
Определитель паразитов пресноводных рыб фауны ссср
Ранее на примерах из различных царств органического мира рассматривались отношения размеров в пределах отдельно взятых таксонов.
Например, размеры птиц СССР сравнивались с размерами млекопитающих СССР независимо от того, в каких биотопах (одинаковых или различных) обитают представители этих двух классов. В настоящей же и в следующей главах сначала будет выделен биотоп, а потом уже сопоставлены размеры тела населяющих его организмов. Причем таксоны будут характеризоваться на этот раз только теми видами, которые обитают в данном биотопе. Поскольку ранее выдерживался ход изложения от животных к низшим организмам, и сейчас вначале рассматривается биотоп, населенный только представителями фауны. Этот биотоп — среда тела рыб для обитающей в ней фауны паразитов.
Указанное число видов с известной длиной тела распределяется по 250 родам, 113 семействам, 49 отрядам и 16 классам. Наиболее значительная доля видов (28,4%) приходится на класс Monogenoidea, далее следуют классы Cnidosporidia (14,1%) и Trematoda (13,2%). Минимальные и максимальные размеры паразитов в пределах отдельных классов различаются в неодинаковой степени, что лишь частично связано с различием по числу видов. Так, максимальные различия по длине наблюдаются в классе Cnidosporidia (в 6500 раз), который по числу видов вдвое уступает классу Monogenoidea. Если иметь в виду всю паразитофауну пресноводных рыб, то размеры (длина) паразитов могут различаться в колоссальной степени — в 300 тысяч раз. Уже одна эта цифра наглядно свидетельствует о емкости и разнообразии среды обитания паразитов. Средние (средневидовые геометрические) характеристики длины для различных классов различаются гораздо меньше, всего в 450 раз.
Приведем краткие характеристики перечисленных в таблице классов, касающиеся их локализации в организме хозяина, формы тела и способа промеров. Все эти характеристики основаны исключительно на данном конкретном материале и ни в коем случае не являются общими характеристиками таксонов.
Flageilata обитают преимущественно в крови, имеют, как правило, удлиненную форму тела, длина тела измеряется от основания жгута. Длина жгута, а также ширина ундулирующей мембраны не входят в характеристику габаритов клетки и приводятся отдельно.
Sarcoditia, представленные двумя видами амеб, паразитирующих в кишечнике, имеют неправильно округлую форму, за дли-, ну клеток принимается диаметр без псевдоподий.
Sporozoa, представленные кокдидиями, являются внутриклеточными паразитами, размеры характеризуются по ооцистам, имеющим обычно неправильно шаровидную форму.
Cnidosporiclia — полостные (но не в кишечнике) и тканевые паразиты, измеряются по округлым цистам.
Haplosporidia — внутриклеточные и тканевые паразиты, измеряются по цистам, имеющим округлую форму.
Ciliata — преимущественно эктопаразиты на жабрах и коже. Имеют колоколообразную форму тела. В качестве характеристики размеров обыкновенно приводится диаметр колокола. Высота колокола, в большинстве случаев превосходящая диаметр, указывается очень редко, так же как и длина ножки у сидячих форм. В тех случаях, когда высота приводится и превышает диаметр, она принимается за характеристику максимальных линейных размеров, т. е. за длину.
Suctoria — два вида, локализуются на жабрах, тело мешковидной формы, характеризуется измерениями длины или диаметра.
Hydrozoa — единственный вид, паразитирует в оводитах, имеет неправильно шарообразную форму тела, измеряется по диаметру.
Monogenoidea — локализуются преимущественно на жаберных лепестках, имеют вытянутую, сплющенную форму тела. Характеризуются по длине и ширине, высота тела не измеряется.
Cestoidea локализуются главным образом в кишечнике. Тело, как правило, сильно удлиненное, сплющенное, измеряется длина и ширина. Высота не измеряется. Несколько видов паразитирует на рыбах только на стадии личинок, эти виды относятся к особым отрядам и в общую характеристику размеров представителей класса не включаются.
Trematoda паразитируют главным образом в кишечнике, обладают несколько удлиненным и сильно уплощенным телом, которое измеряется по длине и ширине. Высота тела не измеряется. Помимо 102 видов, которые паразитируют на рыбах во взрослом состоянии, приводятся также размеры 27 видов, для которых рыбы являются промежуточными хозяевами. Размеры этих личиночных стадий при составлении характеристик размеров класса не учитываются.
Nematoda паразитируют главным образом в кишечнике, гораздо реже в полости тела и мочевом пузыре. Имеют вытянутое, округлое в поперечном сечении тело. Размеры характеризуются по длине и диаметру поперечного сечения.
Acanthocephala локализуются в кишечнике, имеют вытянутую, приблизительно цилиндрическую форму. Измеряются по длине и ширине.
Hirudinea — эктопаразиты, имеют вытянутое, суженное на концах и уплощенное тело. Приводятся измерения длины и ширины. Высота не измеряется.
Bivalvia на стадии личинки прикрепляется к жабрам, плавникам и коже рыб. В дальнейшем обрастают эпителием и, превратившись в эндопаразитов, несколько увеличиваются в размерах. Поскольку в сводке приводятся лишь размеры глоходиев, то нами они в дальнейшем принимаются лишь в качестве эктопаразитов. Имеют характерную для двустворчатых моллюсков форму тела. Размеры характеризуются по длине и высоте.
Crustacea локализуются на жабрах и коже. Имеют часто довольно сложную и разнообразную форму тела, в поперечном сечении чаще всего (в настоящем материале) более или менее округлые, изредка уплощенные. Измеряются по длине. Для характеристики ширины нами использованы рисунки общего вида рачков, которые приводятся для всех видов за единичными исключениями.
В целом можно отметить, что промеры у самых различных групп хорошо выражают характеристики, необходимые для того, чтобы получить представление об объеме тела. То есть в промеры тела не входят различные тонкие придатки, имеющие незначительный объем. Однако отсутствие характеристик высоты тела во многих случаях затрудняет даже приблизительное, ориентировочное, определение объема. Если для форм с более или менее округлым сечением достаточно двух измерений, то для уплощенных форм их оказывается недостаточно. Рассматриваемая фауна характеризуется большим разнообразием в уплощении тела, которое часто трудно более или менее точно оценить на глаз. Тем не менее мы не отказываемся от рассмотрения приведенных линейных размеров и в тех случаях, когда мы не располагаем характеристиками высоты, приравниваем ее к ширине. Хотя такие характеристики приведенных линейных размеров часто далеко не отражают истинного объема животных, они все же представляют определенный интерес. Во-первых, это соответствует нашему намерению рассматривать в равной степени положительные и отрицательные результаты. Во-вторых, учитывая не одну лишь длину, мы все же значительно приближаемся к характеристике объема, когда используем два измерения, тем более что разнообразие по степени вытянутости формы тела у рассматриваемой фауны чрезвычайно велико. В-третьих, мы можем составить представление о тенденции различных характеристик размеров к сдвигу на шкале размеров по мере введения все более и более точных оценок объема, если не всегда о величине этого сдвига, то во всяком случае о его направлении.
Основной характеристикой размеров, имеющейся в нашем распоряжении, легко сопоставимой и известной для большинства видов, является длина тела. Обращает на себя внимание разнообразие размеров в пределах отдельных классов. Как и в Других случаях, амплитуда размеров связана с числом видов, входящих в отдельные классы. Связь эта не очень тесная. По-видимому, можно говорить лишь о слабой корреляции. Но даже и в такой формулировке из общего ряда явно выпадает класс Cftidosporidia, имеющий огромную амплитуду размеров тела, но по числу видов примерно вдвое уступающий классу Monogenoidea. Эта большая амплитуда связана с особым размерным распределением видов внутри этого класса, для которого характерно наличие двух оптимумов видообразования, широко расставленных на размерной шкале.
Нами сопоставляются кривые лишь для 12 крупнейших классов, содержащих каждый более чем по 10 видов, так как строить кривые для меньшего числа видов вряд ли имеет смысл.
Рассмотрим отношения отдельных классов в пределах типов. Мы имеем лишь два таких случая, когда в пределах типа представлен более чем одни класс с достаточным числом видов. Классы типа Protozoa не обнаруживают четкого упорядоченного расположения на размерной шкале ни по длине, ни по приведенным линейным размерам. Правда, в последнем случае расположение классов Flagellata, Sporozoa и Ciliata довольно последовательно, вершины находятся на расстоянии 0,40 единиц шкалы друг от друга. Но кривая для Cnidosporidia заметно нарушает эту последовательность. Расстояние между двумя вершинами этой кривой сопоставимо с величиной 1,50 единиц шкалы, но не согласуется с упомянутым, выше расстоянием. Отсутствие четкой упорядоченности в данном случае вряд ли может вызвать удивление. Действительно, эти четыре класса, по-видимому, очень древние и совсем разными путями приспособившиеся к паразитизму, занимают совершенно различные экологические ниши. Flagellata локализуется в кровяном русле, Sporozoa — внутриклеточные паразиты, Cnidosporidia — тканевые паразиты, наконец, Ciliata — преимущественно эктопаразиты. Возможно, здесь следует видеть в размерных отношениях лишь отдаленное отражение отношений, существовавших в эпоху их дифференциации и захвата различных сред обитания, отношений, затушеванных дальнейшей независимой эволюцией.
Классы типа Scolecida (Беклемишев, 1964 б) обнаруживают значительно более четкую упорядоченность в расположении на размерной шкале. По длине тела пять кривых образуют правильный ряд из четырех зубцов. Кривые для Nematoda и Acanthocephala практически совпадают, образуя одну группу. Среднее расстояние между четырьмя зубцами (второстепенные небольшие вершины кривой для Cestoidea мы пока не принимаем во внимание) оказывается равным 0,67 (2,0:3 = 0,67) единиц логарифмической шкалы. Если считать и крайний правый зубец, тогда среднее расстояние окажется равным 0,70 единиц.
Положение кривых на шкале приведенных линейных размеров меняется. При вычислении этих характеристик для всех животных высота тела принималась равной длине. У рассматриваемых групп относительная высота тела разнообразна в наибольшей степени, поэтому в данном случае приведенные линейные размеры определяют объем тела паразитов весьма приблизительно, и нас не должно удивлять, что упорядоченность расположения кривых здесь снижается. Важно, что она все-таки остается, хотя кривая для Nematoda резко смещается влево (для нематод характерно очень удлиненное тело) и образует единую размерную группу уже не с Acanthocephala, а с Trematoda. Среднее расстояние между оптимальными значениями размеров для различных групп классов оказывается равным уже не 0,70, а 0,57 единиц шкалы. Не приблизится ли это соотношение к 0,50 с введением характеристики высоты тела? Поскольку Cestoidea отличаются большей уплощенностью, чем Monogenoidea, этого можно ожидать. Кроме того, сильно уплощенные Trematoda должны будут также сдвинуться в сторону Nematoda и, возможно, более точно совпадут с ними по размерному распределению.
Любопытно правильное расположение классов сколецид по длине тела. Эта регулярность свидетельствует о важности именно этой характеристики размеров, а не только объема или веса. Удлинение и уплощение играли существенную роль в завоевании паразитами различных экологических ниш в пределах тела хозяев. Важность характеристики длины для паразитов, населяющих кишечный тракт, т. е. относительно узкое и длинное вместилище, не вызывает сомнения. Monogenoidea не являются эндопаразитами, хотя и имеют таковых в качестве отдельных представителей. Вхождение их в общий ряд с остальными эидопаразитическими классами свидетельствует о том, что при дифференциации отдельных ветвей сколеид отношения длин тела играли важную роль.
Попытаемся сопоставить теперь отдельные классы не по принципу ближайшего родства, а по принципу однородного биотопа. Оговоримся, что выделить однородный биотоп в пределах тела хозяев, да еще и различного таксономического положения, довольно сложно. Не менее трудно бывает отнести определенную группу паразитов к определенному биотопу внутри хозяина, поскольку представители одной и той же группы паразитов могут локализоваться иногда в самых различных органах и тканях. Мы видели уже, что внутриклеточные паразиты и паразиты кровяного русла представлены в настоящем материале специфическими и единственными классами, имеющими приблизительно симметричное размерное распределение. Следовательно, в пределах данных биотопов нечего сопоставлять. Среди тканевых паразитов класс Cnidosporidia дает два оптимума, которые, как уже упоминалось, отстоят друг от друга на приблизительно 1,50 единиц шкалы. Причем в данном случае речь идет в равной степени и о соотношении длин, и о соотношении объемов, поскольку цисты книдоспоридий имеют приблизительно шарообразную форму.
Крупные группы паразитов кишечника мы уже сравнили при сопоставлении отдельных классов сколеид. Причем была обнаружена четкая структура расположения классов на шкале длин.
Рассмотрим теперь соотношение положений кривых эктопаразитов, к которым мы относим в данном случае Ciliata, Monogenoidea, Bivalvia, Crustacea и Hirudinea. Bivalvia включены в этот перечень по следующим соображениям. Размеры их взяты по глохидиям — личинкам, которые прикрепляются к коже и жабрам рыб и, следовательно, на этом этапе являются эктопаразитами. В дальнейшем они внедряются в кожу и становятся эндопаразитами, несколько увеличиваясь в размерах. Но размерами молоди, завершающей свое паразитическое существование в рыбах, мы не располагаем, а поэтому и рассматриваем только глохидии в качестве эктопаразитов.
Соотношение положений вершин кривых распределения в зависимости от длины животных дает довольно четкую последовательность, из которой выпадает только класс Monogenoidea. Возможно, это связано с тем, что представители моногенетических сосальщиков являют собой пример наиболее далеко зашедшего приспособления к паразитизму и, кроме того, в пределах данного материала имеют узкую локализацию, главным образом на лепестках жабр. Другие же группы имеют или несколько иной или более широкий спектр локализации. Так или иначе, по длине тела положение Monogenoidea гораздо лучше согласуется с родственными эпдопаразитическими группами, чем с неродственными, но более сходными по локализации эктопаразитическими группами. Возможно, что для эктопаразитов, в отличие от эндопаразитов, отношение длин просто имеет меньшее значение.
По приведенным линейным размерам Monogenoidea уже не нарушают упорядоченности расположения остальных кривых. Расстояние между оптимумами здесь, как и в других аналогичных случаях, приближается к значению 0,50. В данном случае они в среднем равны 0,55 единиц шкалы.
Перейдем от рассмотрения классов к более крупным таксономическим группам. В пределах данного материала оказывается возможным сравнить размерное распределение отдельных подтипов сколецид. Объем типа Scolecida принимается нами по В. Н. Беклемишеву (1964). Кривые для подтипов Nemathelminthes и Acanthoccphala нам уже встречались, поскольку оба подтипа представлены единственными классами. С ними сопоставляется кривая для подтипа Plathelminthes, включающего в себя классы Monogenoidea, Trematoda и Cestoidea. Эта кривая и на шкале длин, и на шкале приведенных линейных размеров, отличается значительной положительной асимметрией. Однако, хотя можно вполне определенно указать на участки кривой, соответствующие трематодам и цестодам, отвечающие им вершины почти не выражены или выражены очень слабо. Хорошо выражена лишь одна вершина, соответствующая классу Monogenoidea.
По длине подтипы Nemathelmmthes и Acanthocephala, так же как и представляющие их классы, образуют одну размерную группу. Ей противопоставлена кривая для подтипа Plathelminthes. Расстояние между оптимумами равно 1,40 единиц логарифмической шкалы, т. е. приблизительно кратно величине 0,67 единиц, которая уже часто нам встречалась.
На шкале приведенных линейных размеров три вершины, соответствующие оптимумам числа видов, располагаются на равном расстоянии друг от друга (0,60 единиц).
Интересно сопоставить кривые для Protozoa и Metazoa, хотя в настоящее время этим группам иногда не придается таксономическое значение. Мы можем принять в данном случае точку зрения В. Н. Беклемишева (1964), который возводил эти группы в ранг подцарств. Во всяком случае простейшие и многоклеточные являют собой воплощение капитальнейших принципов строения организмов, которые любопытно сопоставить друг с другом по признаку размеров. В пределах сопоставляемого материала отношения размеров этих групп характеризуются следующим образом. Обе группы дают кривые размерного распределения с большой положительной асимметрией. Однако между ними есть и довольно отчетливое различие. Обе кривые для Protozoa имеют две хорошо выраженные вершины. У Metazoa нет такой четкой двувершинности, а кривая на шкале приведенных линейных размеров одновершинна.
Любопытно, что положения трех вершим в обоих случаях не совпадают, а чередуются таким образом, что вершина многоклеточных оказывается между двумя вершинами простейших. Однако расстояние между вершинами не остается одинаковым. На шкале длин оптимум для многоклеточных приближен к правому оптимуму простейших, отстоя от него приблизительно на 0,50 единиц шкалы. От левого же оптимума простейших его отделяет 1,00 единиц шкалы. В данном случае эти расстояния определяются довольно точно, ибо вершины кривой для простейших достаточно хорошо выражены. Отметим, что расстояние между последними оказывается равным 1,50 единиц.
На шкале приведенных линейных размеров оптимум многоклеточных оказывается расположенным как раз посредине между двумя оптимумами простейших. Расстояния в данном случае определить сложнее, потому что вершины для простейших сглажены. Если принимать их положение формально, по наивысшим точкам, то эти расстояния равны 1,00 единицам. Если положения мод находить интерполированием, то расстояния между ними можно оценить приблизительно в 0,80 единиц. Срединное положение вершины кривой для многоклеточных в обоих случаях сохраняется.
В сводке, материалом которой мы пользуемся, содержатся характеристики размеров спор двух отрядов книдоспоридий: Microsporidia и Myxosporidia. Размеры спор, за единичными исключениями, приводятся для всех видов и характеризуются весьма точно по длине, высоте и ширине. Споры книдоспоридий выбрасываются во внешнюю среду и служат для заражения ими других особей рыб. Известно, что споры этих двух отрядов имеют резко различную величину, о чем говорит название одного из отрядов.
Располагая полноценным материалом, можно оценить величину этого различия. Положения оптимумов числа видов для этих двух рядов по длине спор различаются приблизительно на 0,35 единиц шкалы. Соответствующее различие по приведенным линейным размерам равно 0,50 единиц. Таким образом, соотношение размеров спор этих главнейших отрядов споровиков в данном материале соответствует обычным отношениям размеров в обнаруженной нами структуре.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Эпштейн В.М. 1954. Некоторые особенности водного обмена пресноводных пиявок. Зоол. журн., Т. 33, вып. 3. – С. 549-555.
Эпштейн В.М. 1961. Обзор рыбьих пиявок (Hirudinea, Piscicolidae) северных морей СССР. Доклады АН СССР. Т. 141, № 6. – С. 1508-1511.
Эпштейн В.М. 1962. Обзор рыбьих пиявок (Hirudinea, Piscicolidae) Берингова, Охотского и Японского морей. Доклады АН СССР. Т. 144, № 6. – С. 1181-1184.
Эпштейн В.М. 1962. Класс Пиявки. С. 617-626. В: Определитель паразитов пресноводных рыб СССР. М.-Л.: Изд-во АН СССР. – 776 с.
Эпштейн В.М. 1963. Рыбьи пиявки пресных вод и морей СССР. Автореферат диссертации на соискание ученой степени кандидата биологических наук. Харьков: Харьковский государственный университет. – 16 с.
Эпштейн В.М. 1966. Acantobdella livanowi sp. n. – новый вид древних пиявок (Archihirudinea) из водоемов Камчатки. Доклады АН СССР, Т. 166, № 4. – С. 955-958.
Эпштейн В.М. 1967. Закономерности географического распространения морских рыбьих пиявок (Hirudinea, Piscicolidae). Зоол. журн., Т. 46, вып. 5. – С. 680-691.
Эпштейн В.М. 1987. Тип Кольчатые черви – Annelida. С. 340-372. В: Бауер О.Н. (ред.). Определитель паразитов пресноводных рыб фауны СССР. Т. 3. Ч. 2. Л.: Наука. – 583 c.
Эпштейн В.М. 1968. Класс пиявки (Hirudinea). Атлас беспозвоночных Каспийского моря. М.: Наука. - С. 113-117.
Эпштейн В.М. 1968. Класс пиявки - Hirudinea. Определитель фауны Черного и Азовского морей. Т. 1. Киев: Наукова Думка. - С. 394-405.
Эпштейн В.М. 1982. О численности пиявок в экосистемах северо-западной части Тихого океана. Гидробиол. журн., Т. 18, вып. 3. – С. 103.
Эпштейн В.М. 1984. Количественная оценка совершенства организации таксонов. Вестник зоологии. № 2. – С. 3-7.
Эпштейн В.М. 1984. Обратные связи между различными органами и центральной нервной системой в филогенезе пиявок. Эволюционные исследования. Макроэволюция. Сб. научн. трудов. Владивосток. – С. 37-48.
Эпштейн В.М. 1988. Системные исследования в таксономии и филогенетике. Современная палеонтология: Методы, направления, проблемы, практические приложения. Справочное пособие. Т. 1. М. Недра. – С. 511-525.
Эпштейн В.М. 1988. Периодизация истории таксономии и филогенетики в связи с эволюцией концепции целостности организма. Вестник зоологии. № 2. – С. 3-8.
Эпштейн В.М. 1988. Анализ и синтез при построении таксонов, их классификаций и реконструкций филогенеза. Вестник зоологии. № 6. – С. 3-7.
Эпштейн В.М. 1989. Щетинконосные, черепашьи и рыбьи пиявки мировой фауны (системный подход к классификации и филогении).Рыбьи пиявки пресных вод и морей СССР. Автореферат диссертации на соискание ученой степени доктора биологических наук. Ленинград: Зоологический институт АН СССР. – 39 с.
Эпштейн В.М. 1993. Методологические основы гуманитаризации биологического образования: учебное пособие. Киев: Изд-во ИСИОУ. - 76 с.
Эпштейн В.М., Грешнер М.Ю. 1997. Лекции по истории биологии. 19 век. Харьков: РИП "Оригинал. - 240 с.
Эпштейн В.М. 2003. Философия систематики. Книга вторая. Принципы построения теории систематики и проблемы целостности организма в истории биологии. М.: Изд-во КМК. – 352 с.
Эпштейн В.М. 2002. Философия систематики. Книга третья. Современные проблемы теории систематики. М.: Изд-во КМК. – 235 с.
Эпштейн В.М. 2004. Философия систематики. Книга четвертая. Теоретическая систематика. Часть первая. Идиографическая систематика. Утверждения и комментарии. Донецк: Норд-пресс. – 432 с.
Epshtein V. M. 2004. On the Origin of the Hirudinea Fauna, Especially Piscicolidae, in Ancient Lakes. Lauterbornia. Zeitschrift fur Fasnistik und Floristik des Susswassers. Heft 52. (Hirudinea: 2.Internationale Fachtagung 03 bis 06 Oktober 2003 in Leipzig, Germany) - P. 181-193.
Yeyger G.V., Epshtein V.M. 2005. Biolinguistics – Status, Problems and Solutions in Connection with General Theory of Developing Systems (GTDS). Proceeding of WOSC 13th International Congress of Cybernetics and Systems. – Vol. 8. Natural Systems and Plenary Session. – P. 31-38.
Эпштейн В.М. 2007. Памяти Я.И. Старобогатова // Моллюски. Морфология, таксономия, филогения, биогеография и экология. Сборник научных работ по материалам Седьмого (VII) совещания по изучению моллюсков (Санкт-Петербург, 14-17 ноября 2006 г.). Под ред. Егоровой Э.Н., Кияшко П.В., Сиренко Б.И. - СПб.: Изд-во ЗИН РАН. - С. 17-20.
Эпштейн В.М. 2009. Философия систематики. Книга пятая. Теоретическая систематика. Часть вторая. Номотетическая систематика. Утверждения и комментарии. Гельзенкирхен: Изд-во Edita Gelsen. – 213 с.
Эпштейн В.М. 2009. Версия современной теории эволюционной систематики. Труды Зоологического института РАН, Приложение 1. Вид и видообразование. Анализ новых взглядов и тенденций. М.: Товарищество научных изданий КМК. - С. 272-293.
Эпштейн В.М. 2011. Философия систематики. Книга шестая. Общая теория развивающихся систем (прогресс в живой природе и обществе). Утверждения и комментарии. Гельзенкирхен: Изд-во Edita Gelsen. – 182 с.
Эпштейн В.М. 2012. Исследования морских паразитических пиявок (1954-2012 гг.): итоги персональной исследовательской программы. Материалы V Всероссийской конференции с международным участием по теоретической и морской паразитологии (23-27 апреля 2012 г., Светлогорск, Калининградская обл.). Нигматуллин Ч.М. (ред.). Калининград: Изд-во АтлантНИРО. – С. 225-228.
Эпштейн В.М. 2012 MS. Философия систематики. Книга седьмая. Теоретическая систематика. Часть третья. Филогенетическая кибернетика. Утверждения и комментарии. Незаконченный черновик рукописи (ее электронный вариант хранится у дочери автора - Л.В. Эпштейн).
Эпштейн В.М. 2013. Философия систематики. Книга восьмая. Щетинконосные, черепашьи и рыбьи пиявки мировой фауны. Часть первая. Внешний вид пиявок Acantobdellidae, Paracanthobdellidae, Ozobranchidae, Piscicolidae Палеарктики. Гельзенкирхен: Изд-во Edita Gelsen. – 65 с.
Эпштейн В.М. 2014. Сага о Соломоне Самуиловиче // От них исходило душевное тепло: к 95-летию со дня рождения доктора биологичсеких наук С.С. Шульмана и кандидата биологических наук Р.Е. Шульман-Альбова. Сост. Л.С. Шульман, Б.С. Шульман. – Минск: Изд-во Медисонт. - С. 169-177.
Не нашли то, что искали? Воспользуйтесь поиском:
Определитель составлен по материалам книги Е. А. Веселова "Определитель пресноводных рыб фауны СССР"; Москва, "Просвещение", 1977 г.


Рис. 4. Осетровые, сельдевые и лососевые: 1 - амударьинский большой лопатонос (Pseudoscaphirhynchus kaufmanni); 2 - каспийский пузанок (Caspialosa caspia); 3 - азовский пузанок (C. tanaica); 4 - тюлька (Clupeonella delicatula); 5 - сима (Oncorhynchus masu), самка; 6 - сима, самец.

Рис. 5. Лососевые: 1- кижуч (Oncorhynchus kisutch), самка; 2 - кижуч, самец; 3 - кета (O. keta), самка; 4 - кета, самец; 5 - чавыча (O. tschawytscha).

Рис. 6. Лососевые: 1 - красная, или нерка (Oncorhynchus nerka) самка; 2 - нерка самец; 3 - горбуша (O. gorbuscha), самец; 4 - горбуша, самка.

Рис. 7. Лососевые: 1 - лосось семга (Salmo salar); 2 - лосось кумжа (S. trutta); 3 - ручьевая форель (S. t. morpha fario); 4 - озерная форель (S. t. m. lacustris).

Рис. 8. Лососевые: 1 - севанская форель ишхан (S. ischchan); 2 - севанская форель, летний бахтак (S. i. infraspecies aestivalis); 3 - голец (Salvelinus alpinus); 4 - мальма, или тихоокеанский голец (S. malma); 5 - палья (S. lepechini).

Рис. 9. Лососевые: 1 - кунджа (Salvelinus leucomainis); 2 - таймень (Hucho taimen); 3 - ленок (Brachymystax lenok).

Рис. 10. Лососевые: 1 - нельма (Stenodus leucichthys nelma); 2 - белорыбица (S. leucichthys); 3 - европейская ряпушка (Coregonus albula); 4 - сибирская ряпушка (C. sardinella); 5 - тугун (C. tugun); 6 - омуль (C. autumnalis); 7 - Байкальский омуль (C. a. migratorius).

Рис. 11. Лососевые: 1 - пелядь (Coregonus peled); 2 - чир (C. nasus); 3 - пыжьян (C. lavaretus pidschian); 4 - муксун (C. muksun).

Рис. 12. Лососевые: 1 - амурский сиг (C. ussuriensis); 2 - валёк (Prosopium cylindraceus); 3 - волховский сиг (Coregonus lavaretus baeri); 4 - лудога (C. l. ludoga).

Рис. 13. Лососевые: 1 - валаамка (Coregonus lavaretus widegreni); 2 - чудской сиг (C. l. maraenoides); 3 - енисейский речной сиг (C. l. pidschian n. fluviatilis).

Рис. 14. Хариусовые, корюшковые и салангасовые: 1 - хариус (Thymallus thymallus); 2 - байкальский хариус (T. arcticus baicalensis); 3 - сибирский хариус (T. arcticus); 4 - азиатская корюшка (Osmerus eperlanus dentex); 5 - малоротая корюшка (Hypomesus olidus); 6 - корюшка (Osmerus eperlanus); 7 - озерная корюшка, или снеток (O. e. eperlanus morpha spirinchus); 8 - лапша-рыба (Salangichthus microdon).

Рис. 15. Щуковые, чукучановые и карповые: 1 - щука (Esox lucius); 2 - щука с пятнистой окраской (E. lucius); 3 - чукучан (Catostomus catostomus rostratus); 4 - плотва (Rutilus rutilus).

Рис. 16. Карповые: 1 - тарань (Rutilus rutilus heckeli); 2 - астраханская вобла (R. r. caspicus); 3 - кутум (R. frisii kutum); 4 - китайская плотва, или черный амур (Mylopharyngodon piceus).

Рис. 17. Карповые: 1 - елец (Leuciscus leuciscus); 2 - сибирский елец (L. l. baicalensis); 3 - чебак иссыкульский (L. schmidti); 4 - голавль (L. cephalus).

Рис. 18. Карповые: 1- восточная "красноперка" (Leuciscus brandti); 2 - язь (L. idus); 3 - амурский язь (L. waleckii); 4 - красноперка (Scardinius erythophthalmus); 5 - озерный гольян (Phoxinus percnurus).


Рис. 20. Карповые: 1 - подуст (Chondrostoma nasus); 2 - пескарь (Gobio gobio); 3 - севанская храмуля (Varicorhinus capoeta sevangi); 4 - закаспийская храмуля (V. c. heratensis); 5 - аральский усач (Barbus brachicephalus).

Рис. 21. Карповые: 1 - днепровский усач, или марена (Barbus barbus borysthenicus); 2 - усач булат-маи, или усач-чанари (B. capito); 3 - конь-губарь (Hemibarbus labeo); 4 - балхашская маринка (Schizothorax argentatus); 5 - илийская маринка (Sch, pseudaksaiensis).

Рис. 22. Карповые: 1 - голый осман (Diptychus dybowskii); 2 - аральская шемая (Chalcalburnus chalcoides aralensis); 3 - черноморская шемая (Ch. ch. danubicus); 4 - густера (Blicca bjoerkna); 5 - уклея, или уклейка (Alburnus alburnus).


Рис. 24. Карповые: 1 - каспийский рыбец (Vimba vimba persa); 2 - черный амурский лещ (Megalobrama terminalis); 3 - белый амурский лещ (Parabramis pekinensis).

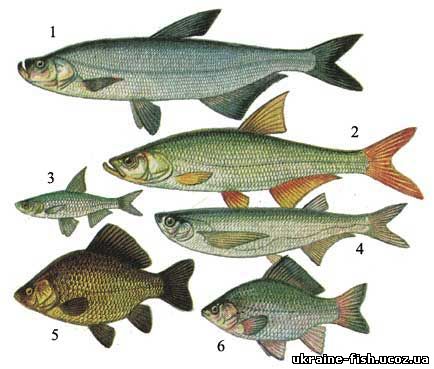

Рис. 27. Вьюновые, сомовые, косатки: 1 - пятнистый губач (Nemachilus strauchi); 2 - сом (Silurus glanis); 3 - амурский сом (Parasilurus asotus); 4 - косатка-скрипун (Pseudobagrus fulvidraco).

Рис. 28. Косатки, угревые, полурыловые, тресковые и колюшковые: 1 - косатка уссурийская (Liocassis ussuriensis); 2 - речной угорь (Anguilla anguilla); 3 - налим (Lota lota); 4 - японский полурыл (Hyporhamphus sajori); 5 - трехиглая колюшка (Gasterosteus aculeatus); 6 - девятииглая колюшка (Pungitius pungitius).

Рис. 29. Кефалевые, атериновые, змееголовые, серрановые: 1 - лобан (Mugil cephalus); 2 - пиленгас (M. so-iuy); 3 - каспийская атеринка (Atherina mochon pontica caspia); 4 - змееголов (Ophiocephalus argus warpachowskii); 5 - ауха, или китайский ерш (Siniperca chua-tsi).


Рис. 31. Окуневые, смаридовые, бельдюговые, подкаменщиковые, байкальские широколобки: 1 - балхашский окунь (Perca schrenki); 2 - азовская перкарина (Percarina demidoffi maeotica); 3 - ерш (Acerina cernua); 4 - смарида (Smaris smaris); 5 - бельдюга (Zoarces viviparus); 6 - ледовитоморская рогатка (Myoxocephalus quadricornis labradoricus); 7 - песчаная широколобка (Cottus kessleri).

Рис. 32. Байкальские широколобки, камбаловые: 1 - красная широколобка (Procottus jeitelesi); 2 - желтлкрылка, черногривка, или желтогривка (Cottocomephorus grewingki); 3 - речная камбала (Pleuronectes flesus); 4 - тихоокеанская речная, или звездчатая, камбала (P. stellatus)

Определитель составлен по материалам книги Е. А. Веселова "Определитель пресноводных рыб фауны СССР"; Москва, "Просвещение", 1977 г.
Понравилась статья? Подпишитесь на канал, чтобы быть в курсе самых интересных материалов
Каждый хороший рыболов обязательно следит за погодой перед выходом на рыбалку. Погодные условия очень сильно на нее влияют.

Рыба очень реагирует на изменение давления в атмосфере, при более высоком уходит на глубину, а при низком, наоборот, поднимается повыше. Направление и скорость ветра играют большую роль, также влияет и температура воды.
Хорошая погода для ловли рыбы:
- Спокойные дни без ветра.
- После зимы, когда наступает хорошая погода, рыба ненасытна и ловится на ура.
Нежелательно ловить при следующей погоде:
- При северных ветрах рыба не особо клюет.
- При резких изменениях уровня воды.
- При сильном дожде с ветром.
- При внезапных похолоданиях.
- Когда ветер начинает дуть вдоль течения.
- В очень жаркую погоду, особенно в середине лета.
В основном о погоде узнают в прогнозах на TV или из интернета, многие пользуются специально для этого барометром. Но до сих пор остались люди, которые верят некоторым признакам природы.
Признаки ухудшения погоды:
- Красивые облака движутся с запада.
- Долго находится роса на траве – к возможной грозе.
- Давление в атмосфере падает несколько дней подряд.
- Облака вечером увеличиваются.
- Ночью ветер усиливается.
- Вечером стало теплее, чем утром.
- Солнце село в тучу.
- Ярко-красная заря с утра.
- Ласточки летают очень низко.
Признаки улучшения погоды:
- В облаках появились просветы.
- В лесу стало теплее, чем в поле.
- Ночью стало прохладно, но тихо.
- Вечером появился туман.
- Понемногу давление повышается.
- Ласточки летают очень высоко.
- В темное время суток много росы на траве.
- Днём ветер сильный, а вечером стих.
- Дым от огня идет вертикально.
Народные приметы для рыбалки:
Есть множество примет, которые реально работают.
Если лягушки усиленно начали квакать и прыгают на берегах водоемов, то будет дождь. Еще, если громко квакать начали – стоит ждать хорошую погоду, а если замолчали – к холодам.
Если одуванчик сжал свой шар – возможен дождь, а ноготки венчики свои развернули ранним утром – ясная погода будет, а если опоздали – к грозе.
Когда пчелы не улетают в свои ульи при приближении дождливых туч, то дождя особо не будет. А если наоборот, утром спрятались в ульи и начали гудеть, то дождь будет обязательно.
Комары, когда роем летают – хорошая погода будет.
Раки вышли на берег – к дождю.
Понравилась статья? Подпишитесь на канал, чтобы быть в курсе самых интересных материалов
Читайте также: