
Строение рефлекторной дуги симпатической нервной системы
СОМАТИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
иннервирует скелетную мускулатуру
центры находятся в передних рогах спинного мозга
рефлекторная дуга состоит как минимум из 2 нейронов:
I нейрон - чувствительный, его перикарион лежит в спинномозговом ганглии, длинный дендрит отходит на периферию, где заканчивается рецептором, аксон входит в задние рога спинного мозга, проходит в передний рог (или переключается на ассоциативный нейрон) и образует синапс со II нейроном; I нейрон - пурин-пептидергический, нейромедиаторы - АТФ, субстанция Р, кальцитонин ген-родственный пептид
II нейрон - двигательный или эфферентный, его перикарион лежит в передних рогах спинного мозга, аксон через передние рога выходит из спинного мозга и идет к скелетной мышце, где образуется аксо- мышечный синапс; II нейрон - холинергический, нейромедиатор - ацетилхолин, на постсинаптической мембране (т.е. на мембране мышечного волокна) имеются Н-холинорецепторы скелетных мышц
ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
иннервирует все внутренние органы, сердце и сосуды, экзокринные и эндокринные железы, органы чувств
подразделяется на 2 отдела - симпатический и парасимпатический
каждый орган, как правило, получает и симпатическую, и парасимпатичеcкую иннервацию
СИМПАТИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
центры находятся в боковых рогах грудного и поясничного отделов спинного мозга
рефлекторная дуга состоит как минимум из 3 нейронов:
I нейрон - чувствительный, его перикарион лежит в спинномозговом ганглии, длинный дендрит отходит на периферию, где заканчивается рецептором, аксон входит в задние рога спинного мозга, проходит в боковой рог (или переключается на ассоциативный нейрон) и образует синапс со II нейроном; I нейрон - пурин-пептидергический, нейромедиаторы - АТФ, субстанция Р, кальцитонин ген-родственный пептид
II нейрон - называется преганглионарным; эфферентный, его перикарион и дендриты лежат в боковых рогах спинного мозга, аксон через передние рога выходит из спинного мозга и идет к симпатическому ганглию,где образует синапсы с III нейроном;II нейрон- холинергический,нейромедиатор - ацетилхолин
III нейрон - называется постганглионарным; эфферентный, его перикарион и дендриты лежат в симпатических ганглиях (пре- и паравертебральные ганглии, ); на перикарионе и дендритах III нейрона имеются Н-холинорецепторы, через которые происходит синаптическая передача между II (преганглионарным) и III(постганглионарным) нейронами (ацетилхолин высвобождается из пресинаптической части, принадлежащей I нейрону и взаимодействует с Н-холинорецепторами, находящимися на постсинаптической мембране, то есть на мембране III нейрона); аксон выходит из ганглия и идет к иннервируемому органу, где образуются синаптические соединения; III нейрон - адренергический, нейромедиатор - норадреналин; синаптическая передача между III (постганглионарным) нейроном и рабочим органом осуществляется с помощью норадреналина, который высвобождается из пресинаптической части (принадлежащей III-нейрону) и взаимодействует с адренорецепторами, находящимися на постсинаптической мембране синапса, а постсинаптическая мембрана - это уже мембрана не нейрона, а органа; в различных органах имеются разные комбинации разных подтипов a-и b-адренорецепторов
ПАРАСИМПАТИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
центры находятся в боковых рогах крестцового отдела спинного мозга, продолговатом мозге и мосте (ядра III, VII, IX, X, черепномозговых нервов)
рефлекторная дуга состоит как минимум из 3 нейронов:
I нейрон - чувствительный, его перикарион лежит в спинномозговом ганглии или в толще нервного ла или в специальных чувствительных ганглиях парасимпатичесокй нервной системы (ganglion geniculi, ganglia superius et inferius, g.nodosum), или непосредственно в стволе мозга (nucleus tractus solitarii - n. vagus; nucleus sensorius principallis nervi trigemini), длинный дендрит отходит на периферию, где заканчивается рецептором, аксон входит в мозг (мост, продолговатый мозг), или в боковые рога спинного мозга (или переключается на ассоциативный нейрон) и образует синапс со II нейроном; I нейрон - пyрино-пептидергический, нейромедиаторы - АТФ, субстанция Р, кальцитонин ген-родственный пептид
II нейрон - называется преганглионарным; эфферентный, его перикарион и дендриты лежат в боковых рогах крестцового отдела спинного мозга или продолговатом мозге, мосте: ядро Edinger- Westphal - III пара черепномозговых нервов; nuclei salivatory - VII и IX черепномозговые нервы; дорсальное ядро n. vagus; nucleus ambiguus - n.vagus) аксон выходит из спинного мозга или в составе черепно-мозговых нервов идет к парасимпатическому ганглию, где образует синапсы с III нейроном; II нейрон - холинергический, нейромедиатор - ацетилхолин
III нейрон - называется постганглионарным; эфферентный, его перикарион и дендриты лежат в парасимпатических ганглиях (вегетативные ганглии черепно-мозговых нервов /g. ciliary, g.oticum, g pterygopalatinum, g. submandibulare/ или внутриорганные ганглии (интрамуральные ганглии)); на перикарионе и дендритах III нейрона имеются Н-холинорецепторы, через которые происходит синаптическая передача между II (преганглионарным) и III (постганглионарным) нейронами (ацетилхолин высвобождается из пресинаптической части, принадлежащей II нейрону и взаимодействует с Н-холинорецепторами, находящимися на постсинаптической мембране, то есть на мембране III нейрона; аксон выходит из ганглия и идет к иннервируемому органу или уже находится в органе, где образуются синаптические соединения; III нейрон - холинергический, нейромедиатор - ацетилхолин; синаптическая передача между III (постганглионарным) нейроном и рабочим органом осуществляется с помощью ацетилхолина, который высвобождается из пресинаптической части (III-нейрон) и взаимодействует с М-холинорецепторами, находящимися на постсинаптической мембране синапса, а постсинаптическая мембрана - это уже мембрана не нейрона, а органа

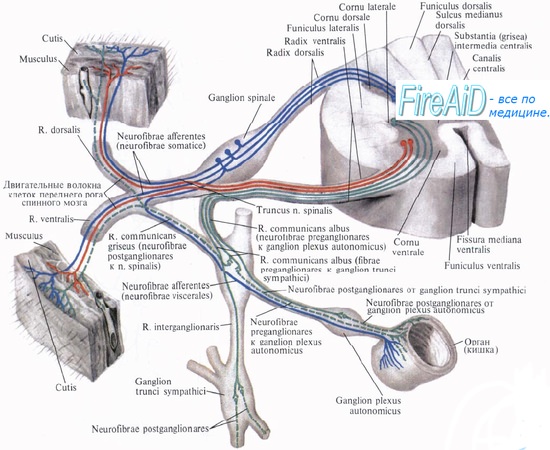
Рефлекторные дуги соматической (А), симпатического (Б) и парасимпатического (В) отделов вегетативной нервной системы
1 — чувствительный нейрон
А: 2 — двигательный нейрон
Б, В: 2 — преганглионарный нейрон
3 — постганглионарный нейрон
D1 — постганглионарный нейрон (или клетка Догеля I типа)
D2 — клетка Догеля II типа
D3 — клетка Догеля III типа (клетки Догеля — см. раздел Ганглии)
Рефлекторная дуга
Спинной мозг выполняет две важнейшие функции: рефлекторную и проводниковую.
Рефлекторная дуга – это цепь нейронов, обеспечивающих передачу возбуждения от рецепторов к рабочим органам. Она начинается с рецептора.
Рецептор – это конечное разветвление нервного волокна, которое служит для восприятия раздражения. Рецепторы всегда образуются отростками нейронов, лежащих вне мозга, в чувствительных ганглиях. Обычно в образовании рецепторов принимают участие вспомогательные структуры: эпителиальные и соединительно-тканные элементы и структуры.
Существует три вида рецепторов:
· Экстрарецепторы – воспринимают раздражение извне. Это органы чувств.
· Интрорецепторы– воспринимают раздражение из внутренней среды. Это рецепторы внутренних органов.
· Проприорецепторы– рецепторы мышц, сухожилий, суставов. Они сигнализируют о положении тела в пространстве.
Имеются простые рецепторы (болевые, например, являются просто нервными окончаниями) и очень сложные (орган зрения, слуха и так далее), имеется также множество вспомогательных структур.
Первый нейрон рефлекторной дуги – это чувствительный нейрон спинального ганглия (ganglion spinale).
Спинальный ганглий – это скопление нервных клеток в задних корешках спинномозговых нервов в межпозвоночном отверстии.
Нейроны спинального ганглия – псевдоуниполярные.Каждая такая клетка имеет один отросток, который очень быстро делится на два – периферический и центральный отростки.
Периферические отростки идут на периферию тела и образуют там своими конечными разветвлениями рецепторы. Центральные же отростки ведут в спинной мозг.
В простейшем случае центральный отросток клетки спинального ганглия, войдя в спинной мозг, образует синапс непосредственно с двигательными (мотонейронами) и вегетативными (боковые рога) нейронами. Аксоны этих нейронов выходят из спинного мозга в составе вентрального корешка (radis ventralis) спинномозговых нервов и идут к эффекторам. Двигательный аксон идёт к поперечно-полосатым мышцам, а вегетативный – к вегетативному ганглию. От вегетативного ганглия волокна направляются к железам и гладким мышцам внутренних органов.
Таким образом, железы, гладкие мышцы и поперечно-полосатые мышцы – это эффекторы, которые отвечают за раздражение.
На одно и то же раздражение возможен ответ со стороны как двигательных, так и вегетативных центров. Например, сухожильный коленный рефлекс. Но даже в самых простейших реакциях участвует не один сегмент спинного мозга, а несколько, и, чаще всего, головной мозг, поэтому необходимо, чтобы импульс распространялся по всему спинному мозгу и доходил до головного. Вот это осуществляется с помощью вставочных клеток (интернейронов) задних рогов серого вещества спинного мозга.
Как правило, между чувствительным нейроном спинального ганглия и мотонейроном переднего рога серого вещества спинного мозга вставлен переключательный нейрон заднего рога. Центральный отросток клетки спинального ганглия объединяет синапс со вставочной клеткой. Аксон этой клетки выходит и Т-образно делится на восходящий и нисходящий отростки. От этих отростков отходят боковые отростки (коллатерали) к разным сегментам спинного мозга и образуют синапсы с двигательными и вегетативными нервами. Так импульс и распространяется по спинному мозгу.
Аксоны переключательных нейронов идут к другим сегментам спинного мозга, где синаптируют с мотонейронами, а также переключательным ядрам головного мозга. Аксоны переключательных нейронов образуют собственные пучки спинного мозга и большинство восходящих проводящих путей. Поэтому принято говорить о рефлекторном кольце, так как в эффекторах имеются рецепторы, которые постоянно посылают импульсы в ЦНС.
Вставочные клетки есть и в передних рогах. Они распределяют импульс по различным мотонейронам. Таким образом, всё многообразие связей в мозге обеспечивается вставочными клетками, или, иначе говоря, переключательными нейронами серого вещества спинного мозга.
Особенности рефлекторной дуги в вегетативной нервной системе
1. Тело воспринимающего нейрона в обоих случаях находится в спинальном ганглии, но воспринимающий нейрон для систем внутренних органов находится также в вегетативных ганглиях.
2. Переключательный вегетативный нейрон находится не в заднем роге серого вещества спинного мозга, как в случае соматической нервной системы, а вбоковом роге. Аксоны этих переключательных нейронов покрыты миелиновой оболочкой, т.о. это мякотные волокна. Их называют преганглионарными. Они выходят из спинного мозга в составе вентрального корешка спинномозговых нервов и идут к вегетативному ганглию.
3. Тело эффекторного вегетативного нейрона находится вне мозга, в вегетативном ганглии, тогда как двигательный нейрон лежит в мозге.
Таким образом, в вегетативных ганглиях находятся как воспринимающие, так и эффекторные нейроны. Поэтому простейшие вегетативные рефлексы могут заканчиваться на уровне вегетативных ганглиев. Это и обеспечивает некоторую автономность вегетативной нервной системы.
Спинномозговые нервы
Nervus spinalis
Тело позвоночных животных и человека на разных стадиях эмбриогенеза или эмбрионального развития сегментировано. Сегменты спинного мозга объединяются в пять отделов: шейный (8 нервов, 7 позвонков; 1 шейный нерв выходит между мозгом и 1 шейным позвонком), грудной(12 нервов), поясничный(5 нервов), крестцовый(4-5 нервов), копчиковый(1 нерв).
Конский хвост – cauda equina – образован корешками нижних спинномозговых нервов, которые вытягиваются в длину, чтобы достигнуть соответствующих им межпозвоночных отверстий.
Нерв – это анатомическое образование, состоящее из большого числа нервных волокон, сгруппированных в пучки, то есть в единую структуру, осуществляющую связь ЦНС со всеми органами в теле и общим кожным покровом.
Каждый нерв состоит из нервных волокон, миелинизированных и немиелинизированных, имеющих различный диаметр. В зависимости от выполняемой функции различают чувствительные, двигательные, и (преимущественно) смешанныенервы.
Чувствительныенервы сформированы отростками нейронов чувствительных черепных или спинномозговых узлов.
Двигательныенервы состоят из отростков нервных клеток, лежащих в двигательных ядрах черепных или ядрах передних стволов спинного мозга.
Вегетативные нервыобразованы отростками клеток вегетативных ядер черепных нейронов или боковых стволов спинного мозга.
Все задние корешкиспинномозговых нервов афферентные, а передние, соответственно, эфферентные.
Спинномозговые нервы иннервируют тело человека сегментарно.
Каждый спинномозговой нерв образуется от слияния передних и задних корешков сразу сбоку от спинального ганглия в межпозвоночном отверстии, через которое нерв выходит из позвоночника.
Каждый спинномозговой нерв сразу делится на четыре ветви: ramus dorsalis, ramus ventralis, ramus communicans и ramus meningeus.
Ramus dorsalis – задняя ветвь –состоит из чувствительных и двигательных волокон и иннервирует кожу и мышцы спинной части соответствующего сегмента.
Ramus ventralis – передняя ветвь –также состоит из чувствительных и двигательных волокон и иннервирует кожу и мышцы брюшной части тела.
Ramus communicans – соединительная ветвь –состоит из вегетативных волокон, которые отделяются ото всех остальных и идут к вегетативному ганглию.
Ramus meningeus – оболочечная ветвь– состоит из вегетативных и чувствительных волокон, которые возвращаются в позвоночный канал и иннервируют оболочки соответствующего сегмента мозга.
Иннервация конечностей
Конечности закладываются в онтогенезе как производные вентральной части тела, поэтому они иннервируются только вентральными ветвями спинномозговых нервов. В ходе онтогенеза конечности утрачивают следы своего сегментарного происхождения. При развитии конечностей и шеи нарушается сегментация, поэтому вентральные ветви, идущие к ним, образуют сплетения.
Сплетения –это нервные сети, в которых вентральные ветви разных спинномозговых нервов обмениваются своими волокнами. В результате из сплетений выходят нервы, каждый из которых содержит волокна от разных сегментов спинного мозга.
Различают три сплетения:
· Шейное сплетение –образовано вентральными ветвями нервов с первого по четвёртую пару шейных нервов, лежит рядом с шейными позвонками и иннервирует шею.
· Плечевое сплетение– образовано вентральными ветвями с пятого шейного по первый грудной нерв. Лежит в области ключицы и подмышечной впадины, иннервирует руку.
· Пояснично-крестцовое сплетение– образовано вентральными ветвями нервов с двенадцатого грудного по первый копчиковый. Лежит рядом с поясничными и крестцовыми позвонками и иннервирует ногу.
Рефлекторная дуга вегетативной нервной системы.
Клеточное тело воспринимающего нейрона как для анимальной, так и для вегетативной нервной системы помещается в спинномозговом узле, ganglion spinale, куда стекаются афферентные пути как от органов животной жизни, так и от органов растительной жизни и который, таким образом, является смешанным анимально-вегетативным узлом.
Клеточное тело вставочного нейрона вегетативной нервной системы в отличие от анимальной нервной системы помещается в боковых рогах спинного мозга. При этом аксон вставочного анимального нейрона, исходящий из клеток заднего рога, заканчивается в пределах спинного мозга среди клеток его передних рогов. Что же касается вставочного нейрона вегетативной нервной системы, то он в спинном мозге не заканчивается, а выходит за его пределы, к нервным узлам, расположенным на периферии.
Выйдя из спинного мозга, аксон вставочного нейрона подходит или к узлам симпатического ствола, ganglia trunci sympathici, относящимся к симпатическому отделу вегетативной нервной системы (они образуют симпатический ствол), или волокна не заканчиваются в этих узлах, а направляются к предпозвоночным узлам, расположенным более периферично, между симпатическим стволом и органом (ganglia coeliaca, ganglia mesenterica).
Кроме макроскопически видимых обособленных узлов, по ходу вегетативных нервов встречаются мигрировавшие сюда в ходе эмбрионального развития небольшие группы эффекторных нейронов — микроганглии. Все волокна, идущие до узлов первого, второго или третьего порядка и являющиеся аксонами промежуточного нейрона, называются предузловыми волокнами, rami preganglionares. Они покрыты миелином.

Третий, эффекторный, нейрон анимальной рефлекторной дуги помещается в передних рогах спинного мозга, а эффекторный нейрон вегетативной рефлекторной дуги вынесен в процессе развития из центральной нервной системы в периферическую, ближе к рабочему органу, и располагается в вегетативных нервных узлах. Из такого расположения эффекторных нейронов на периферии вытекает главный признак вегетативной нервной системы — двухнейронность эфферентного периферического пути: первый нейрон — вставочный; тело его лежит в вегетативных ядрах черепных нервов или боковых рогах спинного мозга, а нейрит идет к узлу; второй — эфферентный, тело которого лежит в узле, а нейрит достигает рабочего органа.
Эффекторные нейроны симпатических нервов начинаются в ganglia trunci sympathici (узлы первого порядка) или ganglia intermedia (узлы второго порядка), а для парасимпатических нервов — в около- или внутриорганных узлах, ganglia terminalia (третьего порядка); так как в названных узлах осуществляется связь вставочных и эфферентных нейронов, то отмеченная разница между симпатическим и парасимпатическим отделами вегетативной нервной системы связана именно с этими нейронами.
Аксоны эфферентных вегетативных нейронов почти лишены миелина — безмиелиновые (серые). Они составляют послеузловые волокна, rami postganglionics. Послеузловые волокна симпатической нервной системы, отходящие от узлов симпатического ствола, расходятся в двух направлениях. Одни волокна идут к внутренностям и составляют висцеральную часть симпатической системы. Другие волокна образуют rami communicantes grisei, соединяющие симпатический ствол с анимальными нервами.
В составе последних волокна достигают соматических органов (аппарата движения и кожи), в которых иннервируют непроизвольную мускулатуру сосудов и кожи, а также железы.
Совокупность описанных эфферентных вегетативных волокон, идущих от узлов симпатического ствола до органов сомы, составляет соматическую часть симпатического отдела. Такая структура обеспечивает функцию вегетативной нервной системы, которая регулирует обмен веществ всех частей организма применительно к непрерывно изменяющимся условиям среды и условиям функционирования (работы) тех или иных органов и тканей.
Соответственно этой наиболее универсальной своей функции, связанной не с какими-либо отдельными органами и системами, а со всеми частями, со всеми органами и тканями организма, вегетативная нервная система и морфологически характеризуется универсальным, повсеместным распространением в организме.
Следовательно, симпатический отдел иннервирует не только внутренности, но и сому, обеспечивая в ней обменные и трофические процессы.
В результате каждый орган, по И. П. Павлову, находится под тройным нервным контролем, в связи с чем он различает три вида нервов:
1) функциональные, осуществляющие функцию данного органа;
2) сосудодвигательные, обеспечивающие доставку крови к органу, и
3) трофические, регулирующие усвоение из доставленной крови питательных веществ.
Висцеральная часть симпатического отдела содержит все эти три вида нервов для внутренностей, а соматическая часть — только сосудодвигательные и трофические. Что же касается функциональных нервов для органов сомы (скелетная мускулатура и др.), то они идут в составе соматической, ани-мальной, нервной системы.
Таким образом, основное отличие эфферентной части вегетативной нервной системы от эфферентной части анимальной заключается в том, что анимальные, соматические, нервные волокна, выйдя из центральной нервной системы, идут до рабочего органа, нигде не прерываясь, тогда как вегетативные волокна на своем пути от мозга до рабочего органа прерываются в одном из узлов первого, второго или третьего порядка. Вследствие этого эфферентный путь вегетативной нервной системы разбивается на две части, из которых он и состоит: предузловые миелиновые волокна, rami preganglionares, и послеузловые, лишенные миелина (безмиелиновые) волокна, rami postganglionares.
Наличие узлов в эфферентной части рефлекторной дуги составляет характерный признак вегетативной нервной системы, отличающий ее от анимальной.
1. Чувствительные (рецепторные) нейроны
Чувствительные окончания рецепторных нейронов находятся во внутренних органах и в стенке сосудов. Локализация остальных частей данных нейронов обычна: дендриты идут в составе смешанных нервов (в т. ч. некоторых черепно-мозговых); тела нейронов находятся в спинномозговых узлах или в ганглиях, расположенных по ходу соответствующих черепно-мозговых нервов.
2. Ассоциативные (центральные) нейроны
а) Тела ассоциативных нейронов, составляющие центральный отдел симпатической системы, лежат в боковых рогах спинного мозга — на уровне от Т1,до L3 Именно отсюда происходит управление всеми теми многочисленными функциями, которые перечислялись выше.
б) Аксоны ассоциативных нейронов, образуя миелиновые волокна, идут сначала в составе передних корешков спинного мозга, а затем отходят от них (в составе т. н. белых соединительных ветвей) по направлению к симпатическому стволу — оканчиваясь в узлах этого ствола или проходя их, не прерываясь, до других симпатических ганглиев.
3. Эффекторные нейроны
а) Тела эффекторных нейронов находятся в следующих симпатических ганглиях: узлах парного симпатического ствола, узлах симпатических смешений и (что менее характерно) внутриорганных (интрамуральных) ганглиях.
б) Аксоны эффекторных нейронов, отходящие от перечисленных узлов, идут в составе постганглиопарных (безмиелиновых) волокон.
Большая часть последних направляется к внутренним органам, образуя их симпатические нервы.
Рефлекторные дуги парасимпатической нервной системы.
1. Чувствительные нейроны. в парасимпатической рефлекторной дуге тела лежат в спинномозговых узлах, связанных с некоторыми черепно-мозговыми нервами.
2. Ассоциативные (центральные) нейроны. Области местонахождения этих нейронов гораздо менее протяжены, чем центральный отдел симпатической нервной системы.
а) Тела центральных парасимпатических нейронов находятся только 1) в нескольких ядрах ствола головного мозга и 2) в промежуточной зоне ряда крестцовых и копчиковых сегментов спинного мозга
б) Аксоны этих нейронов образуют преганглионарные (миелиновые) волокна. Из ствола мозга эти волокна идут в составе ряда черепно-мозговых нервов; наиболее важными являются блуждающие нервы, иннервирующие большую часть внутренних органов и сосудов.
А из нижних сегментов спинного мозга парасимпатические волокна попадают в крестцовое сплетение и иннервируют органы малого таза.
3. Эффекторные нейроны. Тела эффекторных парасимпатических нейронов обычно находятся во внутриорганных (интрамуральных) ганглиях или в ганглиях рядом с органом (чаше всего в составе соответствующих сплетений). Именно здесь оканчиваются аксоны ассоциативных нейронов У эффекторных же нейронов не только дендриты, но и аксоны— относительно короткие.
Замечание. Все вышерассмотренные рефлекторные дуги замыкаются через ЦНС и поэтому называются центростремительными.
В вегетативной нервной системе функционируют также периферические (или местные) рефлекторные дуги, замыкающиеся только через интрамуральные узлы. О них будет сказано позже.
Сенсорные системы (органы чувств): общая морфофункциональная характеристика, классификации. Особенности ультраструктуры рецепторных клеток. Гистофизиология восприятия специфического раздражения.
Совокупность структур, отвечающих за прием, передачу и анализ определенного вида раздражений, называется анализатором.
В каждом анализаторе имеются три части.
а) Периферическая часть — орган чувств (либо иной рецептор), который воспринимает специфические раздражения, поступающие из внешней или внутренней среды, и преобразует их в нервные импульсы, передаваемые в ЦНС.
б) Промежуточная часть — проводящие пути и нервные ядра ЦНС, включенные в передачу сигнала.
в) Центральная часть — определенный участок коры больших полушарий, осуществляющий обработку и анализ сигнала.
В результате деятельности анализаторов ЦНС получает информацию о внешнем мире и состоянии самого организма.
Классификация органов чувств
По природе и организации рецепторного аппарата органы чувств и прочие рецепторы подразделяют на три типа.
а) Первично чувствующие органы чувств: рецепцию в них осуществляют специализированные нервные (нейросенсорные) клетки, находящиеся в органе чувств. Сюда относятся органы зрения и обоняния.
б) Вторично чувствующие органы чувств: раздражение воспринимают специализированные эпителиальные (эпителиосенсорные, или сенсоэпителиальные) клетки, находящиеся в органе чувств. Таковы органы вкуса, слуха и равновесия.
в) Рецепторы, не организованные в органы чувств. Это, во-первых, рецепторы кожи, мышц и внутренних органов. Информация от них по обычным афферентным путям поступает в ЦНС и может достигать коры больших полушарий.
Но имеются рецепторы и другой природы — например, осморецепторы почек и гипоталамуса. Они не входят в тот или иной анализатор, и поэтому их раздражение сознанием человека не воспринимается.
Вкусовая и обонятельная сенсорные системы: тканевые компоненты, источники развития, особенности строения, физиологическая регенерация. Гистофизиология хеморецепции.
Орган вкуса — это совокупность вкусовых почек, расположенных в эпителии боковых стенок многих сосочков языка — листовидных, грибовидных, желобоватых
Листовидные сосочки покрыты многослойным плоским неороговсваюшим эпителием. Между сосочками имеются узкие просветы, выстланные таким же эпителием. С этими просветами контактируют вкусовые почки. Последние находятся в толще покровного эпителия (выстилающего просветы), но отличаются более светлой окраской и имеют округлую или овальную форму.
В эмбриогенезе вкусовые почки развиваются из многослойного эпителия сосочков.
Вкусовая почка включает 30-60 клеток, которые подразделяются на следующие виды.
а) Рецепторные (сенсорные) вкусовые эпителиоциты — имеют темное веретенообразное ядро, находящееся в почке на уровне верхнего ряда ядер.
б) Поддерживающие эпителиоциты отличаются круглым ядром (на уровне среднего ряда ядер) и часто вакуолизированной цитоплазмой.
в) Назальные эпителиоциты - вместе со своими ядрами расположены у основания ночки.
г) Перигеммальные, или периферические, клетки —- расположены на периферии почки.
Возбуждение вкусовых рецепторов на микроворсинках изменяет потенциал клеток. Видимо, это происходит опосредованно:
а) связывание вкусовых веществ с мембранными рецепторами изменяет активность ряда соседних мембранных белков с ферментативной активностью;
б) поэтому в клетке меняется концентрация определенных внутриклеточных медиаторов (таких, как цАМФ);
в) а это, в свою очередь, сказывается на состоянии ионных каналов в плазмолемме, что и меняет потенциал клетки.
Затем возбуждение через синапсы передается афферентным нервным окончаниям.
ОБОНЯНИЕ
Различают два органа обоняния: основной и дополнительный.
1. Основной орган обоняния — это обонятельный эпителий, покрывающий слизистую оболочку верхней части носовой полости. Его площадь — около 2 см 2 с каждой стороны.
От остальной (респираторной) части слизистой оболочки обонятельная область отличается более желтым цветом.
2. Дополнительный орган обоняния — это т. н. вомероназальныи, или якобсонов, орган: две слепые эпителиальные трубочки в нижней части перегородки носа. Обонятельные восприятия этого органа могут возбуждать половое чувство.
Эпителий основного и дополнительного органов обоняния, как и эпителий, выстилающий другие отделы воздухоносных путей, является многорядным мерцательным. Но его клеточный состав несколько иной: вместо мерцательных клеток находятся нейроны, способные к рецепции обонятельных раздражений; отсутствуют также бокаловидные (слизеобразующие) клетки.
Тем не менее в подлежащей ткани располагаются железы, которые выделяют белковослизистый секрет, покрывающий обонятельный эпителий.
Всего в составе эпителия имеются клетки 3-х видов. Нейросенсорные обонятельные клетки
и базальные эпителиоциты
Все клетки контактируют с базальной мембраной. В соответствии со своей нейральной природой эти клетки развиваются из зачатка нервной системы.
а) В сформированном органе ядросодержащие части находятся в средней части эпителия.
В этом заключается еще одно отличие: в обычном мерцательном эпителии реснитчатые клетки образуют верхний слой ядер.
б) Дендриты идут вверх: они достигают поверхности эпителия и имеют на конце утолщения — обонятельные булавы.
в) Аксоны отходят от базальной части рецепторных нейронов: они не имеют миелиновой оболочки и идут через отверстия решетчатой кости к обонятельным луковицам. В луковицах находятся тела вторых (ассоциативных) нейронов – т.н. митральных клеток. Др. клетки обонятельного эпителия. Поддерживающие эпителиоциты отделяющие обонятельные нейроны друг от друга. Базальные эпителиоциты, прилегающие к базальной мембране, они способны превращаться в поддерживающие клетки.

Читайте также: