Запасание аминокислот. Физиология белков плазмы крови
Добавил пользователь Алексей Ф. Обновлено: 30.01.2026
Плазма составляет жидкую часть крови и является водно-солевым раствором белков. Состоит на 90–95 % из воды и на 8—10 % из сухого остатка. В состав сухого остатка входят неорганические и органические вещества. К органическим относятся белки, азотосодержащие вещества небелковой природы, безазотистые органические компоненты, ферменты.
Белки составляют 7–8 % от сухого остатка (что составляет 67–75 г/л) и выполняют ряд функций. Они отличаются по строению, молекулярной массе, содержанию различных веществ. При увеличении концентрации белков возникает гиперпротеинемия, при уменьшении – гипопротеинемия, при появлении патологических белков – парапротеинемия, при изменении их соотношения – диспротеинемия. В норме в плазме присутствуют альбумины и глобулины. Их соотношение определяется белковым коэффициентом, который равняется 1,5–2,0.
Альбумины – мелкодисперсные белки, молекулярная масса которых 70 000—80 000 Д. В плазме их содержится около 50–60 %, что составляет 37–41 г/л. В организме они выполняются следующие функции:
1) являются депо аминокислот;
2) обеспечивают суспензионное свойство крови, поскольку являются гидрофильными белками и удерживают воду;
3) участвуют в поддержании коллоидных свойств за счет способности удерживать воду в кровеносном русле;
4) транспортируют гормоны, неэтерефицированные жирные кислоты, неорганические вещества и т. д.
При недостатке альбуминов возникает отек тканей (вплоть до гибели организма).
Глобулины – крупнодисперсные молекулы, молекулярная масса которых более 100 000 Д. Их концентрация колеблется в пределах 30–35 %, что составляет около 30–34 г/л. При электрофорезе глобулины распадаются на несколько видов:
За счет такого строения глобулины выполняют различные функции:
Защитная функция связана с наличием иммуноглобулинов – антител, способных связывать антигены. Также они входят в состав защитных систем организма, такие как – системы пропердина и комплемента, обеспечивая неспецифическую резистентность организма. Участвуют в процессах свертывания крови за счет наличия фибриногена, занимающего промежуточное положение между β-глобулинами и γ-глобулинами, являющимися источником фибриновых нитей. Образуют в организме систему фибринолиза, основным компонентом которой является плазминоген.
Транспортная функция связана с переносом металлов с помощью гаптоглобина и церулоплазмина. Гаптоглобин относится к β2-глобулинам и образует комплекс с трансферрином, сохраняющим для организма железо. Церулоплазмин является β2-глобулином, который способен соединять медь.
Патологические глобулины образуются в ходе воспалительных реакций, поэтому в норме не обнаруживаются. К ним относятся интерферон (образуется при внедрении вирусов), С-реактивный белок, или белок острой фазы (является β-глобулином и присутствует в плазме при тяжелых, хронических заболеваниях).
Таким образом, белки обеспечивают физико-химические свойства крови и выполняют защитную функцию.
В плазме также содержатся аминокислоты, мочевина, мочевая кислота, креатинин;
Их содержание невелико, поэтому они обозначаются как остаточный азот крови. В норме он составляет примерно 14,3—28,6 %. Уровень остаточного азота поддерживается за счет наличия белков в пище, выделительной функции почек и интенсивности белкового обмена.
Органические вещества в плазме представлены в виде продуктов обмена углеводов и липидов. Компоненты обмена углеводов:
1) глюкоза, содержание которой в норме составляет 4,44– 6,66 ммоль/л в артериальной крови и 3,33—5,55 ммоль/л в венозной и зависит от количества углеводов в пище, состояния эндокринной системы;
2) молочная кислота, содержание которой резко повышается при критических состояниях. В норме ее содержание равно 1–1,1 ммоль/л;
3) пировиноградная кислота (образуется при утилизации углеводов, в норме содержится приблизительно 80–85 ммоль/л). Продуктом липидного метаболизма является холестерин, участвующий в синтезе гормонов, желчных кислот, построении клеточной мембраны, выполняющий энергетическую функцию. В свободном виде он представлен в форме липопротеидов – комплекса белков и липидов. Выделяют пять групп:
1) хиломикроны (участвуют в транспорте триацилглицеридов экзогенного происхождения, образуются в эндоплазматической сети энтероцитов);
2) липопротеиды очень низкой плотности (переносят триацилглицериды эндогенного происхождения);
3) липопротеиды низкой плотности (доставляют холестерин к клеткам и тканям);
4) липопротеиды высокой плотности (образуют комплексы с холестерином и фосфолипидами).
Биологически активные вещества и ферменты относятся к группе веществ, обладающих высокой энзимной активностью, на их долю приходится 0,1 % сухого остатка.
Неорганические вещества являются электролитами, т. е. анионами и катионами. Они выполняют ряд функций:
1) регулируют осмотическое давление;
2) поддерживают pH крови;
3) участвуют в возбуждении клеточной мембраны.
У каждого элемента имеются свои функции:
1) йод необходим для синтеза гормонов щитовидной железы;
2) железо входит в состав гемоглобина;
3) медь катализирует эритропоэз.
Осмотическое давление крови обеспечивается за счет концентрации в крови осмотически активных веществ, т. е. это разность давлений между электролитами и неэлектролитами.
Осмотическое давление относится к жестким константам, его величина 7,3–8,1 атм. Электролиты создают до 90–96 % всей величины осмотического давления, из них 60 % – хлорид натрия, так как электролиты имеют низкую молекулярную массу и создают высокую молекулярную концентрацию. Неэлектролиты составляют 4—10 % величины осмотического давления и обладают высокой молекулярной массой, поэтому создают низкую осмотическую концентрацию. К ним относятся глюкоза, липиды, белки плазмы крови. Осмотическое давление, создаваемое белками, называется онкотическим. С его помощью форменные элементы поддерживаются во взвешенном состоянии в кровеносном русле. Для поддержания нормальной жизнедеятельности необходимо, чтобы величина осмотического давления всегда была в пределах допустимой нормы.
Запасание аминокислот. Физиология белков плазмы крови
а) Запасание аминокислот в виде белков в клетках. Практически сразу после поступления в клетки тканей аминокислоты связываются друг с другом пептидными связями благодаря непосредственному влиянию матричной РНК и рибосомальных систем и образуют белки, поэтому концентрация аминокислот в клетках остается низкой. Кроме этого, свободные аминокислоты никогда не запасаются в клетках, их хранение возможно только в виде белков. Многие из этих внутриклеточных белков могут вновь быстро преобразовываться в аминокислоты под влиянием внутриклеточных лизосомальных пищеварительных ферментов. Появляющиеся при этом аминокислоты поступают в кровь. Исключение составляют только белки, присутствующие в ядре клетки и на хромосомах, и некоторые структурные белки (например, белки коллагеновых волокон и сократительные белки мышц). Эти белки не принимают участия в процессах, приводящих к выходу из клеток аминокислот, составляющих белки, после их переваривания.
Некоторые ткани способны участвовать в запасании аминокислот в большей степени, чем другие. Так, печень — крупный орган, имеющий особые системы для обработки аминокислот, который может запасать большие количества быстрообмениваемых белков. Это также свойственно, хотя и в меньшей степени, почкам и слизистой кишечника.
б) Высвобождение аминокислот из клеток как способ регуляции концентрации аминокислот в плазме. В случаях падения концентрации аминокислот в плазме крови до слишком низкого уровня необходимые аминокислоты могут транспортироваться из клеток для восполнения возникающего дефицита аминокислот в плазме. Таким образом, концентрация конкретных аминокислот в плазме крови поддерживается на достаточно постоянном уровне. Следует отметить, что некоторые гормоны, секретируемые железами внутренней секреции, способны смещать баланс между белками тканей и циркулирующими в крови аминокислотами. Так, гормон роста и инсулин увеличивают образование белков в тканях, в то время как глюкокортикоиды коры надпочечника повышают концентрацию аминокислот в плазме крови.
в) Подвижное равновесие между белками различных частей тела. Вследствие того, что внутриклеточные белки в печени (а также в других тканях, но в несколько меньшей степени) могут быстро синтезироваться из аминокислот плазмы крови и столь же быстро распадаться до возвращающихся в плазму аминокислот, практически во всех клетках организма поддерживается постоянный обмен и равновесие между аминокислотами плазмы крови и мобильными белками.
Например, если возникает потребность в белках в какой-то конкретной ткани, она может синтезировать белки из аминокислот крови. В свою очередь, белки плазмы восполняются за счет распада белков в других клетках организма, особенно белков печени. Эта возможность особенно заметна на примере синтеза белков раковыми клетками. Раковые клетки потребляют аминокислоты в больших количествах, что приводит к заметному опустошению белковых компонентов в других клетках.
г) Верхний предел запасания белков. Каждый конкретный вид клеток характеризуется собственным верхним пределом количества запасаемого белка. После того как все клетки достигнут своего предела возможностей запасания белка, избыток аминокислот, циркулирующих в крови, подвергается распаду до каких-то иных веществ или используется на энергетические нужды либо превращается в жиры и гликоген и запасается в этой форме.
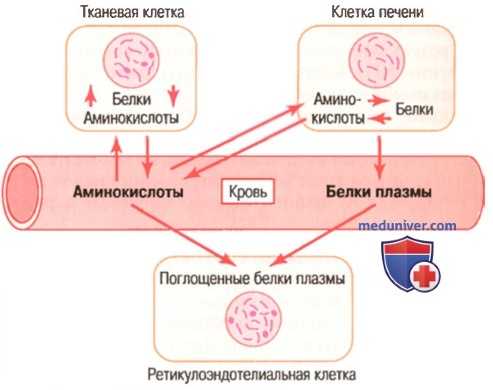
Динамическое равновесие тканевых белков, белков плазмы и аминокислот плазмы
Функциональное предназначение белков плазмы крови
Белки плазмы крови подразделяют на три большие группы: альбумины, глобулиныи фибриноген.
Основной функцией альбуминов является создание коллоидно-осмотического давления плазмы крови, которое препятствует потерям плазмы в капиллярах.
Глобулины обеспечивают многие ферментные функции плазмы и, что не менее важно, отвечают за врожденный и приобретенный иммунитет.
Фибриноген во время свертывания крови полимеризуется в длинные нити фибрина, что служит причиной образования сгустка крови, помогающего восстановлению герметичности системы кровообращения.
а) Образование белков плазмы крови. Практически все альбумины и фибриноген плазмы крови наряду с 50-80% глобулинов образуются в печени. Остальные глобулины образуются в лимфоидной ткани. В большинстве своем это гамма-глобулины, представляющие собой антитела иммунной системы.
Скорость образования белков в печени чрезвычайно высока — более 30 г/сут. Некоторые заболевания приводят к быстрому снижению количества белков в плазме крови. Тяжелые ожоги, сопровождающиеся разрушением большой площади поверхности кожи, могут вызвать ежедневную потерю нескольких литров плазмы через разрушенную поверхность. Высокая скорость образования белков плазмы крови печенью в состоянии предупредить в таких случаях смертельный исход. Иногда тяжелые заболевания почек приводят к ежедневным потерям более 20 г белков плазмы крови с мочой на протяжении многих месяцев, и в течение всего этого времени печень оказывается в состоянии восполнять утрачиваемый белок, продуцируя необходимое его количество.
При циррозах печени в паренхиме печени разрастается соединительная ткань, что сопровождается снижением синтеза белков. Это приводит к снижению коллоидно-осмотического давления плазмы крови и развитию генерализованных отеков.
б) Белки плазмы крови как источник аминокислот для тканей. Если белковые компоненты тканей оказываются израсходованными, белки плазмы крови могут послужить источником для быстрого их восстановления. Путем пиноцитоза белки плазмы крови могут целиком поглощаться тканевыми макрофагами. Попав в эти клетки, белки расщепляются до аминокислот, которые затем вновь поступают в кровь и используются всеми клетками организма для образования белков там, где это необходимо. Таким способом белки плазмы крови используются в качестве источника быстрого поступления белка, содержащего аминокислоты, готовые для использования в тканях, нуждающихся в белке.
в) Динамическое равновесие между белками крови и белками тканей. Между белками плазмы, аминокислотами плазмы и белками тканей постоянно существует состояние равновесия (для облегчения понимания просим вас изучить рисунок выше). В исследованиях с использованием радиоактивно меченных атомов было установлено, что в норме ежедневно синтезируются и распадаются около 400 г белка. Это служит проявлением существующего постоянного обмена аминокислот и демонстрирует правило взаимообмена аминокислотами среди различных белков организма. Даже во время голодания или тяжелых истощающих заболеваний отношение общего количества белков тканей к общему количеству белков плазмы в организме остается относительно постоянным, составляя приблизительно 33:1.
В связи с существованием такого динамического равновесия между белками плазмы и прочими белками тела эффективным способом лечения тяжелых острых состояний дефицита белка в организме может быть внутривенное введение белков плазмы крови. Через несколько дней, а иногда и часов аминокислоты введенных белков распределяются среди клеток организма для образования новых белков там, где это необходимо.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Обмен белков и аминокислот в организме
В последние десятилетия были предприняты значительные усилия, направленные на понимание регуляции белкового обмена у новорожденных. Эта область исследования является наиболее актуальной, поскольку на протяжении того же периода количество случаев преждевременного родоразрешения в США значительно увеличилось. В 2005 г. гестационный возраст 12,5% детей, родившихся в США, составлял менее 37 нед (повышение на 31% по сравнению с 1981 г.).
Повышенная частота преждевременных родов и очень низкая масса тела детей при рождении создают многочисленную популяцию новорожденных, которым необходима дополнительная нутритивная поддержка. Большинство неонатологов согласятся с тем, что, хотя усовершенствование процесса перинатальной помощи увеличило выживаемость, современные нормы питания для многих недоношенных детей и детей с ОНМТ не являются оптимальными.
Итак, существует необходимость в дальнейшем изучении проблемы, каким образом с помощью нутритивной поддержки можно обеспечить максимальный рост новорожденных.
Технологический прогресс, достигнутый благодаря использованию меченных изотопами аминокислот в качестве индикаторов и основным достижениям молекулярной биологии, начал проливать свет на механизм и клиническое воздействие аминокислот на интенсификацию роста и увеличение объема белков в растущем организме новорожденных.
Данные, полученные в результате этих исследований, показывают, что текущие рекомендации по употреблению в пищу белков и аминокислот могут быть неадекватными в плане обеспечения максимального роста и увеличения объема белков у младенцев, родившихся преждевременно.

В задачи дальнейших статей на сайте входит, во-первых, провести обзор основных представлений о белковом обмене, синтезе и кругообороте белка, уделяя больше внимания потребностям новорожденных, необходимым для их роста. Во-вторых, сделать обзор текущих рекомендаций по кормлению недоношенных детей, уделяя особое внимание потреблению белка.
Процесс, при котором белки организма непрерывно разрушаются и ресинтезируются, называют кругооборотом белка. Этот термин, используемый в собирательном значении, обозначает как синтез белка, так и его распад. В дополнение к обмену аминокислот, который происходит при образовании и распаде белков, аминокислоты также необратимо утрачиваются, распадаясь в процессе метаболизма. В организме, в котором сохраняется белковый баланс, количество аминокислот, утраченных в результате распада, эквивалентно количеству аминокислот, полученных с пищей.
Деградация белков заключается в удалении азота, главным образом в виде мочевины и аммиака, а также в расщеплении оставшихся углеродов, относящихся к углеродному скелету. Конечным результатом деградации углеродного скелета является обеспечение организма энергией либо непосредственно, либо через формирование простых соединений, например глюкозы и жирных кислот, которые затем могут быть сохранены или метаболизированы для получения энергии. Потребности организма регулируют интенсивность подачи (потока) аминокислот с помощью метаболических путей.
Энергетический баланс и баланс азота влияют на то, используются ли для синтеза аминокислот и/или углеводов аминокислоты и их углеродный скелет или же они окисляются для выработки энергии. Следует отметить, что, если бы продукты, образовавшиеся в результате распада содержащихся в организме белков, были на 100% утилизированы повторно для образования 20 классических аминокислот, необходимость потреблять белок в рационе сводилась бы к минимуму. Тем не менее отдельные аминокислоты не могут быть синтезированы в человеческом организме даже при наличии достаточного количества азота. Эти аминокислоты называют незаменимыми (основными) аминокислотами.
Аминокислоты в организме также преобразуются в конечные небелковые продукты. Небелковые производные включают в себя такие соединения, как пуриновые и пиримидиновые основания, медиаторы (например, серотонин), а также непептидные гормоны (например, катехоламины). Количество аминокислот, участвующих в этих небелковых путях превращения, гораздо меньше общего количества аминокислот, участвующих в синтезе белков и их распаде.
Поскольку аминокислот, необратимо используемых для синтеза небелковых соединений, как правило, гораздо меньше, чем тех аминокислот, которые были использованы либо для синтеза белков, либо для окисления аминокислот, при оценке кругооборота белка и баланса азота эти пути часто игнорируют. Тем не менее количество некоторых из этих синтезированных соединений может быть значительным (например, при образовании гема, нуклеиновых кислот), поэтому уменьшение числа этих соединений может стать значимым для некоторых аминокислот в продолжительные периоды недостаточного потребления белков.
Аминокислоты крови. Пути поступления аминокислот
а) Аминокислоты крови. Нормальная концентрация аминокислот в крови колеблется от 35 до 65 мг/дл. Это соответствует в среднем 2 мг/дл для каждой из 20 аминокислот, хотя некоторые из них присутствуют в крови в существенно больших количествах, чем другие. В связи с тем, что аминокислоты — относительно сильные кислоты, они присутствуют в крови главным образом в ионизированном состоянии в результате отдачи одного атома водорода от группы NH2.
Количество отрицательно заряженных ионов в крови составляет 2-3 мэкв. Действительное распределение различных аминокислот в крови зависит в ряде случаев от того, какой структуры был съеденный белок, но окончательная концентрация конкретных аминокислот регулируется их избирательным синтезом в различных клетках.
б) Аминокислоты, всасываемые из желудочно-кишечного тракта. Продукты переваривания и всасывания белков в желудочно-кишечном тракте представлены в основном аминокислотами. Очень редко полипептиды и целые молекулы белка поступают из желудочно-кишечного тракта в кровь. Сразу после еды концентрация аминокислот в крови повышается, но это увеличение в норме составляет всего несколько миллиграммов на децилитр, что объясняется двумя причинами:
(1) для переваривания и всасывания аминокислот требуется 2-3 ч, поэтому только небольшое количество аминокислот может всасываться из кишечника сразу после еды;
(2) сразу после появления в крови избытка аминокислот в течение 5-10 мин они абсорбируются всеми клетками организма, особенно клетками печени.
В силу этого практически никогда не создается избыточно высокой концентрации аминокислот в крови, они не накапливаются в плазме крови или тканевой жидкости. Более того, скорость кругооборота аминокислот так высока, что основная масса белка может переноситься из одной части организма в другую в виде аминокислот в течение 1 ч.
в) Активный транспорт аминокислот в клетки. Молекулы аминокислот слишком велики для того, чтобы с легкостью диффундировать через поры в клеточных мембранах, поэтому значительные количества аминокислот могут перемещаться как внутрь, так и наружу через мембраны только путем облегченной диффузии или активного транспорта с использованием переносчиков. Природа транспортных механизмов изучена слабо, но о некоторых из них говорилось в отдельной статье на сайте (просим вас пользоваться формой поиска выше).
г) Почечный порог для аминокислот. В почках различные аминокислоты могут активно реабсорбироваться эпителием проксимальных канальцев, который извлекает их из гломерулярного фильтрата и возвращает обратно в кровь, если они смогли профильтроваться в почечные канальцы через мембраны гломерулярного аппарата. Однако существование активного транспортного механизма в канальцевом аппарате предопределяет наличие предела скорости, с которой может транспортироваться каждый вид аминокислот, поэтому в случаях, когда концентрация какой-либо определенной аминокислоты становится слишком высокой в плазме и гломерулярном фильтрате, ее избыток, который не в состоянии реабсорбировать механизм активного транспорта, теряется с мочой.
Механизмы белкового (азотистого) баланса
Для упрощения понимания процесса все тканевые и циркулирующие белки рассматривают вместе: более того, все свободные аминокислоты упрощают до единого однородного пула, а не рассматривают в сложных взаимодействиях в крови, тканях и внутриклеточных компартментах.
Это упрощение оказалось полезным при разработке концепции и совершенствовании методов измерения обмена аминокислот в свободном и связанном состояниях. Обмен свободных аминокислот с белками организма происходит в ходе процессов синтеза белка и его распада, а также на протяжении всего существования аминокислот от поступления их с пищей до исчезновения при окислении.
Незаменимые свободные аминокислоты поступают в пул организма после переваривания и всасывания белков пищи, а также в результате распада белков организма. Удаление свободных аминокислот из пула происходит либо при синтезе белка, либо посредством их экскреции путем окисления до СО2 и выделения сопутствующих метаболитов азота, преимущественно аммиака и мочевины.
Если количество свободных аминокислот в пуле постоянно, то сумма процессов, удаляющих аминокислоты (синтез белка плюс окисление) равна сумме процессов, посредством которых аминокислоты поступают в свободный пул (деградация белков плюс поступление аминокислот с пищей). Это можно выразить с помощью следующей формулы: S+E=D+I.
При азотистом равновесии потребление азота (I) эквивалентно экскреции азота (Е) и синтез белка (S) равен распаду белка (D). Чтобы происходил рост клеток, необходимо добиться положительного баланса азота, а следовательно, преобладания синтеза белков или их аккреции (S > D). Верно и обратное: для достижения отрицательного баланса азота должны иметь место преимущественный распад белка или его потеря (S < D).
Из описанных соотношений становится ясно, что белок удерживается в организме, когда синтез преобладает над распадом, и организм утрачивает белок, когда распад преобладает над синтезом. В отличие от метода оценки баланса азота, который измеряет только «результирующие» изменения в содержании белков организма, оценка синтеза белка и его распада показывает, что изменение баланса происходит различными путями. Например, потеря белка в организме возможна:
(1) из-за снижения интенсивности синтеза белка без изменения темпов его распада;
(2) из-за увеличения темпов распада без снижения интенсивности синтеза белка;
(3) вследствие изменений, происходящих в обоих процессах, в силу чего один из них доминирует над другим.
При некоторых патологических или стрессовых состояниях распад белка преобладает над синтезом, причем скорость как синтеза белка, так и его распада выше, чем у здоровых лиц. Положительный баланс белка может быть достигнут за счет интенсификации синтеза белка, уменьшения распада или изменений и в синтезе белка, и в его распаде, при которых синтез доминировал бы над распадом.
Например, у выздоравливающих от гипотрофии детей скорость синтеза и распада белка увеличивается, но интенсификация синтеза в большей степени, чем усиление распада, дает положительный баланс белка. Таким образом, измерения синтеза и распада белка обеспечивают информацией о том, какого рода изменения происходят в белковом балансе.
Стоит отметить, что, хотя упомянутое объяснение базируется на содержании белка во всем организме, концепция равновесия между процессами синтеза и распада также строится на уровне отдельных тканей или органов и для отдельных белков.
Читайте также: