Строение политенных хромосом
Добавил пользователь Skiper Обновлено: 29.01.2026
Политенные хромосомы — это гигантские хромосомы, обнаруживаемые в интерфазных клетках некоторых тканей у мух и других двукрылых. Такие хромосомы есть у них в клетках слюнных желез, мальпигиевых сосудов и средней кишки. Они содержат сотни нитей ДНК, которые редуплицировались, но не подверглись расхождению. При окраске в них выявляются четко выраженные поперечные полосы или диски. Многие отдельные полосы соответствуют местоположению отдельных генов. Ограниченное число определенных полос в некоторых дифференцированных клетках образует вздутия, или пуфы, выступающие за пределы хромосомы. Эти вздутые участки находятся там, где гены наиболее активны в отношении транскрипции. Политенные хромосомы развиваются из хромосом диплоидных ядер за счёт последовательных дупликаций каждого элемента. Помимо увеличения размеров ядра и размеров клетки, политенные хромосомы, так как содержат большое число копий генов, усиливают их экспрессию (процесс, в ходе которого наследственная информация от гена преобразуется в функциональный продукт — РНК или белок). Это, в свою очередь, увеличивает производство необходимых специализированной клетке белков. Например, чтобы образовывать большое количество клейкого вещества до окукливания. В других случаях тандемная дупликация участков приводит к возникновению мутации - изменению формы глаза.
При этом образуются так называемые политенные хромосомы, содержащие многие сотни
копий ДНК. Такие хромосомы обнаруживаются, например, в неделящихся клетках слюнных желез личинок некоторых насекомых при обычной световой микроскопии. В ходе митотического деления обеспечивается закономерное распределение сестринских хроматид каждой хромосомы между дочерними клетками. В составе дочерних хромосом (бывших сестринских хроматид) каждая клетка нового поколения получает одну из двух молекул ДНК, образовавшихся в результате репликации материнской двойной спирали. Следовательно, новое поколение клеток получает одинаковую генетическую информацию в составе каждой группы сцепления.
8. Белки, их химический состав, уровни структурной организации. Биологическая роль белков. Понятие о гистоновых и негистоновых белках. Прионовые белки и их медицинское значение.
Белки - непериодические полимеры, мономерами которых являются аминокислоты.Все белки представляют собой полимеры, состоящие из длинных цепей субъединиц, соединенных вместе в линейную структуру. Субъединицы - это 20 разных аминокислот.Общим признаком для всех аминокислот является наличие в их составе карбоксильной и аминогрупп, соединенных атомом углерода. Кроме этих общих атомов, каждая аминокислота содержит свои особые боковые цепи, присоединенные к центральному атому углерода. Таким образом, хотя все аминокислоты принадлежат к одному классу соединений и имеют некоторые общие химические свойства, отдельные аминокислоты резко отличаются друг от друга.
Уровни структурной организации
В строении молекул белков различают 4 уровня организации:
Первичная структура - полипептидная цепь из аминокислот, связанных в определенной последовательности ковалентными пептидными связями;
Вторичная структура - полипептидная цепь в виде спирали. Между пептидными связями соседних витков и другими атомами возникают многочисленные водородные связи, обеспечивающие прочную структуру;
Третичная структура - специфическая для каждого белка конфигурация - глобула. Удерживается малопрочными гидрофобными связями или силами сцепления между неполярными радикалами, которые встречаются у многих аминокислот.
Четвертичная структура возникает при соединении нескольких макромолекул, образующих агрегаты. Так, гемоглобин крови человека представляет агрегат из четырех макромолекул.
Нарушение природной структуры белка называют денатурацией. Она возникает под воздействием высокой температуры, химических веществ, лучистой энергии и др. факторов.
Биологическая роль белков
Строительная (структурная) функция: белки - строительный материал организма (оболочки, мембраны, органоиды, ткани, органы);
Каталитическая функция - ферменты, ускоряющие реакции в сотни миллионов раз;
Опорно-двигательная функция - белки, входящие в состав костей скелета, сухожилий; движение жгутиковых, инфузорий, сокращение мышц;
Транспортная функция - гемоглобин крови;
Защитная - антитела крови обезвреживают чужеродные вещества;
Энергетическая функция - при расщеплении белков 1 г освобождает 17,6 кДж энергии;
Регуляторная и гормональная - белки входят в состав многих гормонов и принимают участие в регуляции жизненных процессов организма;
Рецепторная - белки осуществляют процесс избирательного узнавания отдельных веществ и их присоединение к молекулам.
Понятие о гистоновых и негистоновых белках
Гистоны — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и во вторичной регуляции таких ядерных процессов, как транскрипция, репликация и репарация, проявляют сильно оснóвные свойства.
Негистоновые белки - это все белки хроматина, кроме гистонов, выделяющиеся с хроматином или хромосомами. Это сборная группа белков, отличающихся друг от друга как по общим свойствам, так и по функциональной значимости. Около 80% из негистоновых белков относится к белкам ядерного матрикса, обнаруживаемых как в составе интерфазных ядер, так и митотических хромосом. Гистоновые и негистоновые белки принимают участие в экспрессии генов, участвуют в создании структуры молекулы ДНК. Гистоновые белки - факторы репрессии (блокирования) генов, негистоновые - наоборот способствуют считыванию информации. Взаимодействие гистоновых и негистоновых белков – механизм блокирования и разблокирования молекулы ДНК.
Прионовые белки и их медицинское значение.
Прионы – модифицированные прионовые белки (содержат > 50 полиглютаминовых фрагментов). Прионовые белки – нейромедиаторы и регуляторы циркадных ритмов. Попадая в организм человека, прионы модифицируют (переделывают под себя) нормальные прионовые белки и вызывают следующие болезни: Куру, Кройцфельта-Якоба, смертельная семейная бессонница, Подострый спонгиозный трансмиссийный энцефалит и проч. Способы «заражения»: спонтанное возникновение в мозге прионов, ятрогения, наследственность, употребление в пищу «зараженного» мяса
Строение политенных хромосом
Цель: установить особенности строения политенных хромосом.
Контрольные вопросы:
- 1. Свойства и функции хромосом.
- 2. Генетическая организация политенных хромосом. Взаимоотношение генов и дисков.
- 3. Биохимические процессы, связанные с образованием пуфов.
Лабораторная работа
Изучение функции и морфологии политенных хромосом слюнных
желез дрозофилы
Объект исследования: слюнные железы 4—5-дневных личинок
3-го возраста (2-й стадии) дрозофилы.
Пояснение к заданию
Открытие политенных хромосом было сделано итальянским ученым Бальбиани в 1881 г. при изучении слюнных желез личинок хирономуса. Позже этот тип хромосом был обнаружен в соматических клетках тканей таких органов личинок двукрылых, как кишечник, мальпигиевые сосуды, жировое тело, а также в синергидах, антиподах, эндосперме некоторых растений, в трофобластах млекопитающих, в гигантских нейронах моллюсков, в клетках пищеварительных желез и эпителия матки аскариды, в макронуклеусе инфузорий.
Эти хромосомы в 100—200 раз длиннее и в 1000 раз больше содержат хромонем, чем обычные метафазные хромосомы, именно это и позволяет провести изучение хромомерного рисунка, а также функций отдельных генов. Они являются следствием явления политенния, в основе которого лежит эндомитоз, приводящий к увеличению числа жизненных циклов без расхождения хромосом. Так, например, степень политеннии дрозофилы равна 2 10 , что свидетельствует о 10 циклах, идущих подряд без карно- и цитокенеза.
«Классические» политенные хромосомы (результат истинной политеннии), имеющие синапс по всей длине как следствие максимальной конъюгации гомологичных хромосом с образованием цилиндрических жгутов, были описаны у ряда объектов, в том числе и у дрозофилы (слюнные железы личинок; рис. 1.10).
Каждая хромосома состоит из серии чередующихся междисков и дисков, количество которых значительно преобладает и составляет около 5000. Диски представлены в большей степени повторами, междиски — уникальными районами, в основном это регуляторные гены. Те и другие выступают в качестве маркеров, с помощью которых можно построить цитогенетическую карту хромосом и провести их идентификацию.
Центромеры хромосом дрозофилы агрегируют с образованием хромоцентра, который состоит в значительной степени из гетерохроматина. Из этого полицентрального образования выходит пять, реже шесть концов — лент, каждая лента представлена двумя гомологичными хромосомами, как результат конъюгации.
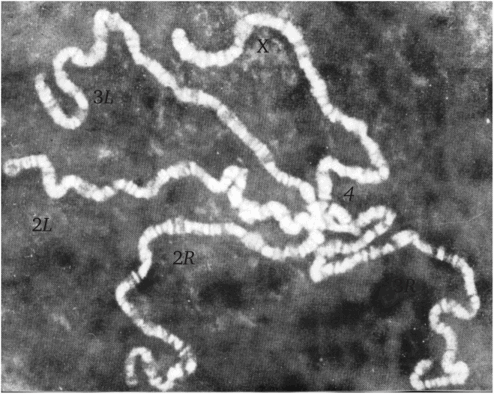
Рис. 1.10. Политенные хромосомы слюнных желез дрозофилы
Все хромосомы отличаются по морфологическому типу. Первая хромосома — X, акроцентрическая, образует одну ленту. Вторая и третья хромосомы очень длинные, метацентрические. Они образуют по две ленты (два плеча) от хромоцентра (21-левое, 2Я-правое; 31-левое, ЗК-правое). У L плеча концы веерообразные, у 3L плеча конец более расширенный с дисками у основания, у 2L — более ровный, диски отсутствуют. У R плеча концы бульбообразные, у 3R плеча — больших размеров, чем 2R плеча. Четвертая хромосома очень маленькая, она образует малую, едва выступающую из хромоцентра ленту.
Генетические исследования политенных хромосом позволяют провести картирование точек разрывов хромосомных перестроек, картирование генов при использовании метода гибридизации in situ, установить характер влияния различных факторов на процессы репликации, транскрипции. Политенных хромосом можно отнести к био-тест- объекту, позволяющему фиксировать последствия действия антропогенных факторов.
Техника приготовления временных препаратов политенных хромосом слюнных желез личинки дрозофилы
- 1. Пробирки с личинками дрозофилы 3-го возраста 2-й стадии (активно двигающиеся по стеклу) предварительно перед началом исследования поместите в термостат с температурой 16—18 °С на сутки.
- 2. Препарировальной иглой личинки перенесите из пробирки на предметное стекло в физиологический раствор. Все последующие операции проводите на предметном столике (черная сторона) бинокулярной лупы.
Приготовление физиологического раствора:
- 1- й вариант (для учебных исследований) — 7,5 г NaCl, 0,35 г КС1, 0,21 г СаС12 растворить в 1 л воды;
- 2- й вариант (для научных исследований) — 85 мг 12Н20 • Na2HP04, 50 мг КН2Р04, 620 мг NaCl, 200 мг КС1, 130 мг MgCl2 • 6Н20, 28 мг 4Н20 • • СаС12
- 3. С помощью двух препарировальных игл отделите слюнные железы личинки дрозофилы — парные образования удлиненной формы, расположенные по обе стороны пищевода в переднем отделе тела. Для этого необходимо одной иглой в левой руке сильно прижать ротовую часть личинки (район 1, рис. 1.11), а другой, взятой правой рукой, давить плашмя на середину тела (район 2, рис. 1.11), оттягивая конец личинки, отделяя две части друг от друга. В результате этой операции на предметном стекле останутся отпрепарированные слюнные железы с жировыми телами. Попытайтесь отделить и жировые тела. Препари- ровальными иглами или пинцетом перенесите слюнные железы на другое предметное стекло с ацетоорсеином.
Рис. 1.1 7. Личинка дрозофилы
Приготовление 2%-го ацетоорсеина: 4 гр орсеина (порошок) соединить с 90 мл ледяной уксусной кислотой, нагреть до кипения (повторить 15 раз), добавить 110 мл дистиллированной воды и довести до кипения еще раз (повторить 15 раз). Охладить, профильтровать.
- 4. Окрашивать 15—30 минут (в зависимости от качества красителя). После окрашивания пинцетом слюнные железы перенесите на другое предметное стекло с 2—3 каплями 45%-ной уксусной кислоты.
- 5. Накройте препарат покровным стеклом.
- 6. Поверх покровного стекла положите фильтровальную бумагу и подушечкой большого пальца с силой надавите на препарат.
- 7. На увеличении микроскопа 40 х 8 начните изучение препарата, найдите место, где хромосомы хорошо распределены и четко виден окрашенный узел — хромоцентр, соединяющий центромеры всех хромосом.
- 8. Опишите каждую хромосому, указав морфологический тип (расположение центромеры, соотношение длины плеч, размеры).
- 1. Приготовить 3—5 препаратов каждому студенту.
- 2. На увеличении 40 х 8 зарисовать общий вид политенных хромосом.
- 3. На увеличении 40 х 15 изучить и зарисовать участки гигантских хромосом.
Оборудование: предметные и покровные стекла; пинцет; салфетки; 45%-ная уксусная кислота; физиологический раствор, ацетоорсеин, препарировала ные иглы; фильтровальная бумага, полоски 2x5 см; бинокулярная лупа; микроскоп.
Политенные хромосомы
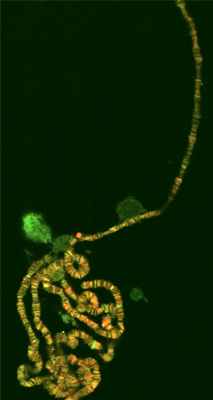
Полите́нные хромосо́мы — гигантские скопления объединённых хроматид, возникающие в некоторых типах специализированных клеток. Классическим примером являются гигантские хромосомы в клетках слюнных желёз личинок Drosophila melanogaster.
Репликация ДНК в этих клетках не сопровождается делением клетки, что приводит к накоплению вновь построенных нитей ДНК. Как клетки, так и политенные хромосомы значительно увеличиваются в размерах, что облегчает наблюдение и позволяло изучать активность генов ещё в 1930-е годы.
Содержание
Впервые политенные хромосомы были описаны Е. Бальбиани в 1881 году в клетках слюнных желёз представителя рода Chironimus из семейства комары-звонцы ( Chironomidae ) [1] . Тем не менее, природа этих структур не была доподлинно известна до их изучения у D. melanogaster Эмилем Хайтцем и Хансом Бауэром в начале 1930-х. В дальнейшем такие гигантские хромосомы были описаны у личинок двукрылых ( Diptera ) в ядрах клеток кишечника, мальпигиевых сосудов (например, у Sciara), а также у некоторых растений в ядрах синергид (например, гороха ( Pisum )) [2] .
Политенные хромосомы во много раз превышают по размеру хромосомы обычных соматических клеток. Они, как правило, в 100—200 раз длиннее и в 1000 раз толще (содержат до 1000 хромосом), чем хромосомы многих интерфазных клеток (как половых, так и соматических). Так, у личинок D. melangaster общая длина четырёх пар хромосом в слюнных железах составляет 2000 мкм, а в обычных соматических клетках эта величина равна 7,5 мкм [2] .
Политенные хромосомы в клетке слюнной железы у представителя рода Chironimus из семейства комары-звонцы ( Chironomidae )
Характерная форма и размеры политенных хромосом достигаются вследствие их максимальной деспирализации и многократного воспроизведения хромосом без их последующего расхождения, то есть они образуются как результат эндомитоза [2] .
Политенные хромосомы имеют характерную поперечную исчерченность, обусловленную наличием участков более плотной спирализации хромонем — хромомер. В тёмных участках (то есть хромомерах) располагается спирализованный неактивный хроматин, в то время как светлые полосы указывают на участок с повышенной транскрипционной активностью. Чёткое различение тёмных дисков и светлых междисковых участков объясняется нерасхождением дочерних хромонем. По этой причине все особенности отдельной хромонемы, в том числе хромомерный рисунок, становятся выраженными более контрастно [2] .
По сути, политенные хромосомы представляют собой пару гигантских гомологичных хромосом, находящихся в состоянии идеально точной соматической конъюгации. При этом диски и междисковые участки гомологов расположены строго параллельно и тесно сближены. Такая конъюгация не характерна для подавляющего большинства соматических клеток [2] .
Впервые карта политенных хромосом была составлена в 1935 году Кэлвином Бриджесом, и она широко используется и по сей день.
Уникальность строения политенных хромосом, а именно возможность чётко различать детали их строения, была использована Т. Пайтнером для изучения их перестроек и характера конъюгации [2] . Вообще, исчерченность политенных хромосом исключительно полезна для исследований, в частности, на примере политенных хромосом была получена визуализация участков активного и неактивного хроматина. На них также можно изучать общую структуру хроматина.
Кроме того, политенные хромосомы помогают идентифицировать личинок комаров-звонцов (Chironomid), которых другим способом отличить сложно [3] [4] .
В политенных хромосомах процесс транскрипции сопровождается формированием т. н. пуфов — характерных вздутий определённых дисков, образующихся в результате локальной декомпактизации в них ДНК. На активную транскрипцию в этих регионах указывает активное включение 3 H-уридина в районе пуфов. Крупные пуфы называются кольцами Бальбиани (в некоторых источниках термины «пуф» и «кольца Бальбиани» употребляют как синонимичные) [2] .
Пуфирование характерно для стадии личинки. Образование и исчезновение пуфов регулируется внутренней средой организма в соответствии со стадией развития. Одним из важнейших регуляторов образования пуфов у насекомых являются стероидные гормоны, в частности, гормон линьки — экдизон. Выявлено также влияние белков, синтезированных более ранними пуфами, на развитие более поздних пуфов [2] .
Таким образом, образование пуфов является ярким примером дифференциальной транскрипции. Другим известным примером этого процесса является формирование хромосом типа ламповых щёток [2] .
Помимо увеличения размеров ядра и размеров клетки, политенные хромосомы, так как содержат большое число копий генов, усиливают их экспрессию. Это, в свою очередь, увеличивает производство необходимых специализированной клетке белков. Например, в клетках слюнных желёз личинок D. melanogaster хромосомы подвергаюся множеству кругов эндоредупликации, чтобы образовывать большое количество клейкого вещества до окукливания.
В других случаях тандемная дупликация участков, расположенных вблизи центромеры Х-хромосомы, наблюдающаяся в клетках слюнных желёз и кишечника, приводит к возникновнении мутации Bar, проявляющеся в изменении формы глаза [5] .
Строение политенных хромосом
Строение политенных хромосом
• Политенные хромосомы двукрылых характеризуются наличием серий полос-дисков, рисунок которых можно использовать в качестве цитологической карты
Интерфазные ядра в некоторых тканях личинок двукрылых мух содержат хромосомы во много раз большего размера, чем обычные хромосомы. У этих хромосом увеличены диаметр и длина. На рисунке ниже в качестве примера представлен хромосомный набор клеток слюнной железы D. melanogaster. Такие хромосомы называют политенными.
Каждая хромосома состоит из видимой серии полос-дисков (которые более правильно, но редко называют хромомерами). Существует примерно десятикратное различие в размере дисков. Наибольшая ширина составляет около 0,5 мкм, а наименьшая — 0,05 мкм. (Наименьшие диски видны только в электронном микроскопе.) В дисках находится большая часть массы ДНК, и они интенсивно прокрашиваются соответствующими красителями.
Участки между отдельными дисками окрашиваются светлее; их называют междисками. В хромосомном наборе D. melanogaster содержится около 5000 дисков.
Центромеры всех четырех хромосом D. melanogaster агрегируют с образованием хромоцентра, который в значительной степени состоит из гетерохроматина (а у самцов включает целиком Y-хромосому). С учетом этого можно считать, что около 75% гаплоидного набора ДНК организовано в диски и междиски. В растянутом состоянии эта ДНК представляет собой линейную структуру, длиной примерно 40 000 мкм. Длина всего хромосомного набора составляет примерно 2000 мкм, т. е. примерно в 100 раз превышает длину гаплоидного набора в митозе.
Это ясно показывает, в каком растянутом состоянии находится генетический материал в политенной хромосоме по сравнению с его обычными состояниями в интерфазном хроматине или в митотических хромосомах.
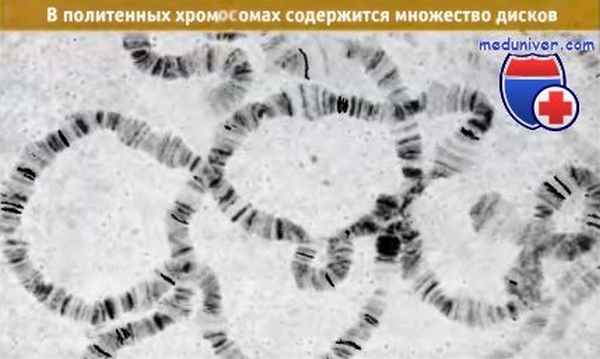
Политенные хромосомы D. melanogaster образуют серии дисков и междисков.
Какова структура этих гигантских хромосом? Каждая хромосома образуется в результате последовательных актов репликации конъюгировавшей пары гомологов. Реплики не разделяются, а остаются прикрепленными друг к другу по всей длине. В начале процесса каждая конъюгировавшая пара содержит количество ДНК, равное 2С (где С — это содержание ДНК в одной индивидуальной хромосоме). Затем число удвоений происходит до 9 раз; в результате максимальное содержание ДНК может достигнуть 1024С. Число удвоений варьирует в различных тканях личинки D. melanogaster.
Каждая политенная хромосома видна в виде большого числа параллельных нитей, сильно сконденсированных в дисках и слабее в междисках. Вероятно, каждая такая нить представлена единичной копией (С) гаплоидной хромосомы. Отсюда и происходит термин «политения». Степень политении определяется числом гаплоидных хромосом, содержащихся в гигантской хромосоме.
Для каждой линии Drosophila характерен свой набор дисков. Постоянство числа и линейного расположения дисков впервые было отмечено в 1930-х годах, когда стало понятно, что диски образуют цитологическую карту хромосом. В результате перестроек — таких как делеции, инверсии или дупликации, — происходит изменение линейного порядка дисков.
Линейный порядок дисков соответствует линейному расположению генов. Таким образом, генетические перестройки, как это видно по карте сцепления, можно привести в соответствие со структурными перестройками на цитологической карте. В конечном итоге, конкретную мутацию можно локализовать в определенном диске. Поскольку общее число генов у D. melanogaster превышает число дисков, в большинстве дисков, вероятно, локализуется множество генов.
Положение определенных генов на цитологической карте можно определить непосредственно с помощью метода гибридизации in situ. Общая схема метода представлена на рисунке ниже. Радиоактивная проба, представляющая некий ген (чаще всего меченный клон кДНК, полученный с иРНК), гибридизуется с денатурированной ДНК политенных хромосом in situ. Методом радиоавтографии можно определить местоположение одного или нескольких исследуемых генов по скоплению зерен серебра в области одного или нескольких дисков. Недавно вместо радиоактивных зондов стали использовать флуоресцентные.
Используя этот метод, оказывается возможным непосредственно выявить полосу, в которой локализована специфическая последовательность.
Индивидуальные диски, содержащие определенные гены, можно идентифицировать методом гибридизации in situ. При большом увеличении показана гибридизация in situ на дисках 87А и 87С с меченной ДНК,
выделенной из клеток, подвергнутых действию теплового шока.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Политенные хромосомы
Федеральное государственное бюджетное образовательное учреждение высшего
образования «Московский педагогический государственный университет»
_________________________________________________________________________
Факультет: Институт биологии и химии
Профиль: Биология и экология (педагогическое образование)
«ПОЛИТЕННЫЕ ХРОМОСОМЫ»
Выполнила:
Студентка 1 курса
102 группы 3 погруппы
Залогина Светлана
2. Содержание:
3. Что такое политенные хромосомы?
Это гигантские хромосомы, обнаруживаемые в
интерфазных клетках некоторых тканей у мух и
других двукрылых. Такие хромосомы есть у них
в клетках слюнных желез, мальпигиевых сосудов
и средней кишки. Они содержат сотни нитей
ДНК, которые редуплицировались, но не
подверглись расхождению. При окраске в них
выявляются четко выраженные поперечные
полосы или диски. Многие отдельные полосы
соответствуют
местоположению
отдельных
генов. Ограниченное число определенных полос
в некоторых дифференцированных клетках
образует вздутия, или пуфы, выступающие за
пределы хромосомы. Эти вздутые участки
находятся там, где гены наиболее активны в
отношении транскрипции. Содержатся в клетках
слюнных желез, кишечника, трахей, жирового
тела и мальпигиевых сосудов личинок
двукрылых.
4. История
Впервые обнаружены Е.Г. Бальбиани в 1881г,
однако их цитогенетическая роль была
выявлена Гейтцем и Бауером. В дальнейшем
такие гигантские хромосомы были описаны у
личинок двукрылых в ядрах клеток кишечника,
мальпигиевых
сосудов,
а
также
у
некоторых
растений
в
ядрах синергид (например, гороха). Термин
«политенная хромосома» предложил П. Коллер
(P. Koller) в 1935 году, а окончательно ввёл в
науку С. Дарлингтон в 1937 году
5. Размеры политенных хромосом
Политенные хромосомы во много раз
превышают
по
размеру
хромосомы
обычных соматических клеток. Они, как
правило, в 100—200 раз длиннее и в 1000 раз
толще (содержат до 1000 хромосом), чем
хромосомы многих интерфазных клеток
(как половых, так и соматических). Так, у
личинок D. melangaster общая длина
четырёх пар хромосом в слюнных железах
составляет 2000 мкм, а в обычных
соматических клетках эта величина равна 7,5
мкм
6. Строение. Исчерченность.
Политенные хромосомы имеют характерную
поперечную
исчерченность,
обусловленную наличием участков более
плотной спирализации хромонем —
хромомер. В тёмных участках (то есть
хромомерах)
располагается
спирализованный неактивный хроматин,
в то время как тёмные полосы указывают
на
участок
с
повышенной
транскрипционной активностью. Чёткое
различение тёмных дисков и светлых
междисковых
участков
объясняется
нерасхождением дочерних хромонем. По
этой причине все особенности отдельной
хромонемы, в том числе хромомерный
рисунок, становятся выраженными более
контрастно.
7. Строение. Пуфы.
В
политенных хромосомах процесс
транскрипции
сопровождается
формированием т. н. пуфов —
характерных вздутий определённых
дисков, образующихся в результате
локальной декомпактизации в них
ДНК. На активную транскрипцию в
этих регионах указывает активное
включение H-уридина в районе
пуфов.
Крупные
пуфы
называются кольцами Бальбиани (в
некоторых источниках термины «пуф»
и «кольца Бальбиани» употребляют
как синонимичные).
Пуфирование характерно для стадии личинки. Образование и исчезновение
пуфов регулируется внутренней средой организма в соответствии со стадией
развития. Одним из важнейших регуляторов образования пуфов у насекомых
являются стероидные гормоны, в частности, гормон линьки — экдизон.
Выявлено также влияние белков, синтезированных более ранними пуфами, на
развитие более поздних пуфов.
8. Функции.
Помимо увеличения размеров ядра и размеров
клетки, политенные хромосомы, так как содержат
большое число копий генов, усиливают их
экспрессию. Это, в свою очередь, увеличивает
производство необходимых специализированной
клетке белков. Например, в клетках слюнных желёз
личинок D. melanogaster хромосомы подвергаются
множеству кругов эндоредупликации, чтобы
образовывать большое количество клейкого
вещества до окукливания.
В других случаях тандемная дупликация участков,
расположенных вблизи центромеры Х-хромосомы,
наблюдающаяся в клетках слюнных желёз и
кишечника, приводит к возникновению мутации ,
проявляющееся в изменении формы глаза
Читайте также: