Строение нуклеоида и цитоплазмы бактерий
Добавил пользователь Евгений Кузнецов Обновлено: 27.01.2026
Структурные компоненты бактериальной клетки делят на 2 вида:
- основные структуры (клеточная стенка, цитоплазматическая мембрана с ее производными, цитоплазма с рибосомами и различными включениями, нуклеоид);
- временные структуры (капсула, слизистый чехол, жгутики, ворсинки, эндоспоры, образующиеся лишь на определенных этапах жизненного цикла бактерий).
Основные структуры.
Клеточная стенка находится с внешней стороны от цитоплазматической мембраны. Цитоплазматическая мембрана не входит в состав клеточной стенки. Функции клеточной стенки:
- защита бактерий от осмотического шока и других повреждающих факторов;
- определение формы бактерий;
- участие в метаболизме бактерий.
Клеточная стенка пронизана порами, через которые происходит транспорт экзотоксинов бактерий. Толщина клеточной стенки составляет 10–100 нм. Основной компонент клеточной стенки бактерий - пептидогликан или муреин, состоящий из чередующихся остатков N-ацетил-N-глюкозамина и N-ацетилмурамовой кислоты, соединенных гликозидными связями.
В 1884 году Х. Грам предложил метод окраски бактерий с помощью генцианвиолета, йода, этилового спирта и фуксина. Все бактерии в зависимости от окраски по Граму подразделяют на 2 группы: грамположительные и грамотрицательные бактерии. Клеточная стенка грамположительных бактерий плотно прилегает к цитоплазматической мембране, ее толщина составляет 20-100 нм. В ней имеются тейхоевые кислоты (полимеры глицерина или рибита), а также в небольших количествах полисахариды, белки и липиды. Клеточная стенка грамотрицательных бактерий многослойна, ее толщина составляет 14-17 нм. Внутренний слой (пептидогликан) образует тонкую непрерывную сетку. Внешний слой состоит из фосфолипидов, липопротеина и белков. Белки наружной мембраны прочно связаны с пептидогликановым слоем.
Различное содержание пептидогликана в клеточной стенке обусловливает различную окраску бактерий. У грамотрицательных бактерий содержание пептидогликана составляет 1-10%, а у грамположительных – от 50 до 90%. Грамположительные бактерии окрашиваются в синий (фиолетовый) цвет, а грамотрицательные бактерии – в красный цвет.
В некоторых условиях бактерии лишаются способности полностью или частично синтезировать компоненты клеточной стенки, в результате чего образуются протопласты, сферопласты и L-формы бактерий. Сферопласты – это бактерии с частично разрушенной клеточной стенкой. Они наблюдаются у грамотрицательных бактерий. Протопласты - это формы, полностью лишенные клеточной стенки. Они образуются грамположительными бактериями. L-формы бактерий - это мутанты бактерий, частично или полностью утратившие способность синтезировать пептидогликан клеточной стенки (бактерии с дефектной клеточной стенкой). Свое название они получили от названия института Листера в Англии, где были открыты в 1935 году.
ЦПМ бактерий выполняет следующие функции:
- барьерная функция (молекулярное “сито”);
- избирательный перенос различных органических и неорганических молекул и ионов с помощью специальных переносчиков – транслоказ или пермеаз;
- репликация и последующее разделение хромосомы.
В процессе роста клетки цитоплазматическая мембрана образует многочисленные впячивания (инвагинаты), получившие название мезосом.
Цитоплазма - это содержимое бактериальной клетки, ограниченное цитоплазматической мембраной. Она состоит из цитозоля и структурных элементов.
Цитозоль - гомогенная фракция, включающая растворимые компоненты РНК, ферменты, продукты метаболизма.
Структурные элементы - это рибосомы, внутрицитоплазматические мембраны, включения и нуклеоид.
Рибосомы - органоиды, осуществляющие биосинтез белка. Они состоят из белка и РНК. Представляют собой гранулы диаметром 15-20 нм. Одна бактериальная клетка содержит от 5000 до 50000 рибосом. Рибосомы являются местом синтеза белка.
В цитоплазме прокариотов обнаруживаются различные включения, представляющие запасные вещества клетки. Из полисахаридов в клетках откладываются гликоген, крахмал и крахмалоподобное вещество - гранулеза. Полифосфаты содержатся в гранулах, называемых волютиновыми, или метахроматиновыми, зернами.
Нуклеоид является ядром у прокариотов. Он состоит из одной замкнутой в кольцо двуспиральной нити ДНК, которую рассматривают как бактериальную хромосому. У нуклеоида отсутствует ядерная оболочка.
Кроме нуклеоида в бактериальной клетке обнаружены внехромосомные генетические элементы – плазмиды, которые представляют собой небольшие кольцевые молекулы ДНК, способные к автономной репликации. Роль плазмид состоит в том, что они кодируют дополнительные признаки, дающие клетке преимущества в определенных условиях существования. Наиболее распространены плазмиды, детерминирующие признаки антибиотикорезистентности бактерий (R-плазмиды), синтез энтеротоксинов (Ent-плазмиды) или гемолизинов (Hly-плазмиды).
К временным структурам относятся капсула, жгутики, пили, эндоспоры бактерий.
Капсула - это слизистый слой над клеточной стенкой бактерии. Вещество капсул состоит из нитей полисахаридов. Капсула синтезируется на наружной поверхности цитоплазматической мембраны и выделяется на поверхность клеточной стенки в специфических участках.
- место локализации капсульных антигенов, определяющих вирулентность, антигенную специфичность и иммуногенность бактерий;
- защита клеток от механических повреждений, высыхания, токсических веществ, заражения фагами, действия защитных факторов макроорганизма;
- способность прикрепления клеток к субстрату.
Жгутики – это органы движения бактерий. Жгутики не являются жизненно важными структурами, поэтому могут присутствовать у бактерий или отсутствовать в зависимости от условий выращивания. Количество жгутиков и места их расположения у разных бактерий неодинаково. В зависимости от этого выделяют следующие группы жгутиковых бактерий:
- монотрихи – бактерии с одним полярно расположенным жгутиком;
- амфитрихи – бактерии с двумя полярно расположенными жгутиками или имеющие по пучку жгутиков на обоих концах;
- лофотрихи – бактерии, имеющие пучок жгутиков на одном конце клетки;
- перитрихи – бактерии с множеством жгутиков, расположенных по бокам клетки или на всей ее поверхности.
Химический состав жгутиков представлен белком флагеллином.
К поверхностным структурам бактериальной клетки относятся также ворсинки и пили. Эти структуры участвуют в адсорбции клеток на субстрате (ворсинки, пили общего типа) и в процессах переноса генетического материала (половые пили). Они образованы специфическим гидрофобным белком пилином.
У некоторых бактерий в определенных условиях образуются покоящиеся формы, которые обеспечивают переживание клеток в течение длительного времени в неблагоприятных условиях - эндоспоры. Они устойчивы к неблагоприятным факторам внешней среды.
Строение нуклеоида и цитоплазмы бактерий
Структура бактериальной клетки
Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и ядерного аппарата, называемого нуклеоидом. Имеются другие структуры: мезосома, хроматофоры, тилакоиды, вакуоли, включения полисахаридов, жировые капельки, капсула (микрокапсула, слизь), жгутики, пили. Некоторые бактерии способны образовывать споры.
Структуру и морфологию бактерий изучают с помощью различных методов микроскопии: световой, фазово-контрастной, интерференционной, темнопольной, люминесцентной и электронной.
Обозначения:
Клеточная стенка
В клеточной стенки грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков. Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40—90% массы клеточной стенки. С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. teichos — стенка).
В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана. На ультратонких срезах бактерий наружная мембрана имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, которую называют цитоплазматической. Основным компонентом этих мембран является бимолекулярный (двойной) слой липидов. Внутренний слой наружной мембраны представлен фосфолипидами, а в наружном слое расположен липополисахарид (ЛПС). Липополисахарид наружной мембраны состоит из трех фрагментов: липида А - консервативной структуры, практически одинаковой у грамотрицательных бактерий; ядра, или стержневой, коровой части (лат. core — ядро), относительно консервативной олигосахаридной структуры (наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота); высоковариабельнои О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями (О-антиген). Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы.
При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима,
пенициллина, защитных факторов организма образуются клетки с измененной (часто шаровидной) формой: протопласты — бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. Бактерии сферо- или протопластного типа, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами.
Они представляют собой осмотически чувствительные, шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, «возвращаясь» в исходную бактериальную клетку.
Между наружной и цитоплазматической мембранами находится периплазматическое пространство, или периплазма, содержащая ферменты (протеазы, липазы, фосфатазы, нуклеазы, бета-лактомазы) и компоненты транспортных систем.
Цитоплазматическая мембрана
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану (2 темных слоя толщиной по 2,5 нм разделены светлым - промежуточным). По структуре она похожа на плазмалемму клеток животных и состоит из двойного слоя фосфолипидов с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты — впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами.
Цитоплазма
Цитоплазма состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул — рибосом, ответственных за синтез (трансляцию) белков. Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, в отличие от 80S-рибосом, характерных для эукариотических клеток. Рибосомные РНК (рРНК) - консервативные элементы бактерий («молекулярные часы» эволюции). 16S рРНК входит в состав малой субъединицы рибосом, а 23S рРНК - в состав большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики, позволяя оценить степень родства организмов.
В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, бета-оксимасляной кислоты и полифосфатов (волютин). Они являются запасными веществами для питания и энергетических потребностей бактерий. Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки.
Нуклеоид
Нуклеоид — эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитевой ДНК, замкнутой в кольцо и плотно уложенной наподобие клубка. Ядро бактерий, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Обычно в бактериальной клетке содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК.
Кроме нуклеоида, представленного одной хромосомой, в бактериальной клетке имеются внехромосомные факторы наследственности - плазмиды, представляющие собой ковалентно замкнутые кольца ДНК.
Капсула, микрокапсула, слизь
Капсула - слизистая структура толщиной более 0,2мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски мазка (например, по Бурри-Гинсу), создающих негативное контрастирование веществ капсулы: тушь создает темный фон вокруг капсулы. Капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов, например, у сибиреязвенной бациллы она состоит из полимеров D-глутаминовой кислоты. Капсула гидрофильна, препятствует фагоцитозу бактерий. Капсула антигенна: антитела против капсулы вызывают ее увеличение (реакция набухания капсулы).
Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее 0,2мкм, выявляемое лишь при электронной микроскопии. От капсулы следует отличать слиэь - мукоидные экзополисахариды, не имеющие четких границ. Слизь растворима в воде.
Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам), их еще называют гликокаликсом. Кроме синтеза
экзополисахаридов бактериями, существует и другой механизм их образования: путем действия внеклеточных ферментов бактерий на дисахариды. В результате этого образуются декстраны и леваны.
Жгутики
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из 3 частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (1 пара дисков - у грамположительных и 2 пары дисков - у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем-мотором, вращающим жгутик. Жгутики состоят из белка - флагеллина (от flagellum - жгутик); является Н-антигеном. Субъединицы флагеллина закручены в виде спирали.
Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм х 0, 3-10мкм) , чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, обладающего антигенной активностью. Различают пили, ответственные за адгезию, то есть за прикрепление бактерий к поражаемой клетке, а также пили, ответственные за питание, водносолевой обмен и половые (F-пили), или конъюгационные пили. Пили многочисленны - несколько сотен на клетку. Однако, половых пилей обычно бывает 1-3 на клетку: они образуются так называемыми "мужскими" клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Col-плазмиды). Отличительной особенностью половых пилей является взаимодействие с особыми "мужскими" сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях.
Споры
Споры - своебразная форма покоящихся фирмикутных бактерий, т.е. бактерий
с грамположительным типом строения клеточной стенки. Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.. Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Спорообразующие бактерии рода Bacillus имеют споры, не превышающие диаметр клетки. Бактерии, у которых размер споры превышает диаметр клетки, называются клостридиями, например, бактерии рода Clostridium (лат. Clostridium - веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля-Нильсена в красный, а вегетативная клетка в синий цвет.
Форма спор может быть овальной, шаровидной; расположение в клетке -терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулиэма, газовой гангрены) и центральное (у сибиреязвенной бациллы). Спора долго сохраняется из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизмов. В благоприятных условиях споры прорастают, проходя три последовательные стадии: активация, инициация, прорастание.
Строение нуклеоида и цитоплазмы бактерий
• Нуклеоид бактерий выглядит как диффузная масса ДНК, однако для него характерна высокая упорядоченность и неслучайное расположение генов
• У бактерий нет нуклеосом, однако организации ДНК способствуют различные белки, связанные с нуклеоидом
• Подобно тому как это имеет место для ядра и цитоплазмы эукариотической клетки, у бактерий транскрипция происходит по всей массе нуклеоида, трансляция — на его периферической зоне
• Важную роль в организации нуклеоида играет РНК полимераза
Фундаментальное отличие клеток прокариот от клеток эукариот заключается в отсутствии у них ядерной оболочки. Присутствие ядерной мембраны у эукариот обеспечивает существование компартментов, которые разделяют процессы транскрипции и трансляции. У прокариот эти процессы не разделены мембраной, и мРНК может транслироваться во время транскрипции. Одновременное протекание этих процессов имеет важные последствия для регуляции активности некоторых генов.
Как показано на рисунке ниже, хромосомальная ДНК бактерий имеет вид аморфной массы, нуклеоида, занимающего большую часть объема в центре цитоплазмы. Нуклеоид состоит из хромосомальной ДНК и связанных с ней белков. Бактерии не содержат нуклеосом, которые участвуют в упаковке ДНК хромосом эукариотов и архей. Однако бактериальная ДНК компактна и упакована с участием многочисленных белков, ассоциированных с нуклеоидом, которые перчислены на рисунке ниже.
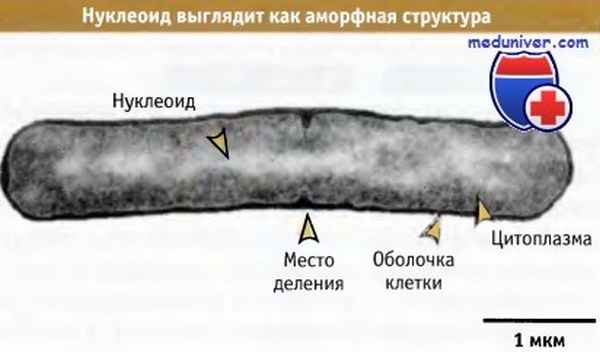
Электронная микрофотография, демонстрирующая,
что нуклеоид представляет собой диффузную массу, находящуюся внутри клетки бактерии.
К числу наиболее важных из этих белков относятся топоизомеразы. Они контролируют суперспирализацию ДНК, которая играет важную роль в ее компактизации, и обеспечивают протекание таких процессов, как репликация и транскрипция, для которых требуется раскручивание молекулы ДНК. Белки семейства SMC, поддерживающие структурную организацию хромосом, также участвуют в организации нуклеоида. Об этом свидетельствует фенотип соответствующих мутантов, однако конкретный механизм их участия остается неясным.
В клетках эукариот белки, близкие к SMC, участвуют в скреплении хромосом между собой и их конденсации в митозе и мейозе. Эти белки различной природы, связанные с нуклеоидом, участвуют в поддержании необходимого уровня его суперспирализации и компактизации. Однако предстоит еще выяснить, каким образом достигается и поддерживается такое состояние гомеостаза нуклеоида.
Хотя нуклеоид обладает аморфной структурой, отдельные гены располагаются в нем упорядоченно. Положение генов в нуклеоиде отражает их относительное расположение на карте хромосомы. По счастью, первое подтверждение этого было получено при исследовании свойств мутантов бактерий В. subtilis, дефектных по гену spoIIIE. Мутант этого организма не способен правильно сегрегировать хромосому при асимметрическом делении, которое сопровождает ранние стадии образования споры. Вместо этого септа деления замыкается вокруг одной копии хромосомы. У этого мутанта определенные гены почти всегда попадают в небольшой компартмент, поблизости от полюса, в то время как другие из него всегда исключаются.
Это наблюдение позволяет предполагать, что до деления хромосома всегда находится в определенном месте и в определенной ориентации.
Прямые данные были получены в исследованиях с использованием гибридизации in situ и флуоресцентной метки (FISH). Этот метод позволяет непосредственно отслеживать положение в клетке определенных генов. Однако при его использовании, перед гибридизацией зонда с ДНК, необходима фиксация препаратов и проведение других жестких воздействий. Еще один подход заключается в использовании конъюгата зеленого флуоресцирующего белка с белком LacI, связывающимся с ДНК. Этот конъюгат может присоединяться к сайтам связывания, находящимся в разных местах клетки. На основании всех этих экспериментов было показано, что гены не диффундируют по бактериальной клетке свободно, а локализованы в определенных, строго ограниченных местах.
Вообще говоря, область хромосомы, содержащая oriC, находится на одном конце нуклеоида, а область, содержащая terC, — на противоположном. Гены, которые на генетической карте расположены между двумя этими точками, распределены по нуклеоиду более или менее пропорционально.
У бактерий в аппарате транскрипции используется одна каталитическая основная РНК-полимераза, состоящая из двух а-, одной b- и одной b-субъединиц. Специфичность промотора определяется на начальном уровне различными сигма (а) факторами, которые также необходимы для инициации транскрипции, однако после этого отщепляются от кора. Регуляция транскрипции осуществляется большим набором дополнительных регуляторов, которые обычно связываются с ДНК вблизи от промотора, с тем чтобы активировать или подавлять инициацию транскрипции. Другие факторы регуляции действуют на уровне терминации (прекращения) транскрипции или изменения стабильности мРНК.
Большая часть молекул основной РНК-полимеразы находится в нуклеоиде в центре клетки. Поэтому, вероятно, здесь в основном происходит транскрипция. Напротив, рибосомы и различные белки, принимающие участие в трансляции, сосредоточены по периферии клетки. Таким образом, даже при отсутствии ядерной оболочки, в бактериальной клетке транскрипция и трансляция пространственно разобщены, подобно тому как это имеет место в клетке эукариот. Однако существуют различные данные в пользу того, что иногда у бактерий транскрипция и трансляция тесно сопряжены друг с другом.
Эти данные не противоречат имеющимся результатам, которые свидетельствуют о том, что РНК-полимеразы и рибосомы локализованы в разных местах клетки. Возможно, что оба процесса происходят на границе центральной, или сердцевинной, и периферийной областях клетки. Пока мы мало знаем об организации центральной, или сердцевинной, и периферийной областей нуклеоида, так же как и о деталях общей организации этой структуры.
Белки, участвующие в организации нуклеоида Escherichia coli.
У большинства других бактерий вместо белков MukB, MukE и MukF присутствуют белки SMC (белки, поддерживающие структуру хромосом),
а также связанные с ними факторы, родственные когезину и конденсинам эукариот. Сегрегация хромосом после образования полярной септы при наступлении споруляции.
В холе споруляции В. subtilis клетки делятся асимметрично, образуя материнскую клетку и небольшую преспору.
Каждая клетка получает одну копию хромосомы. Сегрегация хромосом с образованием преспоры представляет собой двухэтапный процесс.
Вначале полярная разделительная септа замыкается вокруг хромосомы,
а затем белок SpoIIIE активно транспортирует оставшиеся 2/3 хромосомы в преспоровый компартмент.
У мутантов по гену spoIIIE только 1/3 хромосомы сегрегирует в преспору.
Анализ ДНК, захваченной в небольшой компартмент клеток мутанта по гену spoIIIE, показывает, что всегда захватывается специфический участок ДНК.
Это указывает на то, что до деления хромосома должна быть строго ориентирована и упорядочена.
На фотографиях, полученных во флуоресцентном микроскопе, представлены клетки спорулирующих spoIIIE-мутантов и клетки дикого типа, окрашенные на ДНК. Несмотря на отсутствие ядерной оболочки, аппараты транскрипции и трансляции локализуются в отдельных частях бактериальной клетки.
Представлены делящиеся клетки В. subtilis.
Они экспрессируют конъюгаты белка рибосомальной субъединицы RpsB с зеленым флуоресцирующим белком (GFP)
и субъединицы РНК-полимеразы RpoC с GFP-UV, обладающие зеленой и красной флуоресценцией соответственно.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Нуклеоид
Нуклеоид – ядро клетки или генетический материал прокариот. В нем сосредоточен основной объем генетической информации бактериальной клетки (бактерии) [2] [1] .
Факт присутствия в нуклеоиде ДНК впервые был доказан Ж. Кейрнсом при помощи метода радиоавтографии бактерии кишечной палочки (Escherichia coli). Радиоавтографы развернутой молекулы бактериальной ДНК данного вида доказывают, что для бактерий Escherichia coli ДНК имеет форму нити, замкнутой в кольцо. Эта молекула (хромосома) включает в семя несколько тысяч генов, расположенных линейно [2] .
Внешний вид нуклеоида
Полностью «уложенный» нуклеоид представляет собой компактное образование. В зависимости от метода микроскоприрования и фиксации нуклеоид может выглядеть различным образом:
- При фиксировании парами осмия и применении световой микроскопии нуклеоид имеет вид боковидного тела с хорошо очерченными контурами. Он занимает центральную часть клетки и у бактерий кишечной группы имеет длину около 1 мкм [2] .
- В клетке живой бактерии в фазово-контрастном микроскопе нуклеоид так же имеет вид овального тельца светлого на фоне темной цитоплазмы[2] .
- При микроскопировании ультратонких срезов бактерий в электронном микроскопе нуклеоид похож на клубок мотков толстой веревки, состоящей из множества нитей [2] .
- При микроскопировании утратонких срезов замороженных бактерий, нуклеоид просматривается в виде кораллоподобной структуры с плотной сердцевиной и тонкими рожками-выступами. Эти выступы, как ветви, пронизывают цитоплазму, образуя нечто вроде ореола вокруг сердцевины [2] .
Нуклеоид является дифференцированной структурой. В зависимости от стадии развития бактериальной клетки он может быть прерывистым (дискретным) и состоять из отдельных фрагментов. Такую форму нуклеоид обычно принимает в связи с тем, что деление бактериальной клетки наблюдается после завершения репликации молекулы ДНК и формирования дочерних хромосом [1] .
Установлено, что бактериальные клетки большинства видов бактерий содержат только одну хромосому (большинство бактерий – гаплоидны). Однако часто в интенсивно растущей культуре количество ДНК на клетку достигает массы равной трем, четырем, восьми и более хромосом. Из этого следует вывод, что термины «нуклеоид» и «хромосома» не всегда являются идентичными и в зависимости от условий, нуклеоид бактериальной клетки может состоять из одной или нескольких копий одной и той же хромосомы [1] .
Структура нуклеоида
Нуклеоид прокариот представлен одной замкнутой в кольцо двухспиральной нитью ДНК длиной 1,1–1,6 мм, которую рассматривают в качестве одиночной бактериальной хромосомы или генофора. Эта нить уложена в компактную структуру, локализованную в ограниченных участках цитоплазмы [2] [1] .
Нуклеоид бактерий отличается от ядра эукариотических клеток отсутствием ядерной мембраны, ядрышка и митотического способа деления и находится в непосредственном контакте с цитоплазмой клетки [3] .
В состав структур нуклеоида входят РНК-полимераза, основные белки. Хромосома закреплена на цитоплазматической мембране, а у грамположительных бактерий на мезосоме [1] .
Строение бактериальной клетки
Бактериальная клетка в целом устроена достаточно просто. Она отделена от внешней среды цитоплазматической мембраной и заполнена цитоплазмой, в которой располагается зона нуклеоида, включающая кольцевую молекулу ДНК, с которой может «свисать» транскрибируемая мРНК, к которой, в свою очередь прикреплены рибосомы, синтезирующие на ее матрице белок одновременно с процессом синтеза самой матрицы. Одновременно ДНК может быть связана с белками, осуществляющими ее репликацию и репарацию. Рибосомы бактерий меньше эукариотических и имеют конэфициент седиментации 70S. Они, как и эукариотические образованы двумя субъединицами – малой (30S), в состав которой входит 16S рРНК и большую – 50S, включающую молекулы 23S и 5S рРНК.
На фотографии, полученной с помощью трансмиссионной микроскопии (рис.1), отчетливо видна светлая зона, в которой находится генетический аппарат и происходят процессы транскрипции и трансляции. Рибосомы видны как мелкие зернистые включения.
Рисунок 1. Микрофотография бактериальной клетки
Чаще всего в бактериальной клетке геном представлен только одной молекулой ДНК, которая замкнута в кольцо, однако есть и исключения. У некоторых бактерий молекул ДНК может быть несколько. Например, Deinococus radiodurans, бактерия, известная своей феноменальной устойчивостью к радиации и способная спокойно выдерживать дозу радиации в 2 000 раз превышающую летальную дозу для человека, имеет две копии своей геномной ДНК. Известны бактерии, имеющие три или четыре копии. У некоторых видов ДНК может быть не замкнула в кольцо, а некоторые Agrobacterium содержат одну кольцевую и одну линейную ДНК.
Помимо нуклеоида, генетический материал может быть представлен в клетке в виде дополнительных маленьких кольцевых молекул ДНК – плазмид. Плазмиды реплицируются независимо от нуклеоида и зачастую содержат полезные для клетки гены, дающие клетке, например, устойчивость к антибиотикам способность к усвоению новых субстратов[1], способность к конъюгации и многое другое. Плазмиды могут передаваться как от материнской клетке к дочерней, так и путем горизонтального переноса быть переданы от одной клетке другой.
Бактериальная клетка чаще всего окружена не только мембраной, но и клеточной стенкой, причем по типу устройства клеточной стенки бактерий делят на две группы – грамположительные и грамотрицательные[2].
Клеточная стенка бактерий образована пептидогликаном – муреином. На молекулярном уровне муреиновый слой представляет собой сеть, образованную молекулами N-ацетилглюкозамина и N-ацетилмурамовой кислоты, сшитыех между собой в длинные цепи β-1-4-гликозидными связями, соседние цепи, в свою очередь соединяются поперечными пептидными мостиками (Рис.2). Так получается одна большая сеть, окружающая клетку.
Рисунок 2. Схема строения муреина
Грамположительные бактерии имеют толстую клеточную стенку, располагающуюся поверх мембраны. Муреин поперечно прошит еще одним типом молекул – тейхоевыми и липотейхоевыми (если они соединены с липидами мембраны) кислотами. Считается, что эти молекулы придают клеточной стенке эластичность при поперечном сжатии и растяжении, действуя как пружины. Поскольку слой муреина толстый, он легко окрашивается по методу Грама: клетки выглядят ярко-фиолетовыми, поскольку краситель (генциановый или метиловый фиолетовый) застревает в слое клеточной стенки.
Рисунок 3. Фиолетовые клетки - грамположительные, розовые - грамотрицательные
У грамотрицательных бактерий слой муреина очень тонкий (исключение составляют цианобактерии), поэтому при окрашивании по Граму фиолетовый краситель вымывается, а клетки окрашиваются в цвет второго красителя (рис. 3).
Рисунок 4. Схема строения бактериальной клетки. «Гр+» - грамположительные клетки, «Гр-» - грамотрицательные клетки.
Клеточная стенка грамотрицательных бактерий покрыта сверху еще одной, внешней, мембраной, прикрепленной к пептидокликану липопротеинами. Пространство между цитоплазматической мембраной и внешней мембраной называется периплазмой. Внешняя мембрана содержит липополипротеины, липополисахариды (ЛПС), а также белки, образующие гидрофильные поры. Компоненты внешней мембраны зачастую отвечают за взаимодействие клетки со внешней средой. Она содержит антигены, рецепторы фагов, молекулы, участвующие в конъюгации и др.
Поскольку у грамположительных и грамотрицательных клеток различается строение покровов (Рис. 4, сверху), отличается и аппарат, заякоривающий жгутик в клеточных покровах (Рис.4, снизу).
Жгутик грамположительных бактерий закрепляется в мембране двумя белковыми кольцами (S-кольцо и M-кольцо) и приводится в движение системой белков, которые, потребляя энергию, заставляют нить крутиться. У грамотрицательных бактерий в дополнение к этой конструкции есть еще два кольца, дополнительно фиксирующие жгутик во внешней мембране и клеточной стенке.
Сам по себе жгутик у бактерий состоит из белка флагеллина, субъединицы которого соединяются в спираль, имеющую внутри полость и формирующие нить. Нить гибко крепится к заякоривающему и придающему ей кручение аппарату, с помощью крючка.
Помимо жгутиков на поверхности клеток бактерий могут быть и другие выросты – пили. Это белковые ворсинки, позволяющие бактериям присоединяться к различным поверхностям (повышая гидрофобность клетки) либо принимающие участие в транспорте метаболитов и процессе конъюгации (F-пили).
Бактериальная клетка обычно не содержит никаких мембранных структур внутри, в том числе и везикул, но могут быть различного рода включения (запасные липиды, сера) и газовые пузырьки, окруженные белковой мембраной. Без мембраны клетка может запасать молекулы полисахаридов, цианофицин (как депо азота), а также может содержать карбоксисомы – пузырьки, содержащий фермент РуБисКО[3], необходимый для фиксации углекислого газа в Цикле Кальвина.
[1] В микробиологии этот термин означает питательное вещество, которое может быть усвоено микроорганизмом
[2] Такое название групп происходит от фамилии врача Г.К. Грама, разработавшего метод окраски клеточных стенок бактерий, позволяющий различать клетки с различным типом строения клеточной стенки.
Читайте также: