Анатомия и физиология слухового таламуса (медиального коленчатого тела, МКТ)
Добавил пользователь Алексей Ф. Обновлено: 22.01.2026
, prosencephalon. Развивается из конечного отдела нервой трубки и находится кпереди от среднего мозга. Состоит из промежуточного и конечного мозга.
ПРОМЕЖУТОЧНЫЙ МОЗГ
Эпиталамус
, epithalamus. Состоит из поводков, треугольников и спайки поводков, а также шишковидного тела и эпиталамической спайки.
Поводок
Борозда поводка
, sulcus habenulae (habenularis). Проходит между треугольником поводка и подушкой (таламуса). Рис. А.
Треугольник поводка
, trigonum habenulae (habenularе). Треугольная пластинка между мозговой полоской таламуса и поводком. Под ней расположены ядра поводка. Рис. А.
Спайка поводков
, commissura habenularum (habenularis). Образована волокнами поводков, которые пересекают среднюю линию и pасположена сверху от шишковидного углубления. Рис. Б.
Эпиталамическая (задняя) спайка
, commissura epithalamica (posterior). Расположена между шишковидным углублением и входом в водопровод мозга. Содержит перекрещивающиеся волокна от ближайших структур мозга. Рис. Б.
Шишковидное тело (шишковидная железа)
, corpus pineale (glandula pinealis). Лежит над пластинкой четверохолмия. Удерживается поводками, с которыми не имеет функциональных связей. Рис. А, Рис. Б, Рис. В.
Разрезы эпиталамуса
Медиальное и латеральное ядра поводка
Поводково-межножковый путь
, tractus habenulointerpeduncularis. Связь между поводками и межножковым ядром (среднего мозга). Рис. Г
Предкрышечное поле
, area pretectalis. Лежит между верхним краем верхнего холмика и эпиталамической спайкой. Рис. А, Рис. В.
Предкрышечные ядра
, nuclei pretectales. Лежат дорсолатерально от эпиталамической спайки и доходят до верхних холмиков пластинки четверохолмия. К ним подходят волокна от затылочной, предзатылочной коры и от зрительного тракта. Отростки нейронов этих ядер направляются к добавочному ядру Ш нерва, из которого обеспечивается иннервация сфинктера зрачка.
Субфорникальный орган
, organum subfornicale. Находится у межжелудочкового отверстия между столбами свода. Влияет на кровяное давление и водный обмен. Рис. Б.
Субкомиссуральный орган
, organum subcommissurale. Группа специализированных эпендимных клеток под эпиталамической спайкой. Рис. Б.
Задний таламус
Межталамическое сращение
, adhesio interthalamica. Соединение между правым и левым таламусом. Встречается в 70 - 85% случаев. Рис. Б.
Передний бугорок таламуса
, tuberculum anterius thalamicum. Возвышение на переднем конце таламуса, которое соответствует передним ядрам таламуса. Рис. А.
Мозговые пластинки наружная/внутренняя
, laminae medullares interna/externa. Состоят из белого вещества. Неполностью разделяют между собой ядра таламуса.Рис. Д.
Мозговая полоска таламуса
, stria medullaris thalamiса. Расположена на медиальной поверхности таламуса под его лентой и дорсально продолжается в поводок. Содержит волокна от свода, пограничной полоски и предспаечной перегородки. Рис. А, Рис. Б.
Подушка
Метаталамус
Медиальное коленчатое тело
Латеральное коленчатое тело
, corpus geniculatum laterale. Связано с верхним холмиком и зрительной корой. В нем заканчивается большинство волокон зрительного тракта. Рис. А, Рис. В.
Передний таламус
Гипоталамус
Предзрительное поле
, area preoptica. Находится сзади от lam.terminalis и спереди от паравентрикулярного и супраоптического ядер. Предположительно, его ядра входят в состав пути, соединяющего обонятельный тракт и серый бугор. Рис. Б.
Зрительный перекрест
, chiasma opticum. Перекрест медиальных волокон зрительного нерва перед зрительным трактом. Рис. Б, Рис. В.
Зрительный тракт
, tractus opticus. Часть зрительного проводящего пути между зрительным перекрестом и латеральным коленчатым телом. Находится на поверхности основания мозга. Рис. В.
Латеральный корешок
, radix lateralis. Волокна зрительного тракта, оканчивающиеся в латеральном коленчатом теле или верхнем холмике. Рис. В
Медиальный корешок
Сосцевидное тело
, corpus mamillare. Парное округлое возвышение на нижней поверхности промежуточного мозга, связанное с таламусом и средним мозгом. Рис. Б.
Анатомия и физиология слухового таламуса (медиального коленчатого тела, МКТ)
Ознакомьтесь с Условиями пребывания на сайте Форнит Игнорирование означет безусловное согласие. СОГЛАСЕН
Таламус головного мозга
Таламус — парное образование, составляющее основную массу промежуточного мозга (имеет около 120 различных ядер), получающий импульсы всех видов чувствительности, кроме обонятельных, и передающий их в кору больших полушарий и другие образования центральной нервной системы .
Таламус расположен латерал ьнее III желудочка. Он занимает дорсальную часть промежуточного мозга и отделяется от нижележащего гипоталамуса бороздой. Два таламуса соединены по средней линии у 70% людей посредством межталамической промежуточной ткани серого вещества. От базальных ядер таламус отделяется внутренней капсулой, состоящей из нервных волокон, соединяющих кору со стволовыми структурами и спинным мозгом. Многие волокна внутренней капсулы продолжают ход в каудальном направлении в составе ножек мозга.
Ядра и функции таламуса
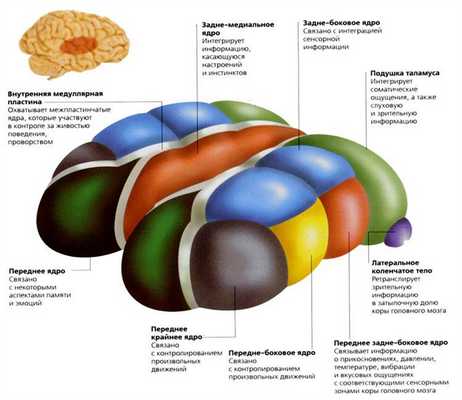
В таламусе выделяют до 120 ядер серого вещества. По месту их расположения ядра делят на передние, латерал ьные и медиальные группы. В задней части латерал ьной группы ядер таламуса выделяют подушку, медиальное и латерал ьное коленчатые тела.
Одной из важнейших функций таламуса является анализ, отбор и передача в кору головного мозга сенсорных сигналов, поступающих к нему из большинства сенсорных систем ЦНС. В этой связи таламус называют воротами, через которые в кору мозга поступают различные сигналы ЦНС. По выполняемым функциям ядра таламуса делятся на специфические, ассоциативные и неспецифические.
Специфические ядра характеризуются несколькими общими особенностями. Все они получают сигналы от вторых нейрон ов длинных восходящих афферентных путей, проводящих в кору мозга соматосенсорные, зрительные, слуховые сигналы. Эти ядра, иногда называемые сенсорными, передают обработанные сигналы в хорошо очерченные области коры — соматосенсорную, слуховую, зрительную сенсорные области, а также в премоторную и первичную моторные области коры. С нейрон ами этих областей коры специфические ядра таламуса имеют реципрокные связи. Нейрон ы ядер дегенерируют при разрушении (удалении) специфических областей коры, в которые они проецируются. При низкочастотной стимуляции специфических таламических ядер регистрируется усиление активности нейрон ов в тех областях коры, в которые нейрон ы ядер посылают сигналы.
К специфическим ядрам таламуса подходят волокна проводящих путей от коры, ретикулярной формации и ядер ствола мозга. По этим путям могут передаваться как возбуждающие, так и тормозные влияния на активность нейрон ов ядер. Благодаря таким связям кора мозга может регулировать потоки идущей к ней информации и отбирать наиболее значим ую в данный момент. При этом кора может блокировать передачу сигналов одной модальности и облегчать передачу другой.
Среди специфических ядер таламуса имеются также несенсорные ядра. Они обеспечивают обработку и переключение сигналов не от чувствительных восходящих путей, а от других областей мозга. К нейрон ам таких ядер поступают сигналы от красного ядра, базальных ганглиев, лимбической системы, зубчатого ядра мозжечка, которые после их обработки проводятся к нейрон ам моторной коры.
Ядра передней группы таламуса участвуют в передаче сигналов от мамиллярных тел к лимбической системе, обеспечивая круговую циркуляцию нервных импульсов по кольцу: лимбическая кора — гиппокамп — гипоталамус — миндалевидное тело — таламус — лимбическая кора. Нейрон ную сеть, сформированную этими структурами, называют кругом (кольцом) Пайпеца. Циркуляция сигналов по структурам этого круга связана с запоминанием новой информации и формированием эмоций — эмоциональное кольцо Пайпеца.
Ассоциативные ядра таламуса расположены преимущественно медиодорсально, латерал ьно и в ядре подушки. Они отличаются от специфических тем, что к их нейрон ам не поступают сигналы из чувствительных восходящих путей, а поступают сигналы уже обработанные в других нервных центрах и ядрах таламуса. Ассоциативность нейрон ов этих ядер выражается в том, что на один и тот же нейрон ядра приходят сигналы разных модальностей. Изменение активности нейрон ов ядер может быть связано (ассоциировано) с поступлением разнородных сигналов из разных источников (например, от центров, обеспечивающих зрительную, тактильную и болевую чувствительность).
Нейрон ы ассоциативных ядер являются полисенсорными и обеспечивают возможность осуществления интегративных процессов, в результате которых формируются обобщенные сигналы, передающиеся в ассоциативные области коры лобной, теменной и височной долей мозга. Потоки этих сигналов способствуют осуществлению корой таких психи ческих процессов, как узнавание предметов и явлений, согласование речевых, зрительных и двигательных функций, формирование представления о позе тела, трехмерности пространства и положении в нем тела человека.
Неспецифические ядра таламуса представлены преимущественно интраламинарными, центральными и ретикулярными группами ядер таламуса. Они состоят из мелких нейрон ов, к которым по многочисленным синаптическим связям поступают сигналы от нейрон ов других ядер таламуса, лимбической системы, базальных ядер, гипоталамуса, ствола мозга. По чувствительным восходящим путям к неспецифическим ядрам поступает сигнализация от болевых и температурных рецептор ов, а по сетям нейрон ов ретикулярной формации — сигнализация практически от всех других сенсорных систем ЦНС .
Эфферентные пути от неспецифических ядер идут ко всем зонам коры как непосредственно, так и через другие талами- ческие и ретикулярные ядра. От неспецифических ядер таламуса начинаются также нисходящие пути к стволу мозга. При повышении активности неспецифических ядер таламуса (например, при электрической стимуляции в эксперименте) регистрируется диффузное повышение нейрон ной активности практически во всех областях коры больших полушарий.
Принято считать, что неспецифические ядра таламуса благодаря своим многочисленным нейрон ным связям обеспечивают взаимодействие, координацию работы различных областей коры и других отделов головного мозга. Они оказывают модулирующее влияние на состояние активности нервных центров, создают условия для их оптимальной настройки на выполнение работы.
Нейрон ы различных ядер таламуса оказывают эффекты через высвобождение ГАМК из нервных окончаний, формирующих синапс ы на нейрон ах бледного шара, нейрон ах локальных цепей, нейрон ах ретикулярного ядра латерал ьного коленчатого тела; возбуждающие глутамат и аспартат в кортикоталамических, мозжечковых терминалях; таламокортикальных проекционных нейрон ах. Нейрон ами секретируются несколько нейропептидов преимущественно в окончаниях восходящих трактов (субстанция Р, сомагостатин, нейропептид Y, энкефалин, холецистокинин).
Метаталамус
Метаталамус включает два таламических ядра — медиальное коленчатое тело (MKT) и латерал ьное коленчатое тело (ЛКТ).
Ядро медиального коленчатого тела является одним из ядер слуховой системы. Его нейрон ы получают афферентные волокна из латерал ьного лемниска прямо или более часто, после их синаптического переключения на нейрон ах нижних холмиков. Эти слуховые волокна достигают MKT через соединительную ветвь нижних холмиков. MKT получает также волокна обратной связи из первичной слуховой коры височной области. Эфферентный выход ядра MKT формирует слуховую радиацию внутренней капсулы, волокна которой следуют к нейрон ам первичной слуховой коры (поля 41, 42).
Нейрон ы MKT вместе с нейрон ами нижних холмиков среднего мозга формируют нейрон ную сеть, выполняющую функцию первичного центра слуха. В нем осуществляется недифференцированное восприятие звуков, их первичный анализ и использование для формирования настораживания, повышения внимания и организации рефлекторного поворота глаз и головы в сторону неожиданного источника звука.
Ядро латерал ьного коленчатого тела является одним из ядер зрительной системы. Его нейрон ы получают афферентные волокна от ганглиозных клеток обоих сетчаток по зрительному тракту. Ядро ЛКТ представлено нейрон ами, расположенными в нескольких слоях (пластинках). Сигналы из сетчатки поступают в ЛКТ так, что ипси латерал ьная сетчатка проецируется к нейрон ам 2, 3 и 5-го слоев; контра латерал ьная — к нейрон ам 1,4 и 6-го слоев. К нейрон ам ЛКТ поступают также волокна обратной связи из первичной зрительной коры затылочной доли (поле 17). Нейрон ы ЛКТ, получив и обработав зрительные сигналы сетчатки, посылают сигналы по эфферентным волокнам, формирующим зрительную радиацию внутренней капсулы в первичную зрительную кору затылочной доли. Некоторые волокна проецируются в ядро подушки и вторичную зрительную кору (поля 18 и 19).
Латерал ьные коленчатые тела вместе с верхними холмиками относят к подкорковым зрительным центрам. В них осуществляется недифференцированное восприятие света, его первичный анализ и использование для формирования настораживания, повышения внимания и организации рефлекторного поворота глаз и головы в сторону неожиданного источника света.
Внутренняя капсула представляет собой широкий плотный пучок афферентных и эфферентных нервных волокон, соединяющих ствол и кору больших полушарий мозга. Волокна внутренней капсулы продолжаются рострально до радиации мозга и каудально до ножек мозга. Во внутренней капсуле проходят волокна таких важнейших нейрон ных нисходящих путей, как кортикоспинальный, кортикобульбарный, кортикорубральный, кортикоталамический, лобномостовой, кортикотекальный, кортиконигральный, кортикотегментальный и волокна восходящих таламокоркового, слухового и части зрительного путей.
Во внутренней капсуле тесно располагаются кортикоталамические и таламокортикальные волокна, поэтому при кровоизлияниях и заболеваниях этой области мозга возникают нарушения, характеризующиеся большим разнообразием, чем при повреждении какой-либо другой области ЦНС. Они могут проявиться развитием контралагеральной гемиплегии, потерей чувствительности на половине тела, потерей зрения на контра латерал ьной стороне (гемианопсия) и потерей слуха (гемигипоакузия).
Функции таламуса и последствия их нарушении
Таламус играет центральную роль в обработке сенсорной информации поступающей к коре больших полушарий мозга . Все сенсорные сигналы соматической и других видов чувствительности, за исключением обоняния, проходят к коре через таламус. Как уже упоминалось, сенсорная информация направляется таламусом в кору по трем каналам: в строго специфичные сенсорные области — от специфических ядер, MKT, ЛKT; в ассоциативные области коры — от ассоциативных ядер и ко всей коре — от неспецифических ядер таламуса.
Таламус участвует в частичном восстановлении таких сенсорных ощущений, как болевые, температурные и грубое осязание, которые исчезают после повреждения сенсорной коры. При этом восстановление ощущения боли, сигналы которого передаются волокнами С-типа, проявляется ноющей, жгучей, нс адресованной к какой-либо части тела болью. Предполагают, что центром таких болевых ощущений является таламус, в то время как ощущение острой, хорошо локализованной боли, передаваемой волокнами А-типа, является соматосенсорная кора. Это болевое ощущение исчезает после повреждения или удаления данной области коры.
У больных с острыми нарушениями кровообращения в области таламуса могут развиться признаки таламического синдрома. Одним из его проявлений является потеря всех видов чувствительности на контра латерал ьной половине тела по отношению к стороне поврежденного таламуса. Однако через некоторое время грубые ощущения боли, осязания и температуры восстанавливаются.
Одной из важнейших функций таламуса является интеграция сенсорной и моторной деятельности. Ее основой является поступление в таламус не только сенсорных, но и сигналов из моторных областей мозжечка, базальных ганглиев, коры. Предполагается, что в вентральном латерал ьном ядре таламуса локализован треморогенный центр.
Таламус, в котором находится часть нейрон ов ретикулярной формации ствола мозга, играет центральную роль в поддержании сознания и внимания. При этом его роль в осуществлении реакций активации и пробуждения реализуется при участии холинергических, серотонинергических, норадренергических и гнетаминергических нейромедиаторных систем, которые начинаются в стволе мозга (ядро шва, голубоватое пятно), основании переднего мозга или гипоталамусе.
Через связи медиального таламуса с прсфронтальной корой таламус участвует в формировании аффективного поведения. Удаление префронтальной коры или ее связей с дорзомедальным ядром таламуса вызывает изменения личности, характеризующиеся потерей инициативы, вялостью аффективной реакции, индифферентностью к боли.
Через связи передних таламических и других ядер таламуса с гипоталамусом и лимбическими структурами мозга обеспечивается их участие в механизмах памяти, контроля висцеральных функций, эмоционального поведения. При заболеваниях таламуса могут развиться различные типы нарушений памяти от мягкой забывчивости с рассеянностью до выраженной амнезии.
Таламус
Таламус(Рис. 7) (зрительный бугор) - это парное образование, расположенное по обеим сторонам 3-го желудочка. В переднем отделе таламус суживается и заканчивается передним бугорком. Задний конец утолщен и называется подушкой.
С нервными клетками таламуса вступают в контакт отростки нервных клеток вторых (кондукторных) нейронов всех проводящих путей (за исключением обонятельного, вкусового и слухового). В связи с этим таламус фактически является подкорковым чувствительным центром. Кроме того, именно таламус регулирует и координирует внешнее проявление эмоций человека (мимику, жесты, изменение пульса, дыхания, давления). Часть отростков нейронов таламуса направляется к ядрам полосатого тела конечного мозга (в связи с этим таламус рассматривается как чувствительный центр экстрапирамидной системы), а часть - таламокортикальные пучки, - к коре большого мозга. Под таламусом располагается так называемая субталамическая область - гипофиз. Сверху к нему прилегает эпифиз.
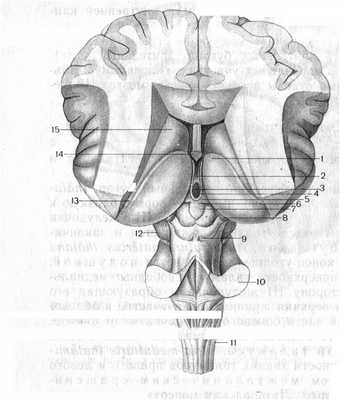
Рис. 7. Ствол головного мозга:
1 - таламус; 2 - межталамическое сращение; 3 - желудочек; 4 - мозговая полоска таламуса; 5 - треугольники поводка; 6 - поводок; 7 - шишковидное тело; 8 - медиальное коленчатое тело; 9 - пластинка крыши мозга; 10 - средняя мозжечковая ножка; 11 - продолговатый мозг; 12 - ножки мозга; 13 - терминальная пластинка; 14 - столб свода; 15 - хвостатое ядро
Таламус состоит из серого вещества, в котором различают отдельные скопления нервных клеток - ядра таламуса. Эти скопления разделены тонкими прослойками белого вещества. Современные ученые выделяют около 60 парных ядер, которые ученые подразделяют на три группы: релейные, ассоциативные и неспецифические. Все ядра таламуса, правда в разной степени, но обладают тремя общими функциями: переключающей, интегративной и модулирующей.
Релейные ядра таламуса (синоним - переключательные, специфические). Делятся на сенсорные и несенсорные.
Сенсорные релейные ядра переключают потоки афферентной (чувствительной) импульсации в сенсорные зоны коры. В них также происходит перекодирование и обработка информации.
Главным «реле» для переключения сенсомоторной афферентной системы являются и вентральные задние ядра (их еще называют вентробазальным комплексом). Импульсация из вентральных задних ядер проецируется в соматосенсорную кору постцентральной извилины, в которой и формируются соответствующие ощущения. Так, формирование зрительных ощущений способствует латеральное коленчатое тело, которое переключает зрительную импульсацию в затылочную кору. Кроме корковой проекции, часть зрительной импульсации направляется в верхние бугры четверохолмия. Данная информавция используется для регуляции движения глаз, а также в зрительном ориентировочном рефлексе. Реле для переключения слуховой импульсации в височную кору задней части сильвиевой борозды (извилины Гешля) является медиальное коленчатое тело.
В результате проведенных исследований в релейных ядрах учеными обнаружено несколько типов нейронов. Таламокортикальные (релейные) нейроны, имеющие длинный аксон, непосредственно обеспечивают переключение поступающей афферентной импульсации на нейроны сенсорной коры. Особенностью этой передачи является ее высокая точность и искажение исходного сигнала бывает минимальным. Регуляция передачи возбуждения через релейные ядра осуществляется с помощью тормозных и возбуждающих вставочных нейронов. Тормозные интеронейроны возбуждаются как от таламокортикальных нейронов ядра, осуществляя при этом возвратное торможение, так и от кортиковых нейронов, проецирующихся в данное ядро и осуществляющих корковый контроль проведения возбуждения.
Несенсорные релейные ядра таламуса (передние и вентральные) переключают в кору несенсорную импульсацию, поступающую в таламус из разных отделов головного мозга.
В передние ядра таламуса импульсация в основном поступает из гипоталамуса. Нейроны передних ядер проецируются в лимбическую кору, откуда аксонные связи идут к гиппокампу и опять возвращаются к гипоталамусу. В результате образуется нейронный круг, движение возбуждения по которому обеспечивает формирование эмоций («эмоциональное кольцо Пейпеца»). Не случайно передние ядра таламуса нередко рассматривают как часть лимбической системы и обозначают «лимбическими ядрами таламуса».
Вентральные ядра участвуют в регуляции движений, выполняя моторную функцию. В этих ядрах переключается импульсация от базальных ганглиев, зубчатого ядра мозжечка, красного ядра среднего мозга, которая проецируется в моторную и премоторную кору.
Наряду с корковыми проекциями релейных ядер каждое из них получает нисходящие корковые волокна из той же проекционной зоны, что создает структурную основу для взаиморегулирующих отношений между корой и таламусом.
Ассоциативные ядра таламуса принимают импульсацию не от проводниковых путей анализаторов, а от целого ряда других ядер таламуса. Эфферентные выходы от этих ядер направляются, главным образом, в ассоциативные поля коры. В свою очередь, кора мозга посылает волокна к ассоциативным ядрам, регулируя их функцию. При этом главной функцией этих ядер является интегративная функция, которая выражается в объединении деятельности как таламических ядер, так и различных зон ассоциативной коры полушарий мозга. Так, подушка получает главную импульсацию от коленчатых тел и неспецифических ядер таламуса. Эфферентные пути от нее идут в височно-теменно-затылочные зоны коры, участвующие в гностических (узнавание предметов, явлений), речевых и зрительных функциях (интеграция слова со зрительным образом). Медиодорсальное ядро получает импульсацию от гипоталамуса, миндалины, гиппокампа, таламических ядер, центрального серого вещества ствола. Проекция этого ядра распространяется на ассоциативную лобную и лимбическую кору и участвует в формировании эмоциональной и поведенческой двигательной активности, а также (по предположению ученых) в образовании памяти. Разрушение этих ядер устраняет у больных страх, тревогу, напряженность, страдание от боли и приводит к снижению инициативы, безразличию и т.д.
Неспецифические ядра составляют эволюционно более древние структуры таламуса, включающую парные ретикулярные ядра и интраламинарную ядерную группу. Ретикулярные ядра преимущественно содержат мелкие, многоотростчатые нейроны и функционально рассматриваются как производные ретикулярной формации ствола мозга.
Неспецифические ядра имеют многочисленные входы от других ядер таламуса, а также внеталамические входы, например, по латеральному спиноталамическому, спиноретикулоталамическому путям, проводящим преимущественно болевую и температурную чувствительность. Кроме того, в неспецифические ядра таламуса поступает информация от всех специфических сенсорных систем, моторных центров ствола, ядер мозжечка, базальных ганглиев и гиппокампа, а также от коры мозга (особенно ее лобных долей). Именно благодаря этим многочисленным связям ядра таламуса выступают в роли интегрирующего посредника между стволом мозга и мозжечком, с одной стороны, и новой корой, лимбической системой и базальными ганглиями, с другой стороны, объединяя их в единый функциональный комплекс. На кору мозга неспецифический таламус оказывает, в основном, модулирующее (изменяющее состояние) влияние. Разрушение неспецифических ядер не вызывает грубых расстройств эмоций, восприятия, сна и бодрствования и пр., а нарушает лишь тонкое регулирование поведения. В связи с этим модулирующее влияние неспецифических ядер таламуса, обеспечивающих «плавную настройку» высшей нервной деятельности, считается главной их функцией.
Промежуточный мозг Diencephalon
Промежуточный мозг (diencephalon) интегрирует сенсорные, двигательные и вегетативные реакции, необходимые для целостной деятельности организма. Основными образованиями промежуточного мозга являются: таламус, гипоталамус, эпиталамус и метаталамус.
Таламус (thalamus, зрительный бугор) — структура, в которой происходит обработка и интеграция практически всех сигналов, идущих в кору большого мозга от спинного, среднего мозга, мозжечка, базальных ганглиев головного мозга.
Морфофункциональная организация. В ядрах таламуса происходит переключение информации, поступающей от экстеро-, проприорецепторов и интероцепторов и начинаются таламокортикальные пути.
Зрительный бугор является центром организации и реализации инстинктов, влечений, эмоций. Возможность получать информацию о состоянии множества систем организма позволяет таламусу участвовать в регуляции и определении функционального состояния организма в целом (подтверждением тому служит наличие в таламусе около 120 разнофункциональных ядер).
Ядра таламуса образуют своеобразные комплексы, которые можно разделить по признаку проекции в кору на 3 группы: передняя, нейроны которой проецирует аксоны своих нейронов в поясную извилину коры большого мозга; медиальная — в лобную долю коры; латеральная — в теменную, височную, затылочную доли коры.
Функционально ядра таламуса по характеру входящих и выходящих из них путей делятся на специфические, неспецифические и ассоциативные.
К специфическим ядрам относятся переднее вентральное, медиальное, вентролатеральиое, постлатеральное, постмедиальное, латеральное и медиальное коленчатые тела. Последние относятся к району метаталамуса и являются подкорковыми центрами зрения и слуха соответственно.
Функция нейронов специфических ядер таламуса заключается в переключении информации, идущей в кору большого мозга от кожных, мышечных и других рецепторов. Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности.
Латеральное коленчатое тело имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
Медиальное коленчатое тело (МТК) получает слуховые импульсы из первичных слуховых центров ствола мозга и от нижних бугров четверохолмий. Эфферетные пути от медиальных коленчатых тел идут в височную зону коры большого мозга, достигая там первичной слуховой области коры.
Ассоциативные ядра таламуса представлены передним медиодорсальным, латеральным дорсальным ядрами и подушкой. Переднее ядро связано с лимбической корой (поясной извилиной), медиодорсальное — с лобной долей коры, латеральное дорсальное — с теменной, подушка — с ассоциативными зонами теменной и височной долями коры большого мозга.
Неспецифические ядра таламуса представлены срединным центром, парацентральным ядром, центральным медиальным и латеральным, субмедиальным, вентральным передним, парафасцикулярным комплексами, ретикулярным ядром, перивентрикулярной и центральной серой массой. Нейроны этих ядер образуют свои связи с корой большого мозга, где контактируют со всеми ее слоями. К неспецифическим ядрам поступают связи из РФ ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса.
Гипоталамус (hypothalamus) — небольшое образование передне-нижней части промежуточного мозга, весом около 4 г . Он образует дно III желудочка. На основании мозга между его ножками и сзади от перекреста зрительных нервов (хиазмы) видны основные структуры гипоталамуса. Серый бугор (tuber cineréum) расположен между сосцевидными телами и перекрестом зрительных нервов, с боков ограничен зрительными трактами. Он соединен с воронкой (infundibulum) которая переходит вентральнее в ножку гипофиза и далее в гипофиз (hypophysis). За серым бугром следуют белого цвета парные круглые образования — сосцевидные тела (corpora mamillaria). Сосцевидные тела граничат с задним продырявленным пространством (substantia perforata posterior). Эта часть поверхности мозга называется продырявленным пространством, потому что пронизана многочисленными сосудами. Серое вещество гипоталамуса образует ядра, которые подразделяются на пять групп: преоптическую, переднюю, среднюю, наружную и заднюю группы
Гипоталамус является высшим подкорковым центром интеграции нервных и эндокринных влияний, вегетативных и эмоциональных компонентов поведенческих реакций и тем самым обеспечивает регуляцию постоянства внутренней среды. Ядра гипоталамуса тесно связаны с другими мозговыми структурами. Эфферентные волокна сосцевидных тел образуют их главный пучок (tractus principalis), или мамиллоталамический (Вик д'Азира), который направляется к переднему ядру таламуса, а мамиллосегментальный — к покрышке среднего мозга (в пучке Вик д'Азира проходит 0,7 млн волокон). Ядра боковых и средних групп гипоталамуса имеют двусторонние связи с базальными ядрами больших полушарий головного мозга. Ядра серого бугра получают афференты от обонятельного мозга, а передняя группа ядер — от свода. Эфференты ядер гипоталамуса связывают их с таламусом, субталамусом и другими подкорковыми структурами. Особо следует выделить связи передней группы ядер с задней долей гипофиза (нейрогипофизом).
Гипофиз (hypophysis), или нижний мозговой придаток, располагается на вентральной поверхности мозга в основании черепа в ямке турецкого седла клиновидной кости. После рождения, по мере роста и развития ребенка вес гипофиза увеличивается и к 14 годам достигает массы гипофиза взрослого человека: 0,7 г у девушек и 0,6 г у юношей. В период беременности масса гипофиза у женщин увеличивается до 1 г. Гипоталамус и гипофиз составляют единую гипоталамо-гипофизарную функциональную систему, обеспечивающую совместную регуляцию функций нервным и гуморальным путем.
По своему строению и эмбриогенезу гипофиз не однороден. В гипофизе различают две главные части: нейрогипофиз и аденогипофиз, имеющие различное эмбриональное происхождение и строение.
НейрогипофизНейрогипофиз представляет собой производное дна воронки промежуточного мозга. Он находится в тесной морфологической и функциональной связи с гипоталамусом, в нем заканчиваются волокна гипоталамо-гипофизарного тракта, идущего от супраоптического и паравентрикулярного ядер гипоталамуса. В нейрогипофиза из аксонов нейросекреторных клеток в кровь секретируются нейрогормоны: вазопрессин и окситоцин
Аденогипофиз (передняя доля) Передняя доля гипофиза имеет тесную сосудистую связь с гипоталамусом. Здесь артерии ветвятся на капилляры, образуя плотное сплетение в форме мантии на поверхности срединного возвышения. Капиллярные ветви этого сплетения образуют вены, достигающие передней доли гипофиза, здесь вены вновь распадаются на капилляры, пронизывающие всю долю. Вся эта сложная система кровеносных сосудов носит название портальной. По ней в аденогипофиз из гипоталамуса поступают пептидные гормоны (либерины и статины), регулирующие синтез и секрецию гормонов аденогипофиза. В аденогипофизе секретируется два типа гормонов — эффекторные, т.е. реализующие свои свойства непосредственно в организме, и тропные — оказывающие регулирующее влияние на периферические железы внутренней секреции. Всего в аденогипофизе синтезируется шесть гормонов — гормон роста, пролактин, тиреотропин, адренокортикотропный гормон (АКТГ), фолликулостимулирующий гормон, лютенизирующий гормон. Фолликулостимулирующий и лютенизирующий гормоны объединяются в группу гонадотропных гормонов.
Эпиталамус (epythalamus) Мозговые полоски,поводки и эпифиз образуют эпиталамус. Эпифиз (corpus pineale, epyphisis) является железой внутренней секреции, контролирующей течение циркадианных ритмов и тормозящей активность половых желез до наступления периода полового созревания.
Головной мозг
Вентролатеральные ядра [таламуса] - Зрительная [[Грасиоле]] лучистость
Вентролатеральные ядра [таламуса]
Заднее латеральное ядро
, nucleus lateralis posterior. Расположено между подушкой (таламуса) и дорсальным латеральным ядром. Связано с корой теменной доли. Рис. А.
Дорсальное латеральное ядро
, nucleus lateralis dorsalis. Расположено спереди и сверху относительно остальных вентролатеральных ядер таламуса. Связано с корой нижней теменной дольки и задней части поясной извилины. Рис. А.
Переднее вентральное ядро
, nucleus ventrаlis anterior. Самое переднее из вентролатеральных ядер (таламуса). Соединяется с внутрипластинчатыми ядрами, бледным шаром и зубчатым ядром мозжечка, а также имеет двухсторонние связи с корой предцентральной извилины. Поражается при болезни Паркинсона. Рис. А.
Вентральное промежуточное ядро
Медиальное вентральное ядро
, nucleus ventralis medialis. Нечетко очерченная группа нейронов с неясной функцией спереди от задних вентральных ядер. Рис. А.
Задние вентральные ядра
Заднелатеральное вентральное ядро
, nucleus ventralis posterolateralis. Латеральная часть заднего вентрального ядерного комплекса. На нейронах этого ядра заканчиваются волокна медиальной петли и спинно-таламического пути. Отсюда импульсы по таламо-корковым волокнам передаются к постцентральной извилине. Рис. А.
Заднемедиальное вентральное ядро
, nucleus ventralis posteromedialis. Находится между центральным срединным и заднелатеральным вентральным ядрами. К нему подходят волокна тройничной петли. Рис. А.
Задние ядра [таламуса]
Ядра подушки
Латеральное ядро [коленчатого тела] [дорсальная часть]
, nucleus [corporis geniculati] lateralis [pars dorsalis]. Нейроны этого ядра входят в состав зрительного пути. Рис. А.
Медиальное ядро [коленчатого тела] [дорсальная часть]
Разрезы переднего таламуса
Ядро латерального коленчатого тела [вентральная часть]
Ядро медиального коленчатого тела [вентральная часть]
, nucleus corporis geniculati medialis [pars ventralis]. Предположительно, основная часть соответствующего коленчатого тела, которая имеет отношение к слуховому пути. Рис. В.
Субталамическое ядро [люисово тело]
, nucleus subthalamicus [corpus Luysii]. Находится между внутренней капсулой и неопределенной зоной. Клиническое значение имеют его связи с бледным шаром. Рис. Б.
Ретикулярные ядра [таламуса]
, nucleireticulares [thalami]. Расположены в латеральной части таламуса между наружной мозговой пластинкой и внутренней капсулой. Рис. Б.
Неопредленная зона
Ядра полей Н, Н1 и Н2
, nuclei regionum H, H1, H2. Группы диффузно расположенных нейронов в составе полей Фореля. Поле Н лежит медиально от zona incerta и спереди от красного ядра. Поле Н1 расположено между таламусом и неопределенной зоной. Поле Н2 находится между zona incerta и субталамическим ядром. Рис. Б.
Пути и пучки таламуса
Латеральная петля
, lemniscus lateralis. Образована волокнами слухового пути, которые вступают в медиальное коленчатое тело. Рис. А.
Медиальная петля
, lemniscus mediаlis. Ее волокна заканчиваются на нейронах заднелатерального вентрального ядра. Рис. А.
Спинномозговая петля
, lemniscus spinalis. Путь, проводящий болевую чувствительность. Заканчивается в заднелатеральном вентральном ядре. Рис. А.
Тройничная петля
, lemniscus trigeminalis. Образована отростками нейронов чувствительных ядер тройничного нерва, которые заканчиваются в заднемедиальном вентральном ядре. Рис. А.
Ручка нижнего холмика
Слуховая лучистость
, radiatio acustica. Часть слухового пути. Начинается от медиального коленчатого тела, проходит через подчечевицеобразную часть задней ножки внутренней капсулы и заканчивается в поперечных височных извилинах. Рис. А.
Ручка верхнего холмика,
, brachium colliculi superioris. Соединяет верхний холмик с латеральным коленчатым телом. Связывает зрительный путь с экстрапирамидной системой. Рис. В.
Зрительная [[Грасиоле]] лучистость
, radiatio optica [[Gratiolet]]. Часть зрительного пути. Начинается от латерального коленчатого тела, проходит через подчечевицеобразную часть задней ножки внутренней капсулы, огибает задний рог бокового желудочка и заканчивается в зрительной коре (область шпорной борозды). Рис. А.
Читайте также: