описано и выделено около 160 различных гормонов из
разных многоклеточных организмов.
По химическому строению гормоны можно
классифицировать по трем классам:
1. белково-пептидные гормоны;
2. производные аминокислот;
3. стероидные гормоны.
Общие данные о гормонах:
К белково-пептидным относятся гормоны гипоталамуса и
гипофиза, а также гормоны поджелудочной и паращитовидной
желез и один из гормонов щитовидной железы.
К производным аминокислот относятся амины, которые
синтезируются в мозговом слое надпочечников и в эпифизе, а
также йодсодержащие гормоны щитовидной железы.
Стероидные гормоны - это гормоны, которые синтезируются в
коре надпочечников и в половых железах. По количеству
углеродных атомов стероиды отличаются друг от друга:
1. С21 - гормоны коры надпочечников и прогестероны;
2. С19 - мужские половые гормоны - андрогены и тестостерон;
3. С18 - женские половые гормоны - эстрогены.
Общим для всех стероидов является наличие стеранового ядра.
Синтез стероидных гормонов:
1. Стероидные гормоны синтезируются из холестерина (27
углеродных атомов), который поступает в стероидогенные клетки из
крови или синтезируется из ацетил-КоА.
2. В этих клетках холестерин в виде эфиров с жирными кислотами
накапливается в липидных каплях цитозоля, откуда мобилизуется
под воздействием холестераз и поступает в митохондрии.
3. Во внутренних мембранах митохондрий боковая цепь укорачивается,
а затем под воздействием ферментов адренодоксина и цитохрома
Р450 в присутствии НАДН2 и О2 гидроксилируется в прегненолон.
4. Прегненолон выводится из митохондрий и из него в микросомах
синтезируется прогестерон.
5. Далее из прогестерона синтезируются глюкокортикоиды и
минералокортикоиды.
Лимитирующим этапом биосинтеза стероидных
гормонов является переход холестерина в
митохондрии и его превращение в прегненолон.
Этот этап специфически контролируется в
надпочечниках адренокортикотропным гормоном
(АКТГ), усиливающим процесс разрушения эфиров
холестерина посредством холестераз и ускоряющим
вовлечение холестерина в митохондрии, а также
последующий синтез стероидов. АКТГ стимулирует
синтез глюкокортикоидов, частично андрогенов и в
небольшой степени влияет на синтез альдостерона.
Усиливать биосинтез кортикостероидов и особенно
альдостерона способны и простагландины.
Синтез женских половых гормонов:
Эстрогены синтезируются в клетках внутренней
оболочки фолликул и частично синтезируются в
гранулезных клетках, выстилающих полость фолликул
яичников, в желтом теле и плаценте. Определенное
количество этих гормонов синтезируются
лейдиговскими клетками семенников и клетками
сетчатой зоны коры надпочечников.
Исходным соединением для биосинтеза эстрогенов
служит холестерин, преобразуемый в прегненолон,
прогестерон, андрогены и затем в эстрогены.
Важнейшими стадиями формирования специфической
структуры эстрогенов (С18 стероиды) являются:
1. Отщепление у С19 в составе метильной группы от
молекулы тестостерона, с образованием С19 –
нортестостерона.
2. Ароматизация кольца А с образованием
эстрадиола и эстрона.
Эстрадиол и эстрон – основные эстрогены, могут
взаимно превращаться, и синтезируются в яичниках и
тестикулах.
В плаценте, наряду с эстрогенами, из андрогенов,
поступающих из организма плода, синтезируется
третий представитель эстрогенов – эстриол.
Синтез эстрогенов
Биосинтез эстрогенов регулируется лютеонизирующим
гормоном (ЛГ). В яичниках эффект ЛГ усиливается
фолликулостимулирующий гормон (ФСГ) и пролактином
(лютеотропный гормон -ЛТГ). В плаценте синетез эстрогенов
контролируется хорионическим гонадотропином (ХГТ), хотя на
ранних стадиях беременности ХГТ может активизировать
образование эстрогенов и желтым телом яичников. Секреция
эстрогенов эмбриональными яичниками млекопитающих имеет
второстепенное значение для формирования женского полового
аппарата в эмбриогенезе, но в то же время эстрогены активно
задерживают во времени маскулинизацию гипоталамических
центров, оказывая непосредственное влияние на
дифференцировку преоптической зоны.
Функции эстрогенов
В постэмбриональный период эстрогены играют решающую
роль в окончательной феминизации организма, воздействуя на
процессы половой дифференцировки репродуктивного аппарата
и на формирование особенностей структуры и функций
различных органов и тканей.
В половозрелом женском организме эстрогены совместно с
прогестероном обеспечивают ритмичность женских половых
циклов, основное содержание которых – ритмическая смена
двух альтернативных процессов:
Готовность женского организма к половому акту и
оплодотворению яйцеклетки.
Обеспечение развития оплодотворенной яйцеклетки,
отрицающее, как правило, возможноть дополнительного
спаривания и оплодотворения.
Эстрогены обеспечивают следующие изменения структуры
полового тракта:
1. Усиливают синтез РНК, ДНК и белков в матке, идет увеличение
массы матки за счет роста миометрия и пролиферации
эндометрия.
2. Обеспечивают стадии проэструса и эструса, стимулируя
гипертрофию и гиперплазию эпителиальных клеток влагалища с
образованием наружного ороговевающего слоя.
3. Стимулируют рост яйцеводов и их сократимость, что
обеспечивает возможность продвижения яйцеклетки к матке.
4. Усиливают продукцию секрета полового тракта и поддерживают
рН секрета влагалища в пределах от 4,5 до 5,0.
5. Стимулируют рост и развитие яичников – формирование
фолликулов, их кровоснабжение и повышают реактивность
фолликулярного аппарата к пролактину и ФСГ.
6. По принципу положительной обратной связи индуцируют
выброс гипофизом ЛГ.
7. Стимулируют разрастание и разветвление молочных протоков,
повышают чувствительность альвеолярного аппарата к
прогестерону и пролактину (ЛГ)
В не репродуктивных тканях женского и мужского организма
эстрогены оказывают анаболический эффект, ускоряя синтез
РНК, ДНК и белков, а также синтез жирных кислот и
фосфолипидов. Эстрогены ускоряют окисление углеводов по
пентозному пути и накопление НАДФН2 и замедляют синтез
холестерина, снижают выделение ионов Са++, ускоряя их
отложение в костной ткани.
В печени эстрогены стимулируют синтез белков,
участвующих в специфическом связывании и транспорте
гормонов (транскортин, секс-стероид-связывающий
глобулин, тироксин-связывающий глобулин и другие), синтез
ангиотензиногена, протромбина, пре – бета – и бета –
липопротеидов. В почках стимулируют синтез ренина и
способствуют выведению натрия с мочей. В мужском
организме усиливают рост простаты и семенных пузырьков.
Синтез прогестерона
Прогестерон синтезируется в желтом теле яичников и
плаценте. Определенное количество прогестинов
образуется в фолликулах яйцеклетки и лейдиговских
клетках семенников. В коре надпочечников
прогестины являются промежуточными продуктами на
пути биогенеза кортикостероидов из холестерина. В
желтом теле яичников прогестерон также
синтезируется из холестерина через стадию
образования прегненолона.
Функция прогестерона
Прогестерон и его аналоги играют второстепенную роль в процессах
полового созревания. Основная их роль в развивающемся организме
млекопитающих сводится к стимуляции дифференцировки
альвеолярного аппарата молочных желез, а также к антиэстрогенным
и антиандрогенным эффектам.
Прогестероны подготавливают матку к беременности,
предотвращают течку, тормозят активность циклического центра
гипоталамуса, созревание фолликул в яичнике и тормозят овуляцию.
В этот период желтое тело продуцирует прогестерон около 20 мг в
сутки. С наступлением берменности синтез прогестерона резко
возрастает (100 мг в сутки) и достигает максимума (250 мг в сутки) к
концу беременности.
Участие прогестерона в подготовке к беременности
состоит в том, что этот гормон стимулирует
разрастание эндометрия, стимулирует выведение
железистыми клетками слизистого секрета,
необходимого для продвижения и имплантации
оплодотворенной яйцеклетки в эндометрий.
В стадию диэструса (межтечка) – стадия покоя
вагинального эпителия, гестагены тормозят
разрастание, стратификацию и ороговение
вагинального эпителия, стимулирируют секрецию
слизи поверхностным слоем клеток, который
препятствует продвижению сперматозоида в матку.
Заключение:
Из всего вышесказанного следует, что биосинтез
стероидных гормонов жизненно важный процесс без
которого невозможно нормальное развитие
человеческого организма.
Данная тема может нам пригодится в дальнейшей
практике для правильного понимания и
интерпретирования различных клинических случаев.
Список литературы:
1.
Тапбергенов С.О., Тапбергенов Т.С. Медицинская и клиническая биохимия.
-Алматы: ТОО “Эверо”, 2012.
2.
Березов Т.Т., Коровкин Б.Ф. Биологическая химия.- М.,2005
3.
Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека. 2003
4.
А.М. Чибиряев "Биологически активные соединения живых организмов",
2009
5.
Физиология человека (в 3-х томах). Ред. Шмидт Р., Тевс Г. М., Мир, 1996.
6.
Татарчук Т. Ф., Гуньков С. В., Ефименко О. А. Репродуктивная
эндокринология. — 2012.
7.
Большая советская энциклопедия : гл. ред. А. М. Прохоров. — 3-е изд. —
М. : Советская энциклопедия, 1969—1978.
Читайте также:
- Топография межреберных артерий. Межреберные артерии. Межреберные нервы. Внутригрудная фасция.
- Неотложная помощь при остром коронарном синдроме.
- Лимфоматоидный и аллергический гранулематоз. Генерализованный узелковый полиартериит
- Сахарный диабет тучных и тощих. Кожный зуд при диабете
- Пальпация височно-нижнечелюстного сустава спереди