Строение и функции ядерной ламины
Добавил пользователь Евгений Кузнецов Обновлено: 10.01.2026
Строение и функции ядерной ламины
• Ядерная ламина состоит из белков промежуточных филаментов, называемых ламинами
• Ядерная ламина расположена за внутренней ядерной мембраной и физически связана с ней интегральными мембранными белками
• Ядерная ламина участвует в сборке ядерной оболочки и может выполнять функции ее физической поддержки
• Ядерная ламина связана с хроматином посредством белков. Это может обусловливать ее роль в репликации и транскрипции ДНК
• У дрожжей и некоторых других одноклеточных эукариот ядерная ламина отсутствует
Наличие ядерной ламины — сети промежуточных филаментов, расположенной за внутренней ядерной мембраной, является характерным признаком, свойственным ядрам клеток Metazoa. Как показано на рисунке ниже, ламина легко выявляется методом непрямой иммунофлюоресценции, с использованием антител к одному из белков, входящих в ее состав.
В электронном микроскопе ламина выглядит как сеть, состоящая из отдельных волокон. На рисунке также представлены типичные неупорядоченные филаменты ламины, расположенные прямо за внутренней ядерной мембраной.
Белки ядерной ламины (называемые ламины) напоминают кератиновые белки промежуточных филаментов цитоплазмы. Так же как и кератины, они называются белками промежуточных филаментов, поскольку образуемые ими филаменты (10-20 нм) по размеру занимают промежуточное положение между актиновыми микрофиламентами (7 нм) и микротрубочками (25 нм). Целостность филаментов ламины нарушается ЯПК, которые прикрепляются к ней.
Наряду с белками промежуточных филаментов, ядерная ламина также содержит набор интегральных мембранных белков, которые называются белками, связанными с ламиной (LAP). Некоторые из этих белков участвуют во взаимодействиях между ламиной и внутренней мембраной ядра. Как показано на рисунке ниже, ламина закрепляется на внутренней ядерной мембране двумя типами связей. Один тип образуется между белками ламины и интегральными белками внутренней мембраны ядра Другой тип возникает при взаимодействии полипептидной цепи ламинов с липидной фарнезильной группой внутренней ядерной мембраны.
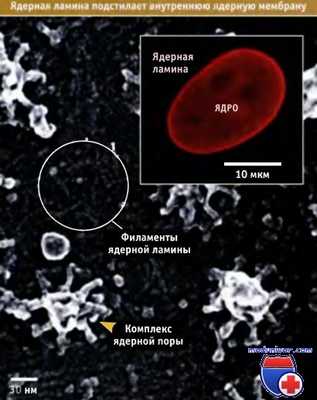
Для визуализации ядерной ламины с помощью флуоресцентной микроскопии использовали антитела к белкам ламины (в рамке).
На фотографии, полученной с помощью электронного микроскопа, видны ядерные корзины ядерных поровых комплексов (ЯПК) и филаменты ядерной ламины.
Геном растений не содержит генов ядерных ламинов, однако растения содержат другие структурные белки, которые выполняют такие же функции, как и ламины клеток млекопитающих. Дрожжи (S. cerevisiae и S. pombe) и некоторые другие одноклеточные эукариоты не содержат ламинов, и поэтому не образуют ламины. С чем это может быть связано?
Существуют, по меньшей мере, две возможные причины, объясняющие отсутствие ламины у одноклеточных различием в величине ядер у клеток дрожжей (около 1 мкм в диаметре) и многоклеточных организмов (порядка 10 мкм в диаметре). Первая возможность заключается в том, что для дрожжевых клеток характерен закрытый митоз, при котором ядерная оболочка в течение всего времени остается интактной. Напротив, в клетках многоклеточных эукариот ядерная оболочка разрушается в начале митоза. После сегрегации хромосом вокруг каждого их набора образуется новая ядерная оболочка. Считается, что в это время ламина играет главную роль в перестройке организации ядра.
Во-вторых, ламина может обеспечивать дополнительную структурную поддержку для более крупной ядерной оболочки в клетках Metazoa.
Наряду с той ролью, которую ядерная ламина выполняет в сборке и поддержании структуры ядра, ее взаимодействие с хроматином может представляться необходимым для протекания репликации ДНК. Данные, свидетельствующие о роли ядерной ламины в репликации, были получены в экспериментах, в которых хроматин сперматозоидов Xenopus laevis добавляли к экстрактам яйцеклеток. При этом вокруг хроматина отмечалось образование ядерной оболочки.
Эти ядра увеличивались в размерах, и происходила деконденсация хромосом, находившихся в ядрах сперматозоидов в конденсированном состоянии (деконденсация маскирует картину, которая наблюдается при оплодотворении, когда ДНК сперматозоида взаимодействует с некоторыми факторами в ооцитах). Затем в этих ядрах происходит репликация ДНК. В экстрактах яйцеклеток в большом количестве содержатся ядерные ламины и LAP.
После удаления ламины из этих экстрактов с помощью иммобилизованных антител к белкам ламины формирование ядерной оболочки вокруг хроматина сперматозоидов еще возможно. Однако ядра маленькие и хрупкие, и репликации ДНК в них не происходит. Эти результаты свидетельствуют о том, что ламина может играть важную роль в организации хроматина и репликации ДНК.
Повышенная хрупкость клеток связана с мутациями, затрагивающими белки промежуточных филаментов. Мутации в ламинах и LAP обусловливают развитие ряда генетических заболеваний, особенно поражающих мышечную систему. Эти заболевания называются ламинопатиями. По-видимому, изменения ламины придают ядру хрупкость и большую чувствительность к повреждениям. Особенно склонны к повреждениям мышечные клетки, поскольку при сокращении мышц ядра их клеток испытывают большую механическую нагрузку, чем ядра клеток других тканей.
Ядерная ламина сцеплена с внутренней ядерной мембраной двумя типами связей. Когда у сперматозоидов Xenopus laevis удаляют плазматическую мембрану и инкубируют их с экстрактом яйцеклеток,
происходит деконденсация хроматина и вокруг него образуется функционально-активная ядерная оболочка.
Флуоресценция обнаруживает локализацию ДНК.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Ядерная ламина; структура и функции
Ядерная ламина (ядерная пластинка) — жесткая фибриллярная белковая сеть, образованная белками-ламининами (промежуточными филаментами), подстилает ядерную мембрану, поддерживая ее. Она представляет собой фиброзный слой ядерной оболочки с поровыми комплексами. Связана с интегральными белками слоем, состоящим из переплетенных промежуточных филаментов (ламинов), образующих кариоскелет. К ламине прикреплены нити хромосомной ДНК, т.е. участвует в организации хроматина. Белки поровых комплексов структурно связаны с белками ядерной ламины, которая участвует в их организации. Таким образом, ламина играет очень важную роль в поддержании формы ядра, упорядоченной укладке хроматина, структурной организации поровых комплексов и формировании кариолеммы при делении клеток (дезинтеграция ядерной оболочки в профазе и интеграция в телофазе).
75.Хроматин. Хроматин – мелкие зернышки и глыбки, который обнаруживается в ядре клеток и хорошо воспринимает краситель (хромос), откуда и произошло его название. В химическом плане хроматин состоит из комплекса ДНК и белка (туда также входит РНК) и соответствует хромосомам, которые в интерфазном ядре представлены тонкими нитями и неразличимы как индивидуальные структуры. Хроматин – вещество хромосом. Хроматин состоит из хроматиновых фибрилл, которые могут располагаться в ядре рыхло или компактно. На этом основании различают два вида хроматина: эухроматин - рыхлый или деконденсированный (деспирализованный) хроматин, слабо окрашивается основными красителями и не виден в световой микроскоп, он доступен для транскрипции; гетерохроматин - компактный или конденсированный (спирализованный) хроматин, хорошо окрашивается этими же красителями и виден в световой микроскоп. При подготовке клетки к делению (интерфаза) в ядре происходит спирализация хроматиновых фибрилл и превращение хроматина в хромосомы. После деления в ядрах дочерних клеток происходит деспирализация хроматиновых фибрилл и хромосомы снова преобразуются в хроматин. Следовательно, хроматин и хромосомы представляют собой различные фазы одного и того же вещества. Таким образом, по морфологическим признакам ядра (по соотношению содержания эу- и гетерохроматина) можно оценить активномть процессов транскрипции (синтетическую функцию клетки).При ее повышении это соотношение изменяется в пользу эухроматина, и наоборот.При полном подавлении функции ядра (например, в поврежденных и гибнущих клетках, при ороговении эпителия) оно уменьшается в размерах, содержит только гетерохроматин и окрашивается основными красителями интенсивно и равномерно. Это явление – кариопикноз. Распределение гетерохроматина и соотношение содержания эу- и гетерохроматина характерны для клеток каждого типа, что позволяет осуществлять их идентификацию. Вместе с тем имеются общие закономерности распределения гетерохроматина в ядре: его скопления располагаются под кариолеммой, прерываясь в области пор (что обусловлено его связью с ламиной) и вокруг ядрышка. Более мелкие глыбки разбросаны по всему ядру. Тельце Бара – скопление гетерохроматина, соответствующее одной Х-хромосоме у особей женского пола, которая в интерфазе скручена и неактивна. В большинстве клеток оно лежит у кариолеммы, а в гранулоцитах крови имеет вид маленькой добавочной дольки ядра («барабанной палочки»). Выявление тельца Бара (обычно в эпителиальных клетках слизистой оболочки полости рта) используется как диагностический тест для определения генетического пола.
76.Диффузный и конденсированный хроматин (эу- и гетерохроматин).
Хроматин – мелкие зернышки и глыбки, который обнаруживается в ядре клеток и хорошо воспринимает краситель (хромос), откуда и произошло его название. В химическом плане хроматин состоит из комплекса ДНК, РНК и белка, где ДНК находится в разной степени конденчации, и соответствует хромосомам, которые в интерфазном ядре представлены тонкими нитями и неразличимы как индивидуальные структуры. Хроматин – вещество хромосом. Хроматин состоит из хроматиновых фибрилл, которые могут располагаться в ядре рыхло или компактно. На этом основании различают два вида хроматина: эухроматин - рыхлый или деконденсированный (деспирализованный) хроматин, слабо окрашивается основными красителями и не виден в световой микроскоп, он доступен для транскрипции; гетерохроматин - компактный или конденсированный (спирализованный) хроматин, хорошо окрашивается этими же красителями и виден в световой микроскоп. При подготовке клетки к делению (интерфаза) в ядре происходит спирализация хроматиновых фибрилл и превращение хроматина в хромосомы. После деления в ядрах дочерних клеток происходит деспирализация хроматиновых фибрилл и хромосомы снова преобразуются в хроматин. Следовательно, хроматин и хромосомы представляют собой различные фазы одного и того же вещества. Таким образом, по морфологическим признакам ядра (по соотношению содержания эу- и гетерохроматина) можно оценить активномть процессов транскрипции (синтетическую функцию клетки).При ее повышении это соотношение изменяется в пользу эухроматина, и наоборот.При полном подавлении функции ядра (например, в поврежденных и гибнущих клетках, при ороговении эпителия) оно уменьшается в размерах, содержит только гетерохроматин и окрашивается основными красителями интенсивно и равномерно. Это явление – кариопикноз. Распределение гетерохроматина и соотношение содержания эу- и гетерохроматина характерны для клеток каждого типа, что позволяет осуществлять их идентификацию. Вместе с тем имеются общие закономерности распределения гетерохроматина в ядре: его скопления располагаются под кариолеммой, прерываясь в области пор (что обусловлено его связью с ламиной) и вокруг ядрышка (перинуклеарно). Более мелкие глыбки разбросаны по всему ядру. Тельце Бара – скопление гетерохроматина, соответствующее одной Х-хромосоме у особей женского пола, которая в интерфазе скручена и неактивна. В большинстве клеток оно лежит у кариолеммы, а в гранулоцитах крови имеет вид маленькой добавочной дольки ядра («барабанной палочки»). Выявление тельца Бара (обычно в эпителиальных клетках слизистой оболочки полости рта) используется как диагностический тест для определения генетического пола.
Строение и функции ламинов
К типу V белков промежуточных филаментов относятся ламины, которые по ряду своих свойств существенно отличаются от белков цитоплазматических филаментов:
• Они локализуются внутри ядра
• Для них характерен альтернативный сплайсинг
• Они подвергаются модификациям, которые обеспечивают им прикрепление к мембране
• Они обладают стержнеобразными доменами с крайне протяженными спиральными участками 1В. Эту особенность они разделяют с белками промежуточных филаментов беспозвоночных, и
• Для них характерен совершенной иной способ сборки филаментов, и они никогда не образуют сополимеров с белками промежуточных филаментов, не относящимися к типу V.
Ламины образую прочную сеть филаментов на внутренней поверхности ядерной мембраны, где они взаимодействуют с набором специфических белков, включая белки, связывающиеся с ламином, и рецептор ламина В. В результате такого взаимодействия образуется и поддерживается функциональное окружение ядра клетки. У млекопитающих три гена (LMNA, LMNB1 и LMNB2) кодируют синтез шести белков: ген LMNA в результате альтернативного сплайсинга образует три иРНК, соответствующие белкам А, С1 и С2 (известные под общим названием ламины A-типа).
Ген LMNB1 кодирует ламин В, а иРНК ламина В2 в результате сплайсинга может транслироваться в В2 или В3. Ламины В-типа экспрессируются во всех клетках, начиная с раннего эмбионального периода, в то время как образование ламинов A-типа ограничено более дифференцированными клетками. Например, базальные кератиноциты кожного эпидермиса экспрессируют только ламин В2, а по мере их дифференцировки одновременно начинают экспрессироваться ламины A-типа. Экспрессия ламинов С2 и В3 и их гомологов у других позвоночных ограничивается герминативными клетками.
Белки филаментов ламины характеризуются значительными структурными отличиями от белков цитоплазматических промежуточных филаментов. Сборка их в филаменты также отличается от процесса, характерного для цитоплазматических филаментов. У ламинов спирализованный субдомен 1 содержит шесть гептад аминокислот (всего 42 остатка), которые отсутствуют в белках цитоплазматических филаментов позвоночных.
Такой разницы в размерах стержнеобразных доменов ламинов достаточно для предотвращения возможности их сополимеризации с белками других промежуточных филаментов. Длинные спирализованные субдомены 1 также обнаружены в белках промежуточных филаментов цитоплазмы у беспозвоночных, что рассматривается как веское доказательство того, что они являются достаточно «древними» в эволюционном плане.
Иммунофлуоресцентное окрашивание ламина В в фибробластах на разных стадиях клеточного цикла.
Ламин В локализуется в ядерной оболочке в интерфазе, однако в профазе, по мере его фосфорилирования, фрагментирует.
В метафазе ламин В остается в диспергированном состоянии и в телофазе реассоциирует с хроматином,
формируя новую оболочку вокруг каждого дочернего ядра, образующегося при цитокинезе.
Поэтому считается, что ламины относятся к эволюционно наиболее древним формам промежуточных филаментов. Вероятно, создание механизма защиты хрупких нитей ДНК от разрывов явилось ранним и существенным шагом на пути эволюции сложных организмов.
Как показано на рисунке ниже у клеток млекопитающих, в митозе, ядерная оболочка распадается, и для того, чтобы это могло произойти, ядерная ламина должна диссоциировать. В профазе происходит фосфорилирование в головных и хвостовых доменах ламинов. Это осуществляется под действием митотической киназы Cdk1 и приводит к разборке ламины. Хвостовые домены также содержат последовательности, которые специфически направляют ламины в ядро (сигнал ядерной локализации) и удерживают их там (мембранное заякоривание, которое отсутствует у ламинов А-типа).
В митозе ламины А- и В-типа ведут себя по-разному. Ламины В-типа во время всего митоза остаются связанными с фрагментами везикул ядерной мембраны, в то время как ламины A-типа распределяются по всей цитоплазме. В конце митоза, когда вновь формируется ядерная оболочка, ламины постепенно связываются с конденсированным хроматином. Ламины В-типа взаимодействуют с ядерной мембраной посредством посттрансляционной модификации липидов по местам расположения консервативных последовательностей на конце хвостового домена.
В ламинах A-типа этот сайт удаляется при сплайсинге РНК или при посттрансляционном протеолизе. Таким образом, в большинстве клеток лишь В-ламин может непосредственно взаимодействовать с ядерной мембраной. Однако в генеративных клетках (сперматозоиды) С2 ламин обладает дополнительным механизмом заякоривания, который обеспечивает связывание его головного домена с ядерной мембраной.
Выяснение механизмов взаимосвязи между мутациями ламинов и возникновением определенных заболеваний представляет собой трудную задачу. Мутации в ламинах A-типа связаны с разнообразными генетическими нарушениями, которые затрагивают мышечную, нервную и жировую ткань. Эти нарушения варьируют от семейной парциальной липодистрофии (потеря жировой ткани из некоторых участков тела) до различных форм прогрессирующей периферической нейродегенерации (болезнь Charcot-Marie-Tooth типа 2В1) и прогерии. Как показывают эксперименты на культурах клеток, мутации, которые нарушают функции ламинов или полностью их выключают, приводят к снижению прочности ядерной оболочки, которая становится неспособной поддерживать свою форму; у больных развиваются спонтанные грыжи и пониженная устойчивость к механическим нагрузкам.
У человека мутации в ламине В-типа не обнаружены, и, вероятно, они являются летальными, поскольку эти ламины экспрессируются в раннем эмбриогенезе во всех клетках. Однако мыши, в клетках которых экспрессируется укороченный, поврежденный вариант ламина В, характеризуются множественными дефектами развития, напоминающими многие патологии человека, связанные с мутациями в ламинах A-типа, которые вскоре после рождения оказываются летальными. Эта модель поможет прояснить механизмы развития некоторых болезней человека.
Исследование последствий нарушения функций некоторых белков оказывается чрезвычайно информативным для понимания белковых функций вообще, и, конечно, это, в частности, относится и к промежуточным филаментам. Разумеется, мы не может утверждать, что в полной мере выяснили патогенез того или иного заболевания, однако в случае кератинов, десмина, GFAP и белков нейрофиламентов существует интуитивная связь между клиническими его проявлениями и повышенной травмируемостью клеток Однако постоянно пополняющийся список разнообразных «ламинопатий» порождает вопрос, действительно ли все они связаны лишь со снижением эластичности клеток и тканей, имеющей первичную или вторичную природу.
Конечно, на уровне клетки, многие типы физиологического стресса могут переходить в механический стресс. Так, например, воздействие химических соединений может вызвать увеличение осмотического давления. Наоборот, ламинопатии могут отражать ускоренную селективное удаление из ткани поврежденных клеток с первичной тканевой мишенью, определяемой другими, пока неидентифицированными факторами. В этой области предстоит выполнить еще много исследований.
Клеточное ядро.
Форма ядра различных клеток неодинакова: встречаются клетки с округлым, овальным, бобовидным, палочковидным, многолопастным, сегментированным ядром. Чаще всего форма ядра в целом соответствует форме клетки: оно обычно сферическое в клетках округлой или кубической формы, вытянутое или эллипсоидное в призматических клетках, уплощенное в плоских. Расположение ядра варьирует в разных клетках, оно может лежать в центре (в клетках округлой, плоской, кубической или вытянутой формы), у ее базального полюса (в клетках призматической формы) или на периферии (жировые клетки). Величина ядра в среднем 5-10 мкм, и она относительно постоянна для каждого типа клеток, однако может меняться в определенных пределах, увеличиваясь при увеличении функциональной активности клетки и уменьшаясь при ее угнетении. В разных видах клеток наблюдается неодинаковое соотношение ядра и цитоплазмы. Так, например, в клетках ядерного типа - крупное ядро и узкий ободок цитоплазмы (лимфоцит), в клетках цитоплазматического типа объем цитоплазмы превосходит объем ядра (н-р, бокаловидные клетки).
Компоненты ядра.Кариолемма - или ядерная оболочка, хроматин, ядрышко и кариоплазма (ядерный матрикс или ядерный сок).
Функции ядра:
1. хранение генетической информации (в молекулах ДНК, находящихся в хромосомах)
2. реализация генетической информации, контролирующей осуществление разнообразных процессов в клетке - от синтетических до запрограммированной гибели (апоптоз)
3. воспроизведение и передачу генетической информации (при делении клетки)
Ядерная оболочка - на светооптическом уровне практически не определяется, под электронным микроскопом обнаруживается, что она состоит из двух мембран - наружной и внутренней разделенных полостью шириной 15-40 нм (перинуклеарным пространством). Наружная - покрыта рибосомами и тесно связана с гр. ЭПС. Нередко можно видеть, как наружная мембрана продолжается в канальцы гр. ЭПС. Внутренняя мембрана является местом прикрепления хромосом. В нуклеолемме имеются ядерные поры. В их состав входят поровые комплексы, в составе которых имеются: отверстие поры диаметром около 90 нм, гранулы поры и мембрана поры. Отверстие поры образуется в результате слияния наружной и внутренней мембран. Гранулы поры располагаются в 3 ряда, по 8 гранул в каждом ряду. Размеры гранул около 25 нм. От гранул к центру сходятся фибриллы, формирующие диафрагму, в середине которой лежит центральная гранула (по некоторым представлениям - это транспортируемая через пору субъединица рибосомы.
Функции комплекса ядерной поры:
1. Обеспечение регуляции избирательного транспорта веществ между цитоплазмой и ядром.
2. Активный перенос в ядро некоторых белков, имеющих особую маркировку и распознаваемой рецепторами в комплексе поры.
3. Перенос в цитоплазму субъединиц рибосом.
Чем больше пор в нуклеолемме, тем активнее ядро, если активность снижена, то количество пор уменьшается, если синтетическая активность ядра близка к нулю, то поры в ядре отсутствуют (н-р, в кариолемме ядра сперматозоида).
Хроматин занимает основную часть объёма ядра. Он представлен тёмными (электроноплотными) глыбками - т.н. гетерохроматином (функционально неактивныеотделы и целые хромосомы, которые конденсированы, образуя глыбки) и светлыми (электронопрозрачными) областями -эухроматиномэто функционально активные, участвующие в транскрипции части хромосом, которые находятся в деконденсированном (диффузном) состоянии. Причём, глыбки гетерохроматина находятся, главным образом, на периферии ядра
и прилежат к ядерной оболочке. При изменении состояния клетки или в процессе дифференцировки возможен переход части гетерохроматина в эухроматин и обратно.Таким образом, чем больше в ядре доля гетерохроматина, тем ниже функциональная активность ядра, т.е. тем меньше скорость синтеза РНК. Так, в ядре нервной клетки гетерохроматина очень мало - ядро и клетка в целом функционально очень активны. Напротив, в лимфоците наблюдается преобладание гетерохроматина. Это вполне коррелирует с очень малым объёмом цитоплазмы, которая к тому же бедна органеллами. Данная клетка циркулирует в крови, и процессы синтеза РНК и белков идут в ней с небольшой скоростью. Хроматин дает положительную реакцию на ДНК (реакция Фельгена) - ДНК окрашивается в вишневый цвет, а все прочие структуры в зеленый, также по методу Браше хроматин окрашивается метиловым зеленым в соответствующий цвет красителя. Весь хроматин в целом - это совокупность 46 хромосом. Каждая из них представляет собой нуклеопротеидный комплекс - двуцепочечную молекулу ДНК, которая определённым образом связана с ядерными белками. Содержание белков в хромосоме по массе в 1,3-1,7 раза больше, чем ДНК. Кроме того, в хромосоме обнаруживается и РНК, являющаяся продуктом транскрипции. Тарнскрипция сопровождается экспрессией рибосомных и нерибосомных генов. Ген - единица наследственной информации, состоящий из нуклеотидов.
Экспрессия нерибосомных генов:
1. Сопровождается синтезом гигантской молекулы гетерогенной и-РНК при участии катализатора ДНК-полимеразы
2. Гигантская и-РНК подвергается сплайсингу, в результате длина гигантской и-РНК укорачивается в 10-30 раз и она превращается в и-РНК.
3. и-РНК взаимодействует с белками и образует РНК-азную гранулу (интерхроматиновая гранула диаметром 30 нм).
4. Перемещаясь на перпиферию ядра интерхроматиновая гранула перезревает в перихроматиновую гранулу диаметром 45 нм.
5. Перихроматиновая гранула покидает ядро в форме информосом.
Одним из компонентов гетерохроматина может быть т.н. половой хроматин.
У мужчин в наборе хромосом каждой клетки содержатся, как известно, по одной Х- и Y-половой хромосоме. Обе они находятся в деконденсированном состоянии, т.е. входят во фракцию эухроматина. У женщин в клетках содержатся по две Х-хромосомы. Одна из них деконденсирована. Вторая же Х-хромосома всегда находится в конденсированном состоянии, образуя в ядре компактное тельце - половой хроматин. Для обнаружения полового хроматина обычно исследуют мазок крови. В нейтрофильных лейкоцитах женщин половой хроматин выявляется в виде барабанной палочки, находящейся в одном из сегментов ядра. По этому признаку в судебной медицине отличают кровь женщин от крови мужчин. В деконденсированном состоянии длина одной молекулы ДНК равна в среднем около 5 см, а общая длина молекулы ДНК всех хромосом в ядре более 2м. В этой связи очевидна необходимость компактной упаковки молекул ДНК.
Выделяют 4 уровня компактизации ДНК:
1. Нуклеосомный (длина уменьшается в 6-7 раз). Нуклеосома - это белковая частица, состоящая из основных белков - гистонов. Основа каждойнуклеосомы- глобула из 8 молекул гистонов (октамер). Двуцепочечная молекула ДНК последовательно "намотана" на огромное количество таких глобул, делая вокруг каждой из них почти по 2 оборота (1,75 раз). В участках между глобулами с ДНК связано ещё по 1 молекуле гистона. В итоге, совокупность нуклеосом выглядит как цепь бусин, а деконденсированный хроматин имеет гранулярную структуру.
2. Нуклеомерный (компактизация в 40 раз). Формируется суперспираль. Ее витки образуют нуклеосомы обвитые ДНК. Каждый виток суперспирали образован 6-ю нуклеосомами и называется нуклеомерой, ее диаметр 25-30 нм. Суперспираль - это элементарная нить эухроматина. Гетерохроматин и полностью конденсированные хромосомы тоже имеют нуклеосомную организацию. Однако здесь добавляются и следующие уровни укладки хромосомы, что приводит к резкому сокращению её длины.
3. Хромомерный (компактизация в 680 раз). Негистоновые белки сшивают
суперспираль в боковые петельные домены, с образованием хромомеры.
4. Хромонемный - связывают со сближением хромомеров, боковые петли переплетаются и образуют кластеры, которые формируют хромонему. Их диаметр от 300 нм. Кластер рассматривают, как компонент гетерохроматина, видимый в световой микроскоп, как глыбка.
Совокупность числа, размеров и особенностей строения хромосом называется кариотипом. Оценка кариотипа производится путем изучения хромосом в метафазной пластинке. Для кариотипирования получают культуру клеток, в которую вводят колхицин, блокирующий формирование веретена. Из таких клеток извлекают хромосомы, которые далее окрашивают и идентифицируют. Нормальный кариотип человека представлен 46 хромосомами - 22 пары аутосом и двумя половыми хромосомами. Кариотипорование позволяет диагносцировать ряд заболеваний, связанных с хромосомными аномалиями, в частности синдром Дауна - трисомия 21 хромосомы.
Ядрышко - это место образования рибосом в клетке. Оно образовано специализированными участками хромосом, которые называются ядрышковыми организаторами. У человека такие участки имеются в пяти хромосомах - 13, 14, 15, 21 и 22, где находятся многочисленные копии генов, кодирующих рибосомальные РНК. Ядрышко выявляется в интерфазном ядре на светооптическом уровне как мелкая плотная гранула, интенсивно окрашивающаяся основными красителями. При окраске по методу Браше дает + реакцию на РНК - окрашивается пиронином в розовый цвет. Оно располагается в центре ядра или эксцентрично. Размеры и число ядрышек увеличиваются при повышении функциональной активности клетки. Особенно крупные ядрышки характерны для эмбриональных и активно синтезирующих белки клеток, а также клеток быстро растущих злокачественных опухолей. Под электронным микроскопом в ядрышке обнаруживают 3 компонента.
1. центральный фибриллярный светлый компонент - окрашивается бледно, где находится ДНК ядрышкового организатора, содержащей информацию о р-РНК.
2. периферический фибриллярный компонент - в виде тонких нитей и представляет собой транскрипты р-РНК.
3 гранулярный компонент - субъединицы рибосом.
Кариоплазма - жидкий компонент ядра, в котором располагаются хроматин и ядрышко. Содержит воду и ряд растворенных и взвешенных в ней веществ: РНК, гликопротеины, ионы, ферменты, метаболиты. Некоторые авторы разделяют понятие кариоплазмы и ядерного матрикса, к последнему помимо кариоплазмы относят также и кариоскелет, состоящий из ядерной ламины и фибриллярной сети, пронизывающей ядро. Ламина это пластинка белковой природы, которая связана с внутренней мембраной. К ядерной ламине и внутриядерной фибриллярной сети крепятся хромосомы, а также
разнообразные белковые комплексы с ферментативной или регуляторной функцией.
Клеточный цикл -это время существования клетки от одного деления до другого, или от деления до гибели. Клеточный цикл включает собственно митотическое деление и интерфазу - промежуток между делениями. Интерфаза значительно более длительна, чем митоз (обычно занимает не менее 90% клеточного цикла) и подразделяется на 3 периода: пресинтетический или постмитотический (G1), синтетический (S) и постсинтетический или премитотический (G2).
G1 - наступает сразу после митоза и характеризуется активным ростом клетки и синтезом белка и РНК, благодаря чему клетка достигает нормальных размеров и восстанавливает необходимый набор органелл (продолжительность от неск часов до неск дней)
G2 - В течение этого периода клетка осуществляет непосредственную подготовку к делению. Происходит созревание центриолей, запасается энергия, синтезируется РНК и белки (в частности тубулины, необходимые для образования веретена деления). Продолжительность 2-4 часа. Некоторые клетки могут выходить из клеточного цикла, это обозначается буквой G0. Клетка, вошедшая в этот период, утрачивает способность к митозу. В том случае, если клетка временно утрачивает способность к делению, она подвергается начальной дифференцировке. При этом дифференцированная клетка специализируется для выполнения определенной функции, после чего она способна вновь возвратиться в клеточный цикл. Н-р, при повреждении печени гепатоциты, подвергшиеся начальной дифференцировке, возвращаются в клеточный цикл, и за счет их деления происходит быстрое восстановление ткани. Те клетки, которые окончательно утрачивают способность к делению, не могут возвратиться в клеточный цикл и погибают. Н-р, гранулоциты крови, подвергшиеся дифференцировке, функционируют в течение 8 суток, а затем погибают. Также высокоспециализированные клетки - кардиомиоциты или нервные клетки не способны делиться.
Митоз - непрямое деление. В процессе, которого происходит равномерное распределение хромосомного материала между дочерними клетками. Выделяют 4 фазы, общая продолжительность которых 2 часа. Профаза - 30-60 мин, метафаза 10-20 мин, анафаза - 2-3 мин, телофаза - 30-40 мин.
Мейоз - это такое деление, при котором в дочерних клетках оказывается половинный (гаплоидный) набор хромосом. Такое деление имеет место при образовании половых клеток. Особенности мейоза: состоит из 2-х делений, второе деление без S-периода в интерфазе, 80% времени занимает профаза первого деления. В ней выделяют периоды: 1. Лептотена - хромосомы спирализуются и приобретают вид тонких нитей. 2. Зиготена - гомологичные хромосомы конъюгируют друг с другом.Пахитена - пары хромосом ещё больше спирализуются и, утолщаясь, укорачиваются. Хромосомы обмениваются гомологичными участками (кроссинговер).Вместе с тем активируются синтезы РНК и белка. Благодаря этому, сильно увеличивается объём ядра и клетки. Диплотена - гомологичные хромосомы начинают расходиться. Но между ними сохраняются хиазмы - перекрёсты в местах происшедшего кроссинговера. Из-за ещё большей спирализации хромосомы утолщаются и подразделяются на хроматиды. Поэтому каждая пара гомологичных хромосом выглядит как тетрада. Диакинез - хиазмы исчезают из-за ещё большего расхождения гомологичных хромосом.
Амитоз - этот тип деления характеризуется тем, что хромосомный материал ядра материнской клетки может распределяться неравномерно между дочерними клетками. Такой тип деления считается ненормальным.
Полиплоидия - это процесс увеличения количества хромосом в ядре клетки. В результате образуются полиплоидные клетки. Это может происходить в результате блокирования одной из фаз митоза, либо нарушения цитотомии во время телофазы. Н-р, гепатоциты, мегакариоциты красного костного мозга, гландулоциты ацинусов слюнных желез.
Эндорепродукция - это последовательное многократное удвоение ДНК, в результате увеличивается набор хромосом, при этом хромосомы связаны тонкими нитями, эти структуры называются политенами. Характерными для клеток плаценты.
Строение и функции ядерной оболочки
• Ядро окружено оболочкой, состоящей из двух мембран
• Наружная ядерная мембрана является продолжением мембран ЭПР, а перинуклеарное пространство (просвет) переходит в просвет ЭПР
• В ядерной оболочке присутствуют многочисленные ЯПК, которые представляют собой единственные каналы обмена молекулами и макромолекулами между ядром и цитоплазмой
Ядро окружено оболочкой, состоящей из двух концентрически расположенных наружной и внутренней ядерных мембран. Каждая мембрана содержит определенный набор белков и сплошной двойной слой фосфолипидов. За исключением некоторых одноклеточных эукариот, внутренняя ядерная мембрана поддерживается сетью филаментов, закрепленных в сетчатой структуре. Эта сеть филаментов называется ядерная ламина.
Наружная ядерная мембрана переходит в мембраны ЭПР и, так же как большая часть его мембран, покрыта рибосомами, принимающими участие в синтезе белка. На рисунке ниже показана связь наружной мембраны с ЭПР.
Пространство между наружной и внутренней ядерными мембранами представляет собой перинуклеарное пространство (ПП). Так же как наружная мембрана связана с мембраной ЭПР, ПП ядерной оболочки контактирует с внутренним пространством ЭПР. Толщина каждой из двух мембран составляет 7-8 нм (нм), а ширина ПП ядерной оболочки — 20-40 нм.
При исследовании препаратов ядерной оболочки в электронном микроскопе, наиболее заметной особенностью структуры являются ЯПК (ядерные поровые комплексы), которые служат каналами транспортировки большинства молекул между ядром и цитоплазмой. Оболочка ядер большинства клеток содержит около 10-20 ЯПК на квадратный микрон поверхности. Так, клетки дрожжей содержат 150-250 ЯПК, а соматические клетки млекопитающих 2000-4000.
Однако некоторые клетки обладают гораздо большей плотностью пор, вероятно, потому, что для них характерна высокая интенсивность процессов транскрипции и трансляции, что предполагает транспорт большого количества макромолекул в ядро и из него. Например, поверхность ядра ооцитов амфибий почти полностью покрыта ЯПК.
Каким образом могла возникнуть двойная ядерная мембрана? В эукариотической клетке, митохондрии и хлоропласта также имеют двойную мембрану. Согласно гипотезе эндосимбиоза, эти органеллы образовались в ходе эволюции, когда одни клетки захватили других в процессе эндоцитоза. Затем поглощенные клетки оказались окруженными двумя мембранами: своей и мембраной клетки-хозяина. Оказалось, что некоторые из поглощенных клеток проявляют метаболическую активность, например, в отличие от клеток хозяина, способны осуществлять фотосинтез.
Наиболее убедительное доказательство в пользу эндосимбиотического происхождения митохондрий и хлоропластов заключается в том, что рибосомы обеих органелл больше напоминают рибосомы современных прокариот, и в меньшей степени эти же микроструктуры цитоплазмы эукариотической клетки. Гораздо менее ясным представляется происхождение ядра. Однако существование двойной ядерной мембраны, подобно мембране митохондрий и хлоропластов, позволяет предполагать, что захваченная прокариотическая клетка превратилась в ядро, содержащее всю клеточную ДНК.
Ядерная оболочка соединена с эндоплазматическим ретикулумом (ЭПР). Поверхность ядерной мембраны ооцита Xenopus laevis покрыта комплексами ядерных пор. Ядро могло образоваться в результате эндосимбиоза, процесса,
при котором одна прокариотическая клетка захватывает другую клетку; затем захваченная клетка становится примитивным ядром.
Читайте также: