Строение октамера гистонов нуклеосомы
Добавил пользователь Евгений Кузнецов Обновлено: 07.01.2026
ГИСТОНЫ
ГИСТОНЫ (от греч. histos-ткань), группа сильноосновных простых белков (р/ 9,5-12,0), содержащихся в ядрах клеток животных и растений. Различают пять осн. групп гистонов, каждую из к-рых составляют белки с близкими св-вами, выделенные из разных организмов. Группы Н2А, Н2В, НЗ и Н4 имеют мол. м. от 11 до 14 тыс. (т. наз. низкомолекулярные гистоны), группа HI -ок. 22 тыс. Для первичной структуры гистонов характерно высокое содержание остатков лизина и аргинина, а также отсутствие триптофана. Гистоны одной и той же группы, полученные из разл. источников, имеют очень сходную первичную структуру. Так, гистоны из тимуса быка и проростков гороха, относящиеся к группе Н4, отличаются расположением только двух аминокислотных остатков. Во вторичной структуре преобладают структура появляется только при необратимой агрегации гистонов. Третичную структуру образует глобула (80-100 аминокислотных остатков), содержащая гл. обр. гидрофобные и кислые аминокислотные остатки; N-концевая (10-25 остатков), а в ряде случаев и С-концевая часть (5-10 остатков) не структурированы, подвижны и обогащены аргинином и особенно лизином. Группа HI отличается от др. групп значительно более длинным (ок. 100 остатков) подвижным N-концом.
Гистоны-осн. белковые компоненты (по массе) хромосомы. В ядре они тесно связаны с ДНК (их кол-ва по массе приблизительно равны), образуя цепочку нуклеопротеидных частиц-нуклеосом, представляющих собой низший уровень упаковки ДНК в хромосоме. Группы гистонов различаются по их роли в образовании нуклеосомного и последующих уровней, чем и вызвано их подразделение на три подгруппы: аргинин-богатые (НЗ и Н4), умеренно лизин-богатые (Н2А и Н2В) и лизин-богатые (HI и родственные ему Г).
Белковое ядро нуклеосомы-октамерный комплекс гистонов, существующий в своб. виде только в р-рах с высокой ионной силой и представляющий собой клиновидный диск размером 7,0 х 7,0 х 5,6 нм с осью симметрии С2. Основа октамера-прочный тетрамерный комплекс аргинин-богатых Г, (Н3—Н4)2, имеющий подковообразную форму. К ветвям "подковы" обратимо присоединены два димера умеренно лизин-богатых гистонов (Н2А-Н2В). На пов-сть октамера навивается ДНК (ок. 160 пар остатков нуклеотидов), образующая два витка левой суперспирали. Гистоны группы HI присоединяется своей глобулярной частью к периферии нуклеосомы, фиксируя места выхода из нее ДНК, а также располагается на межнуклеосомных участках ДНК. Этот же гистон с помощью длинных подвижных концевых сегментов осуществляет поперечные сшивки между нуклеосомами и ответствен за сворачивание их цепочки в структуры высшего порядка.
Гистоны в ядре подвергаются многочисл. постсинтетич. модификациям (напр., ацетилированию, фосфорилированию, метилированию), значение к-рых точно неизвестно. Ацетилирование гистонов обычно связывают с транскрипционно-активным состоянием хромосом.
Предполагают, что гистоны участвуют в переводе чрезвычайно длинных молекул хромосомной ДНК в форму, удобную для пространств. разъединения отдельных хромосом и их перемещения в ходе деления клетки. Полагают также, что гистоны участвуют в механизмах транскрипции и репликации. Так, угнетение синтеза РНК в зрелых эритроцитах птиц связывают с появлением в них гистона, относящегося к подгруппе HI.
Строение октамера гистонов нуклеосомы
• Октамер гистонов состоит из ядра, включающего тетрамер Н32-Н42, связанного с двумя димерами Н2А-Н2В
• Каждый гистон плотно переплетается со своими партнерами
• Все коровые гистоны имеют структурный мотив гистоновой складки
• N-терминальный конец гистона выходит из нуклеосомы
До сих пор мы рассматривали строение нуклеосомы с точки зрения организации ДНК на ее поверхности. Для того чтобы понять организацию белковой части нуклеосомы, мы должны знать, каким образом гистоны взаимодействуют друг с другом и с ДНК. Способны ли они к взаимодействию только в присутствии ДНК или могут независимо образовывать октамеры? Большая часть данных о межгистоновых взаимодействиях получена на основании исследований способности этих белков к образованию стабильных комплексов и из экспериментов по образованию белковых сшивок.
Коровые гистоны образуют два типа комплексов. Н3 и Н4 образуют тетрамер (Н32-Н42). Гистоны Н2А и Н2В образуют различные комплексы, наиболее часто димеры (Н2А-Н2В).
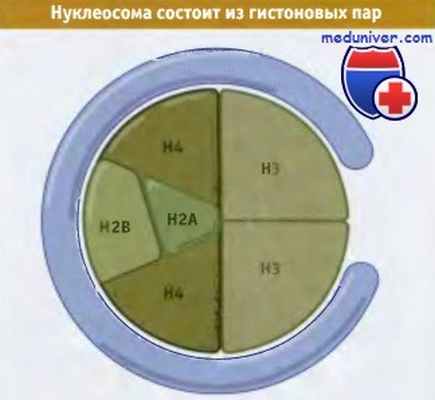
В симметричной модели нуклеосомы тетрамер гистонов Н32-Н34 обладает формой ядра.
Один димер Н2А-Н2В виден сверху, второй находится снизу.
Интактные октамеры гистонов можно получить или экстрагируя их из хроматина, или при ассоциации концентрированных растворов белков in vitro, содержащих много солей. Октамер может диссоциировать с образованием гексамера, и Н2А-Н2В димера гистонов. Это говорит о существовании в нуклеосоме центрального «ядра», содержащего тетрамер Н32-Н42. In vitro тетрамер участвует в организации ДНК в частицы, обладающие некоторыми свойствами коровых.
Эксперименты с применением сшивающих агентов показали, какие пары гистонов в нуклеосоме располагаются рядом. (Трудность интерпретации полученных данных состоит в том, что сшивается лишь небольшая часть белков, и поэтому следует с осторожностью экстраполировать эти данные на ядро нуклеосомы in vivo) Полученные данные использовались для построения модели организации нуклеосомы. Схематически она представлена на рис. 6.40.
Структурные исследования показывают, что изолированный октамер гистонов по форме напоминает коровую частицу. Это свидетельствует о том, что общая структура частицы задается в результате гистон-гистоновых взаимодействий. Расположение отдельных гистонов в октамере было установлено на основании их способности к взаимодействию друг с другом и экспериментов со сшивающими агентами.
На основании анализа кристаллической структуры (с разрешением 3,1 А) была предложена модель строения октамера гистонов. Исследование направления отдельных полипептидных элементов кристаллической структуры предполагает, что гистоны не организованы в виде индивидуальных глобулярных белков, а взаимосвязаны со своими партнерами: Н3 с Н4, и Н2А с Н2В. Таким образом, в модели отдельно выделены тетрамеры Н32-Н42 (белым цветом) и димеры Н2А-Н2В (голубым цветом), и не показаны индивидуальные гистоны.
На верхнем рисунке в таком же ракурсе представлена модель, схематически изображенная на рисунке ниже. Тетрамер Н32-Н42 составляет диаметр октамера. Он обладает формой подковы. Пара Н2А-Н2В представлена двумя димерами, но на рисунке показан один из них. Вид сбоку представлен в том же ракурсе, как и на рисунке ниже. На этом изображении можно четко видеть расположение тетрамера Н32-Н42 и двух димеров Н2А-Н2В. Белок образует некое подобие бобины, по поверхности которой проходит суперспиральная ДНК, делающая почти два полных витка вокруг нуклеосомы.
Модель характеризуется осью симметрии второго порядка, расположенной перпендикулярно боковой плоскости.
В основном связывание происходит с фосфодиэфирным скелетом (что обусловлено необходимостью упаковки любой ДНК, независимо от нуклеотидного состава). Как можно видеть в нижней части рисунка, тетрамер Н32-Н42 образуется за счет взаимодействия между двумя субъединицами Н3.
Каждый из коровых гистонов рассматривают как глобулярный белок, определяющий массу центрального белка нуклеосомы. Каждый гистон также обладает гибким N-концевым хвостом, способным модифицироваться, что играет важную роль в функциональной активности хроматина. Расположение хвостов, составляющих примерно четверть от массы белка, твердо не установлено, и на рисунке ниже показано произвольно.
Однако хвосты гистонов Н3 и Н2В проходят между витками суперспирали ДНК и выходят за пределы нуклеосомы, как показано на рисунке ниже. Под действием УФ между хвостами и ДНК образуются сшивки. При этом нуклеосомы образуют большее количество продуктов, чем коровые частицы. Это означает, что хвосты контактируют с линкерной ДНК. По-видимому, хвост гистона Н4 контактирует с димером Н2А-Н2В, находящимся в соседней нуклеосоме. Это взаимодействие может служить важным фактором, обеспечивающим общую структуру нуклеосомы.
Пространственная модель кристаллической структуры октамера коровых гистонов.
Тетрамер Н32-Н42 выделен белым цветом, а Н2А-Н2В димеры голубым.
Вид сверху показывает только один из Н2А-Н2В димеров, поскольку второй расположен снизу.
ДНК выделена зеленым цветом. Вид половины нуклеосомы сверху, показывающий расположение пар гистонов Н3-Н4 и Н2А-Н2В.
При наложении второй половины можно видеть симметричную организацию нуклеосомы. Глобулярные участки молекул гистонов локализуются внутри гистонового октамера коровой частицы,
однако расположение N-терминальных «хвостов», на которых располагаются сайты модификации,
неизвестно и может быть не столь строго фиксировано. N-терминальные «хвосты» гистонов организованы беспорядочно
и могут выходить между витками спирали ДНК, обвивающей коровые частицы.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
22. Гистоны. Структура нуклеосом.
Гистоны — основной класс нуклеопротеинов, ядерных белков, необходимых для сборки и упаковки нитей ДНК в хромосомы. Существует пять различных типов гистонов, названных H1/Н5, H2A, H2B, H3, H4. Последовательность аминокислот в этих белках практически не различается в организмах различного уровня организации. Гистоны — небольшие, сильно основные белки, связывающиеся непосредственно с ДНК. Гистоны принимают участие в структурной организации хроматина, нейтрализуя за счет положительных зарядов аминокислотных остатков отрицательно заряженные фосфатные группы ДНК, что делает возможной плотную упаковку ДНК в ядре.
По две молекулы каждого из гистонов Н2А, Н2В, Н3 и Н4 составляют октамер, обвитый сегментом ДНК длиной 146 п.о., образующим 1,8 витка спирали поверх белковой структуры. Эта частица диаметром 7 нм называется нуклеосомой. Участок ДНК (линкерная ДНК), непосредственно не контактирующий с гистоновым октамером, взаимодействует с гистоном Н1.
Группа негистоновых белков высоко гетерогенна и включает структурные ядерные белки, множество ферментов и факторов транскрипции, связанных с определенными участками ДНК и осуществляющих регуляцию генной экспрессии и других процессов.
Гистоны в октамере имеют подвижный N-концевой фрагмент («хвост») из 20 аминокислот, который выступает из нуклеосом и важен для поддержания структуры хроматина и контроля за генной экспрессией. Так, например, формирование (конденсация) хромосом связано с фосфорилированием гистонов, а усиление транскрипции — с ацетилированием в них остатков лизина. Детали механизма регуляции до конца не выяснены.
Нуклеосома - субъединица хроматина, состоящая из ДНК и набора из четырех пар гистоновых белков Н2А , Н2В , Н3 и Н4 одной молекулы гистона H1. Гистон Н1 связывается с линкерной ДНК между двумя нуклеосомами.
Нуклеосома является элементарной единицей упаковки хроматина. Она состоит из двойной спирали ДНК, обмотанной вокруг специфического комплекса из восьми нуклеосомных гистонов ( гистонового октамера ). Нуклеосома представляет собой дисковидную частицу с диаметром около 11 нм, содержащую по две копии каждого из нуклеосомных гистонов (Н2A, Н2В, НЗ, Н4 ). Гистоновый октамер образует белковую сердцевину, вокруг которой дважды обмотана двуспиральная ДНК (146 нуклеотидных пар ДНК на гистоновый октамер).
Нуклеосомы, входящие в состав фибрилл, расположены более или менее равномерно вдоль молекулы ДНК на расстоянии 10-20 нм друг от друга.
23. Уровни упаковки хромосом эукариот. Конденсация хроматина.
Таким образом, уровни упаковки ДНК следующие:
1) Нуклеосомный (2,5 оборота двуспиральной ДНК вокруг восьми молекул гистоновых белков).
2) Супернуклеосомный - хроматиновая спираль (хромонема).
3) Хроматидный - спирализованная хромонема.
4) Хромосома - четвертая степень сперализации ДНК.
В интерфазном ядре хромосомы деконденсированы и представлены хроматином. Деспирализованный участок, содержащий гены, называется эухроматин (разрыхленный, волокнистый хроматин). Это необходимое условие для транскрипции. Во время покоя между делениями определенные участки хромосом и целые хромосомы остаются компактными.
Эти спирализованные, сильно окрашивающиеся участки, называются гетерохроматином. Они неактивны в отношении транскрипции. Различают факультативный и конститутивный гетерохроматин.
Факультативный гетерохроматин информативен, т.к. содержит гены и может переходить в эухроматин. Из двух гомологичных хромосом одна может гетерохроматической. Конститутивный гетерохроматин всегда гетерохроматичен, неиформативен (не содержит генов) и поэтому всегда неактивен в отношении транскрипции.
Хромосомная ДНК состоит из более 108 пар оснований, из которых образуется информативные блоки - гены, расположенные линейно. На их долю приходится до 25% ДНК. Ген - функциональная единица ДНК, содержащая информацию для синтеза полипептидов, или всех РНК. Между генами находятся спейсеры - неинформативные отрезки ДНК разной длины. Избыточные гены представлены большим числом - 104 идентичных копий. Примером являются гены для т-РНК, р-РНК, гистонов. В ДНК встречаются последовательности одних и тех же нуклеотидов. Они могут быть умеренно повторяющимися и высоко повторяющимися последовательностями. Умеренно повторяющиеся последовательности достигают 300 пар нуклеотидов с повторениями 102 - 104 и представляют чаще всего спейсеры, избыточные гены.
Высокоповторяющиеся последовательности (105 - 106) образуют конститутивный гетерохроматин. Около 75% всего хроматина не участвует в транскрипции, он приходится на высокоповторяющиеся последовательности и нетранскрибируемые спейсеры.
Строение нуклеосомы
Гистоны играют важную роль в структурной и функциональной организации нуклеосомы. В статье представлена информация о структурных аспектах нуклеосомы, взаимодействиях между гистонами, межнуклеосомных контактах и вариантных формах гистонов.
Тезисы статьи , Leonardo Mariño-Ramírez, Maricel G Kann, Benjamin A Shoemaker, and David Landsman
Keywords: структура хроматина, вариантные формы гистонов, нуклеосома
Строение нуклеосомы
- Общая формула: ([H2A–H2B][2] [H2A–H2B])
- Вокруг нуклеосомы обернуто 147 п.н. (1.7 оборота ДНК)
- Каждый из коровых гистонов состоит из трех α-спиралей, соединенных двумя петлями, которые позволяют взаимодействовать гистонам по типу «рукопожатие» (handshake motif).
- N-концы гистонов подвергаются посттрансляционным модификациям (ПТМ), что влияет на состояние хроматина (конденсированное/декондесированное).
- Между нуклеосомами могут возникать интернуклеосомные взаимодействия (дисульфидные связи между N-концом H4 и второй α-спиралью H2A), что стабилизирует высшие структуры нуклеосомной укладки.
- Гистоны H2B и H2A наиболее вариабельны, H4 and H3 – наиболее консервативны.
Рис.1. Строение нуклеосомы.
А. Строение нуклеосомы и модификации гистонов. (Protein Data Bank accession code 1KX5). Цвета гистонов: голубой – H3, светло-зеленый – H4, оранжевый – H2A, желтый – H2B. Модификации гистонов показаны в виде шариков. Два димера H2A-H2B взаимодействуют с тетрамером H3–H4 через два взаимодействия H2B–H4. В.Домены гистонов H3 и H4 взаимодействуют по типу «рукопожатие»
Коровые гистоны контактируют с фосфодиэфирным остовом ДНК. ДНК обернута вокруг октамера с помощью трех-шести водородных связей между аминокислотными остатками гистонов и фосфодиэфирным остовом ДНК. Хвосты коровых гистонов экспонированы снаружи от ДНК и могут быть вовлечены в межнуклеосомные взаимодействия. Хвосты гистонов положительно заряжены (голубые участки), а центр октамера гистонов заряжен отрицательно. Эта разница в зарядах играет важную роль в межнуклеосомных взаимодействиях через гистоновые хвосты и вносит вклад в структурную пластичность нуклеосомной частицы.
Рис.2. Карта электростатических зарядов на поверхности и внутри нуклеосомной частицы (Protein Data Bank accession code 1KX5).
Электростатический потенциал варьирует от +20 (голубой) до −20 (красный) kTe−1. A. Вид нуклеосомы сбоку. Голубым отмечены положительные заряды. B. Вид нуклесомы сверху. Красным отмечены негативные заряды.
Взаимодействия гистонов в нуклеосоме
Таблица 1 (Protein Data Bank identifier 1KX5). Режимы связывания между коровыми гистонами. Режимы связывания были созданы на основе расположения аминокислотных цепей, доменов, участвующих в связывании, и числа остатков, контактирующих между собой.
Рис.3. Регионы аминокислотных последовательностей коровых гистонов, участвующих в связывании.
Каждый гистон представлен в виде черной линии. Участки, ответственные за связывание, показаны в виде цветных боксов, рисунок которых зависит от характера связывания.
Рис.4. Различные типы связывания гистонов в нуклеосоме
гистон H3 (голубой), H4 (светло-зеленый), H2A (оранжевый) и H2B (желтый) A Режим связывания 3 (взаимодействия между H3 и H2A). B Режим связывания 4 (взаимодействия между H4 и H2A). C Режим связывания 5 (взаимодействия между H3 и H3). D Режим связывания 6 (взаимодействия между H4 и H2B). E Режим связывания 7 (взаимодействия между H4 и H2B). F Режим связывания 8 (взаимодействия между H2A и H2A).
Рис.5. Консервативные регионы гистонов H4 и H2A, которые участвуют в межнуклеосомных взаимодействиях.
Множественные выравнивания гистонов H4 и H2A. Последовательности были взяты из Histone Database. Элементы вторичной структуры показаны как линии (петли) и боксы (спирали). А. Выравнивание гистона Н4. Лизины 16 и 20 могут подвергаться ПТМ. Б. Выравнивание гистона H2A.
Вариантные формы гистонов
Многие эукариотические организмы имеют несколько копий генов гистонов. Большинство генов гистонов представлено в виде кластера генов и экспрессируются в S-фазе клеточного цикла. Эти гистоны используются для сборки нуклеосомы и упаковки новосинтезированной ДНК. Однако некоторые вариантные формы гистонов могут экспрессироваться на всем протяжении клеточного цикла, и их экспрессия не ограничена S-фазой.
Гистон H2A
Гистон H2A имеет наибольшее количество вариантных форм, которые включают H2AZ, H2AX, H2A. Bbd и MacroH2A. Различаются С-концевыми последовательностями и локализацией в геноме.
- H2AZ ассоциирован с активацией транскрипции у дрожжей.
- H2A.Bbd имеет усеченный С-конец, и его локализация коррелирует с транскрипционно-активным хроматином.
- MacroH2A был найден в неактивных Х-хромосомах, имеет макродомен с энзиматической активностью. H2AX имеет на С-конце консенсус SQ[E/D]Φ, где Φ – гидрофобная аминокислота. Остаток серина в консенсусе фосфорилируется в ответ на двунитевые разрывы ДНК (DSBs). Также фосфорилирование H2AX привлекает белки, вовлеченные в репарацию ДНК.
Гистон H3
Вариантные формы гистона Н3 сильно расходятся по последовательности на N-конце c каноническим Н3.
- H3.3 – найден в транскрипционно активном хроматине.
- CenHS – локализован в прицентромерном хроматине.
- H3.4 – содержится в первичных сперматоцитах.
Гистон H4
Гистон H4 – наиболее консервативный гистон. H4 имеет много контактов с другими коровыми гистонами. На данный момент не известны вариантные формы.
Нуклеосома
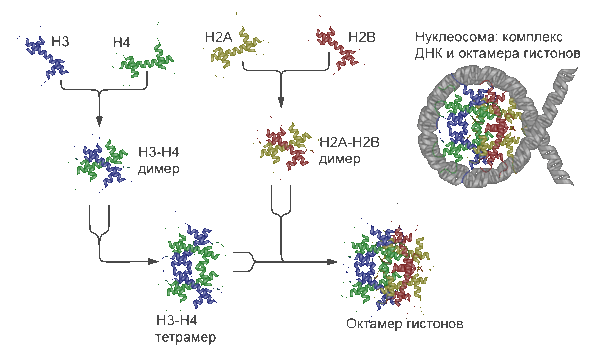
Структура и «сборка» нуклеосомы. Показано образование и строение гистонного октамера и структура комплекса ДНК-гистонный октамер — нуклеосомы.
Нуклеосома — это структурная часть хромосомы, образованная совместной упаковкой нити ДНК с гистоновыми белками H2А, H2B, H3 и H4. Последовательность нуклеосом, соединенная гистоновым белком H1, формирует нуклеофиламент (nucleofilament), или иначе нуклеосомную нить.
Вокруг нуклеосомного ядра, представленного гистонным октамером, ДНК делает 1,67 оборота (147 п.н.). Участок ДНК, между нуклеосомами, называется линкерной ДНК и составляет 10—100 п.н.
Сборка нуклеосомы происходит на ДНК. При репликации ДНК материнские гистоны распределяются случайным образом по дочерним цепям. Гистоновые шапероны временно экранируют заряд гистонов, обеспечивая правильную сборку нуклеосомы. Шаперон CAF1 связан с PCNA, сидит в репликационной вилке, связывая «старые» димеры H3H4, начинает пострепликационную сборку нуклеосом с посадки этого димера.
Последовательности ДНК могут в 1000 раз отличаться по потенциалу связывать нуклеосому. Если подряд следуют последовательности, изгибающие ДНК в одну сторону (например, ТАТА), связывание нуклеосомы будет неустойчиво.
В геноме присутствуют:
- участки, свободные от нуклеосом (сайты связывания транскрипционных акторов, регуляторных белков);
- участки, где положение нуклеосомы строго фиксировано;
- участки, в которых нуклеосомная укладка подвержена регуляции белками АТФ-зависимого ремоделинга хроматина.
Литература
- Хромосомы
- Эпигенетическое наследование
- ДНК
Wikimedia Foundation . 2010 .
Полезное
Смотреть что такое "Нуклеосома" в других словарях:
Нуклеосома — * нуклеасома * nucleosome or nu(v) particle дискообразные структуры эукариотических хромосом диаметром ок. 10 нм, являющиеся элементарной единицей упаковки хромосмной ДНК в хроматине. Состоит из белкового ядра, включающего октомеры из четырех пар … Генетика. Энциклопедический словарь
нуклеосома — nucleosome, nu (ν) particle нуклеосома. Дисковидная структура диаметром около 10 нм, являющаяся элементарной единицей упаковки хромосомной ДНК в хроматине ; состоит из белкового ядра (включает октамер гистонов Н2 … Молекулярная биология и генетика. Толковый словарь.
Нуклеосома — дисковидная структура, являющаяся элементарной единицей упаковки хромосомной ДНК в хроматине. Состоит из белка (гистона) и обернутой вокруг него двойной спирали ДНК … Словарь по психогенетике
минимальная нуклеосома — core particle, minimal nucleosome коровая частица, минимальная нуклеосома. Единица упаковки ДНК, стабильно существующая при формировании нуклеосомной структуры и включающая 146 пар нуклеотидов и октамер коровых гистонов ;… … Молекулярная биология и генетика. Толковый словарь.
Клеточное ядро — Клетки HeLa, ДНК которых окрашена голубым красителем Хёхста 33258. Центральная и правая клетки находятся в интерфазе, по … Википедия
Кариоплазма — Клетки ДНК которых окрашена голубым красителем Хойста. Центральная и правая клетки находятся в интерфазе, поэтому окрашено всё ядро. Клетка слева находится в состоянии митоза (анафаза), поэтому её ядро не видно, а ДНК сконденсирована так, что… … Википедия
Нуклеоплазма — Клетки ДНК которых окрашена голубым красителем Хойста. Центральная и правая клетки находятся в интерфазе, поэтому окрашено всё ядро. Клетка слева находится в состоянии митоза (анафаза), поэтому её ядро не видно, а ДНК сконденсирована так, что… … Википедия
Эукариотическое ядро — Клетки ДНК которых окрашена голубым красителем Хойста. Центральная и правая клетки находятся в интерфазе, поэтому окрашено всё ядро. Клетка слева находится в состоянии митоза (анафаза), поэтому её ядро не видно, а ДНК сконденсирована так, что… … Википедия
Ядерная оболочка — Клетки ДНК которых окрашена голубым красителем Хойста. Центральная и правая клетки находятся в интерфазе, поэтому окрашено всё ядро. Клетка слева находится в состоянии митоза (анафаза), поэтому её ядро не видно, а ДНК сконденсирована так, что… … Википедия
Читайте также: