Электрофизиология сердца: мембранный (монофазный) потенциал действия
Добавил пользователь Евгений Кузнецов Обновлено: 22.01.2026
Кардиология:
Мембранный потенциал покоя кардиомиоцитов (КМЦ) сердца (фаза 4 потенциала действия)
Внутриклеточный потенциал во время электрического покоя в течение диастолы колеблется от -50 до -95 мВ в зависимости от типа клетки, поэтому заряд внутри клетки на 50-95 мВ более отрицателен, чем снаружи, что обусловлено распределением таких ионов, как К + , Na + и Cl - .
Поскольку в КМЦ в состоянии покоя имеется избыток открытых K + -каналов, трансмембранный потенциал (в фазе 4) близок к Еk. Выходящий ток калия через открытые К + -каналы, выпрямляющие входящий ток IKI, в нормальных условиях в значительной мере зависит от мембранного потенциала покоя предсердных и желудочковых КМЦ, а также клеток Пуркинье.
Отличие мембранного потенциала покоя от Еk обусловлено движением одновалентных ионов с равновесным потенциалом большим, чем Еk, например током Cl - через активированные хлоридные каналы, такие как ICi.camp, ICl.Ca и IЕCl.swellКальций напрямую не влияет на мембранный потенциал покоя, но изменения концентрации свободного внутриклеточного кальция могут отразиться на других показателях мембранной проводимости.
Например, повышение нагрузки Са 2+ в СР может обусловить появление спонтанных кальциевых волн, которые, в свою очередь, активируют Са 2+ -зависимую хлоридную проводимость ICl.Ca и, следовательно, приводят к возникновению спонтанных транзиторных входящих токов и сопутствующей мембранной деполяризации. Повышение [Са 2+ ], также может стимулировать Na + /Ca 2+ -обмен (INa/Ca). Этот белок меняет 3 иона Na+ на 1 ион Са 2+ ; направление зависит от концентрации натрия и кальция по обе стороны мембраны и разницы трансмембранного потенциала.
Во время мембранного потенциала покоя и во время спонтанного высвобождения Са 2+ в СР этот обменник будет генерировать итоговый ток Na + , возможно вызывающий временную мембранную деполяризацию. Также было показано, что [Са 2+ ], активирует IKI в КМЦ и таким образом косвенно влияет на кардиальный мембранный потенциал покоя. Поскольку Na + /K + -нacoc перекачивает Na + из клетки против его электрохимического градиента и одновременно отправляет К + в клетку также против его электрохимического градиента, концентрация внутриклеточного К + остается высокой, а концентрация внутриклеточного Na + — низкой.
Этот насос, энергетический ресурс которого обеспечивается ферментом Na + /К + -АТФазой, гидролизирующей АТФ для высвобождения энергии, связан с мембраной. Для его работы необходимы и Na + , и К + ; он может перемещать 3 иона Na + из клетки и 2 иона К + — внутрь клетки. Следовательно, насос может быть электрогенным, а итог его работы — направленный наружу поток положительных зарядов.
Скорость работы Na + /K + -насоса для увеличения тех же ионных градиентов может повышаться подобно ЧСС, поскольку клетка при каждой деполяризации накапливает небольшое количество ионов Na + и теряет небольшое количество ионов К + .
Гликозид-индуцированное ингибирование Na + /К + -АТФазы повышает сократимость КМЦ за счет увеличения концентрации внутриклеточного Na + , который, в свою очередь, уменьшает вытеснение Са 2+ путем Na + /Са 2+ -обмена, что в итоге и повышает сократимость клеток.
ЭЛЕКТРОФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ЭЛЕКТРОКАРДИОГРАФИИ
Функция сердца как органа, ответственного за перемещение крови в организме человека, включает в себя:
1) автоматизм - способность сердца к спонтанной диастолической деполяризации, т.е. способность вырабатывать импульсы, вызывающие возбуждение миокарда;
2) проводимость - проведение импульса к рабочему миокарду;
3) возбудимость - способность к возбуждению под влиянием импульсов;
4) рефрактерность - невозможность возбуждения при определенных обстоятельствах;
5) сократимость - способность сокращаться под воздействием импульсов, что обеспечивает продвижение крови в организме человека.
Электрокардиограмма позволяет оценить все указанные функции сердца, за исключением сократимости миокарда.
Потенциал действия сердечной клетки
В покое все клеточные мембраны поляризованы таким образом, что поверхность одиночных мышечных волокон и миокарда в целом заряжена положительно. Это обусловлено избирательной проницаемостью клеточной мембраны и постоянным действием ионных насосов. В состоянии покоя концентрация ионов калия (К + ) внутри клетки в 30 раз выше их внеклеточной концентрации, внеклеточные концентрации ионов натрия (Na + ) в 20 раз, анионов хлора (Cl - ) в 13 раз, а кальция (Ca 2+ ) в 25 раз выше их концентрации внутри клетки. Эти различия в концентрации ионов и их зарядов определяют поляризацию мембраны кардиомиоцита, формируя трансмембранный потенциал покоя(ТМПП) который составляет около -85mV.
Деполяризация , или активация клетки под влиянием электрического импульса, приводит к изменению заряда мембран: внешняя сторона возбужденного участка (клетки, волокна, всего миокарда) приобретает отрицательный заряд. Это объясняется тем, что при возбуждении изменяется проницаемость мембран по отношению к ионам разных типов, что влечет за собой перемещение ионов через ионные каналы мембраны, изменение их концентрации и, соответственно, трансмембранного потенциала (трансмембранный потенциал действия,
ТМПД). Появление и стремительное распространение ТМПД создает разность потенциалов между активированным и неактивированным миокардом и формирует электродвижущую силу (ЭДС). По завершении деполяризации разность потенциалов между соседними участками миокарда исчезает, так как вся поверхность миокарда становится электроположительной.
Сущность реполяризации заключается в активном перемещении ионов с помощью ионных насосов, восстановлении исходного ионного статуса и потенциала покоя, обеспечивая готовность к очередному возбуждению, т.е. в восстановлении положительного заряда внешней стороны клеточных мембран. Постепенное замещение им отрицательного заряда вновь создает ЭДС - на этот раз ток реполяризации.
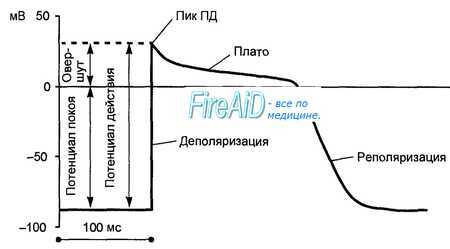
Рисунок. Фазы потенциала действия одиночного кардиомиоцита фаза 0 - начальная фаза деполяризации; фаза 1 - фаза начальной быстрой реполяризации (небольшое
начальное снижение ТМПД от +20 mV до 0 или чуть ниже);
фаза 2 - фаза плато (величина ТМПД поддерживается на одном уровне);
фаза 3 - фаза конечной быстрой реполяризации (восстанавление исходной поляризации клеточной мембраны);
фаза 4 - фаза диастолы (величина ТМПД сократительной клетки сохраняется примерно на уровне -85 mV, происходит восстановление исходной концентрации К + , Na + , Ca 2 + и Cl - , благодаря работе «Na + -K + -
В начале ТМПД (фазы 0, 1, 2) клетки полностью не возбудимы (абсолютный рефракторный период). Во время быстрой конечной реполяризации (фаза 3) возбудимость частично восстанавливается (относительный рефракторный период). Во время диастолы (фаза 4 ТМПД) рефрактерность отсутствует и кардиомиоцит полностью готов к новому возбуждению.
Система автоматизма сердца
Свойством генерировать самостоятельно электрический импульс возбуждения наделены специализированные, так называемые пейсмекерные клетки (ПК) проводящей системы сердца. Способность к автоматизму, принципиально отличает их от сократительных клеток миокарда. Так в норме сократительные кардиомиоциты сохраняют в диастолу постоянное значение ТМПП, равное -85 mV, а ПК характеризуются медленным спонтанным уменьшением мембранного потенциала, носящим название медленной спонтанной диастолической деполяризации , обусловленным медленным самопроизвольным поступлением внутрь клетки Na + . Как только ТМПП ПК достигнет порогового уровня (-60mV), ионные токи лавинообразно возрастают, что приводит к быстрой деполяризации мембраны, возбуждению клетки и влечѐт за собой возбуждение окружающих клеток сократительного миокарда, которые активизируются только под влиянием импульсов, исходящих из ПК.
Наивысший автоматизм с максимальной частотой генерации импульсов присущ синоатриальному (СА) узлу, который подавляет все другие источники импульсов с меньшей частотой и в норме выступает в качестве основного водителя ритма или центра автоматизма I порядка. Нижерасположенные ПК - в предсердиях, атриовентрикулярном (АВ) соединении и желудочках, являются центрами автоматизма II и III порядка и в обычных условиях работают в режиме пассивных проводников возбуждения.
Порядок возбуждения миокарда
Возникая в СА-узле, импульс возбуждения активирует сначала правое, затем левое предсердия и, после задержки в АВ-соединении (физиологическая задержка проведения в АВ-узле способствует тому, что желудочки начинают возбуждаться только после окончания полноценного сокращения предсердий) по системе Гиса передаѐтся на миокард желудочков. В желудочках возбуждение начинается с межжелудочковой перегородки (возбуждение охватывает перегородку слева направо). Далее импульс распространяется на стенки желудочков начиная с верхушки
сердца. Последними возбуждаются базальные отделы правого и левого желудочков.
Рисунок. Проводящая система сердца и последовательность возбуждения отделов сердца (А) в сопоставлении с ЭКГ (Б)
В миокарде ход возбуждения направлен с внутренней субэндокардиальной области, где ветвятся волокна Пуркинье, к эпикарду. Таким образом, возбуждение стенок желудочков происходит в направлении изнутри кнаружи.
Векторная теория
Де - и реполяризация клеточной мембраны могут рассматриваться в качестве диполя, который подразумевает сосуществование и перемещение двух равных по величине, но разных по знаку зарядов, находящихся на бесконечно малом расстоянии друг от друга. В процессе возбуждения миокарда в сердце начинает функционировать огромное количество микродиполей одиночных мышечных волокон - элементарных источников электродвижущей силы (ЭДС). Суммируясь, они образуют более крупные макродиполи отдельных участков миокарда, отделов сердца и в конечном итоге образуют единый сердечный диполь и ЭДС всего сердца, которые распространяется на поверхность человеческого тела и служит основой для регистрации ЭКГ. При этом следует учитывать, что ЭДС - векторная величина, которая характеризуется не только количественным значением потенциала, но и его направлением -
Основы электрофизиологии сердца
Сердечная мышца является неоднородной с точки зрения структуры и функции. Основную ее массу составляют кардиомиоциты,обеспечивающие реализацию механической функции сердцаи способные воспринимать импульсы возбуждения,которые образуются в клетках специализированной проводящей системысердца, обеспечивающей реализацию биоэлектрической активности сердечной мышцы.
В состав проводящей системы сердца входят клетки, способные вырабатыватьимпульсы возбуждения (пейсмекеры или Р-клетки), проводящие миоциты,обеспечивающие проведениеимпульса возбуждения, и клетки (волокна) Пуркинье,терминальные клетки проводящей системы сердца, обеспечивающие непосредственную передачуимпульса возбуждения на кардиомиоциты,в которых, благодаря системе электромеханического сопряжения, энергия электрического импульса преобразуется в энергию механического сокращения.
Клетки-пейсмекерыобразуют два скопления,одно из которых(синусовыйузел) располагается в правом предсердии в устье полых вен, второе (атриовентрикулярное (АВ) соединение)- в нижней части межпредсердной перегородки.
Проводящие миоциты предсердий,непосредственно соприкасаясь с синусовым узлом и АВ-соединением, образуют проводящую систему предсердийв виде межузловых и межпредсердных трактов,объединяющих два узла (синусовый и атриовентрикулярный) и оба предсердия.
Проводящая система желудочков (система Гиса-Пуркинье), состоит из проводящих миоцитов, которые, непосредственно примыкая к АВ-соединению, образуют ствол, ножки(правая для правого желудочка, левая - для левого) и ветви пучка Гиса,заканчивающиесяволокнами (клетками) Пуркинье.
Автоматам -способность сердечной мышцы самостоятельно вырабатывать импульсы возбуждения.
В нормеимпульсы возбуждения вырабатываются в синусовом узле,который является основным водителем ритмадля сердечной мышцы. При слабости или отказесинусового узла функцию водителя ритма берет на себя АВ-соединение,при слабости или отказе АВ-соединения функция водителя ритма передается проводящим миоцитам системы Гиса-Пуркинье.
Синусовый узел является облигатным(обязательным) водителем ритма, АВ-соединение и проводящие миоциты желудочков - факультативными(необязательными), составляя вспомогательную (резервную) систему,клеточные элементам которой в экстремальной. ситуации берут на себя роль водителя ритма.
Уровень автоматизма, то есть способность вырабатывать импульсы возбуждения, у здорового человека максимален у синусового узла, поэтому он называется центром автоматизма первого порядка. Несколько ниже - у АВ-соединения(центр автоматизма второго порядка), еще ниже у проводящих миоцитов желудочков (центры автоматизма третьего и четвертого порядка), при этом в желудочках уровень автоматизма снижается по направлению к дистальным отделам системы Гиса-Пуркинье.
Уровень автоматизма определяется частотой,с которой водитель ритма способен вырабатывать импульсы возбуждения. Синусовый узел может вырабатывать импульсы возбуждения с частотой от 200 (и выше) до 60 (и ниже) в минуту (в зависимости от ситуации -при физической нагрузкечастота синусового ритмаувеличивается, в спокойном состоянии - уменьшается, во время ночного сна снижается до 60 и ниже). В состоянии покоя частота синусового ритма у здорового человека составляет примерно 60-85 в минуту.
Способность синусового узла изменять уровень автоматизма в широких пределах позволяет человеку легко адаптироваться к меняющимся условиям окружающей среды, выполнять тяжелую физическую работу.
Уровень автоматизма синусового узла регулируетсясимпатической и парасимпатической нервной системой (катехоламины повышают, ацетилхолин понижает частоту образованияимпульса возбуждения в клетках пейсмекерах синусового узла). На частоту образования импульса возбуждения в синусовом узле влияет также температура тела и окружающей среды. При повышении температуры тела автоматизм и, следовательно, частота выработки импульсов возбуждения в синусовом узле увеличивается
АВ-соединениевырабатывает импульсы возбуждения с частотой 70-60 в минуту, причем частота узлового ритма (ритма из АВ-соединения) существенно не меняется в зависимости от уровня физической активности человека. Активность АВ-соединения также находится под вегетативным контролем, однако влияние симпатической и парасимпатической нервной системы здесь менее выражено, чем на уровне синусового узла, что объясняет его определенную физиологическую ригидность
Проводящие миоциты системы Гиса-Пуркиньевырабатывают импульсы возбуждения с частотой 50-25 в минуту и ниже (в зависимости от локализации идиовентрикулярного,то есть желудочкового, водителя ритма).
Импульсы возбуждения образуются в клетках-пейсмекерах в процессе медленной диастолической деполяризацииблагодаря перемещению ионов калия, натрия и кальция через полупроницаемую клеточную мембрану по медленным ионным каналам в двух направлениях (из межклеточного пространства в клетку и наоборот).
Перемещение ионов через клеточную мембрану в фазу медленной диастолической деполяризации приводит к постепенному увеличению внутриклеточного заряда Р-клетки, который становиться выше, чем заряд ее клеточной мембраны, после чего импульс возбуждения "выплескивается" из Р-клетки на проводящие миоциты предсердий.
Проводимость.Проведение импульса возбуждения по проводящим миоцитам осуществляется по тому же механизму, который обеспечивает распространение волны возбуждения по кардиомиоцитам сократительного миокарда предсердий и желудочков, то есть путём быстрой деполяризации мембраны клеток.
Скорость проведения импульса возбуждения по проводящим миоцитам предсердий и желудочков очень высока и составляет примерно 2 м/с (от 0,9 до 1,7 м/с) в предсердиях и 1-1,5 м/с в системе Гиса-Пуркинье, превышая в несколько раз скорость проведения импульса возбуждения через АВ-соединение (0,05 м/с) и скорость распространения волны возбуждения по миокарду предсердий (0,8 м/с) и желудочков (0,4 м/с)
Физиологическая задержка проведенияимпульса возбуждения в АВ-соединение,обеспечивающая синхронизированное по времени последовательное сокращение предсердий и желудочков, связана с наличием в АВ-соединении Р-клеток,обладающих автоматической активностью и определяющих более высокое сопротивление АВ-соединения проводимому импульсу возбуждения.
Длительность проведения импульса возбуждения через АВ-соединение составляет примерно 0,12-0,22 с. Зависит от частоты сердечного ритма: при синусовой тахикардии уменьшается до 0,12 с, при синусовой брадикардии увеличивается до 0,20-0,22 с.
Возбудимость.Определяется способностью кардиомиоцитов предсердий и желудочков воспринимать и распространять (по сократительному миокарду) импульсы возбуждения. Реализуется путём быстрой систолической деполяризации мембраны кардиомиоцитов в результате перемещения ионов натрия, калия, кальция и хлора через клеточную мембрану.
В состоянии электрического покояна поверхности клеточной мембраны и внутри клетки имеется неравновесная концентрация ионов натрия и калия.Ионов натрия на поверхности клетки примерно в 19 раз больше, чем внутри клетки, ионов калия, наоборот, примерно в 30 раз больше внутри клетки, чем на ее поверхности.
В состоянии покоя, благодаря избыточной концентрации положительно заряженных ионов натрияна поверхности клетки, наружная поверхность клеточной мембраныимеет положительный заряд. Внутри клеткив состоянии покоя регистрируется отрицательный заряд,что обеспечивается повышенным содержанием в клетке ионов калия,которые так изменяют геометрию внутриклеточных белков, что их отрицательные валентностивыходят на поверхностьбелковых молекул.
В покое разницы потенциалов на поверхности кардиомиоцита нет.Если в состоянии покоя с помощью микроэлектродов снять потенциал с поверхности клетки, то на электрограмме (ЭГ) одиночного мышечного волокна отклонений от изоэлектрической линии не будет. В этот период с помощью микроэлектродов можно зарегистрировать только разницу между зарядом на поверхности клетки и внутри нее.Это так называемый потенциал покоя,мощность которого колеблется от -50 до -90 мВ.
В состоянии электрического покоя клеточная мембрана непроницаема для ионов,что поддерживает высокий концентрационный градиент ионов натрия и калия с ее наружной и внутренней поверхности.Под воздействием импульса возбуждения в плазматической мембране кардиомиоцита открываются потенциал зависимые быстрые натриевые каналы,по которым в клетку по градиенту концентрации без затраты энергии перемещаются ионы натрия (быстрый потенциал-зависимый ток натрия в клетку).
Происходит изменение заряда клеточной мембраны как на ее поверхности, так и внутри (фаза деполяризации клеточной мембраны)В процессе деполяризации один полюс клетки (тот, к которому поступил импульс возбуждения) становится отрицательным, другой (противоположный) - положительным. Возникает разность потенциалов (потенциал действия),которая при записи ЭГ регистрируется как положительное, почти вертикальное отклонение от изоэлектрической линии (фаза 0 потенциала действия).
В процессе деполяризации отмечается постепенное уменьшение отрицательного значение потенциала покоя. Когда потенциал покоя снижается до - 50 мВ, в клеточной мембране открываются медленные натриевые и натрий-зависимые кальциевые каналы,по которым осуществляется медленный ток натрия и кальция внутрь клетки.
При деполяризации мембраны до - 40 мВ в клеточной мембране открываются медленные калиевые каналы,по которым калий выходит за пределы клетки. Это выходящий из клетки "задержанный" К-ток,ответственный за процесс деполяризации и реполяризации клеточной мембраны,то есть процесс восстановления исходной поляризации клетки.
В процессе деполяризации клеточной мембраны потенциал покоя быстро исчезает, то есть с -90 мВ снижается до нуля, в конце фазы деполяризации (на пике кривой ЭГ) даже становиться положительным (реверсионный потенциал),достигая примерно +20 мВ.
Быстрый натриевый ток прекращается, когда в клетку входит небольшое количество отрицательно заряженных ионов хлора.На электрограмме в этот момент регистрируется короткая отрицательная волна. Это фаза ранней быстрой реполяризацииклетки (фаза 1 потенциала действия).
Далее наступает момент, когда вся наружная поверхность клеточной мембраны становится отрицательной, а внутренняя - положительной (период обратной поляризации клетки).Разницы потенциалов на поверхности клетки почти нет, поэтому на ЭГ одиночного мышечного волокна в этот период регистрируется плато, имеющее постепенно убывающий характер (фаза медленной реполяризации или фаза 2 потенциала действия),что объясняется медленным перемещением ионов кальция, натрия и калия через клеточную мембрану (натрия и кальция с помощью кальций-натриевого обменного механизма, калия по медленным калиевым каналам)
Фаза плато плавно переходит в конечную фазу быстрой реполяризацииклеточной мембраны, когда, благодаря работе калий. натриевого насоса,восстанавливается исходная неравновесная концентрация ионов калия и натрия по обе стороны клеточной мембраны и исходный потенциал покоя.На электрограмме в этот период регистрируется плавное снижение кривой до изоэлектрической линии(фаза 3 потенциала действия).
Калий-натриевый насосклеточной мембраны представляет собой белковую молекулу, обладающую ферментативной активностью, способную расщеплять АТФ,благодаря энергии которого создается возможность перемещения ионов натрия и калия через клеточную мембрану против их концентрационного градиента.Так как процесс конечной быстрой реполяризации мембраны энергозависим,он осуществляется значительно медленнее, чем процесс деполяризации, при котором быстрый ток натрия в клетку обеспечивается концентрационным градиентом и осуществляется почти без затраты энергии.
Продолжительность потенциала действия для единичного мышечного волокна обычно не превышает 400 мл/с. Это электрическая систолакардиомиоцита, после окончания которой наступает период электрической диастолы (фаза 4 потенциала действия), когда мембрана кардиомиоцита становится, в отличие от мембраны клеток пейсмекеров, непроницаемой для ионов. Перемещения ионов через мембрану кардиомиоцитов и проводящих миоцитов в этот период нет.
Одним из основных биоэлектрических свойств сердечной мышцы является рефрактерность,то есть способность не воспринимать импульс возбуждения.Это свойство, как обратная сторона медали, связано с возбудимостью и проявляется в определенные фазы потенциала действия. Выделяют абсолютную и относительную рефрактерностьклетки (сердечной мышцы). Первая совпадает с фазой 0, 1 и 2 потенциала действия, вторая с фазой 3 потенциала действия. В начале 3 фазы (периода конечной быстрой реполяризации) возникает так называемая "экзальтационная фаза"(по Н.Е.Введенскому), когда рефрактерность на очень короткий период сменяется сверхнормальной возбудимостьюВ этот уязвимый период даже маломощный импульс возбужденияспособен вызвать повторную (преждевременную) волну возбуждения.
3.Ионные механизмы генеза биопотенциалов кардиомиоцитов
Возбудимость, проводимость и автоматия миокарда обеспечиваются электрохимическими процессами, происходящими на плазматической мембране кардиомиоцитов (сарколемме). Важнейшим параметром, характеризующим эти процессы, является электрический заряд внутренней поверхности мембраны - мембранный потенциал. Мембранный потенциал кардиомиоцитов периодически изменяется (рис.2 ). В период времени между двумя электрическими импульсами возбуждения мембранный потенциал (так называемый максимальный диастолический потенциал) достигает в разных клетках миокарда от -50 до -95 мВ. В сократительных кардиомиоцитах весь этот период мембранный потенциал стабилен и называется поэтому потенциалом покоя.
Мембранные потенциалы клеток водителей ритма во время диастолы не стабильны. Поэтому для этих клеток термин «потенциал покоя» не применяется. В момент возбуждения (генерации электрического импульса) регистрируется резкое колебание мембранного потенциала в сторону положительных величин (деполяризация), с последующим возвращением к уровню максимального диастолического потенциала. Этот электрический импульс называется потенциалом действия.
Рис.2 . Ионные градиенты и транспорт ионов через плазматическую мембрану кардиомиоцита
в скобках - концентрации ионов, ммоль/л; сплошные стрелки - активный транспорт; пунктирные стрелки - пассивный транспорт; прямоугольники - ионные каналы; заштрихованные кружки - ионные насосы; светлый кружок - Na + /Ca 2+ - переносчик.
Величина мембранного потенциала в любой момент времени определяется трансмембранными токами ионов (в основном -
калия, натрия и кальция). Скорость движения ионов через сарколемму зависит от функционирования мембранных белков-переносчиков: ионных каналов и насосов (рис.2 ).
Через ионные каналы осуществляется облегченная диффузия ионов - пассивный транспорт, не требующий затрат энергии. Направление и скорость диффузии определяются величиной электрохимического градиента для данного иона (т. е. разностью его внутри- и внеклеточной концентраций и зарядом мембраны). Большинство каналов ионоселективны, т. е. преимущественно проницаемы для какого-либо одного вида ионов,
Движение ионов против электрохимического градиента (активный транспорт) осуществляется ионными насосами, которые сопряжены с мембранными ферментами (АТФ-азами). Последние ускоряют гидролиз АТФ, а выделяющаяся при этом энергия расходуется на перенос ионов. Среди таких переносчиков наибольшую роль в кардиомиоцитах играют: K + /Na + -насос (переносит ионы Na + наружу клетки, а ионы К + - внутрь) и Са 2+ -насос (выводит ионы Са 2+ из кардиомиоцита). Основным результатом деятельности ионных насосов является создание и поддержание градиентов концентрации ионов по сторонам плазматической мембраны: снаружи значительно повышена концентрация ионов Na + и Са 2 + , а внутри - ионов К + .
В межимпульсный период проницаемость мембраны кардиомиоцитов для ионов К + значительно больше, чем для других ионов. Поэтому возникновение отрицательного диастолического потенциала определяется, в основном, пассивно выходящим из клетки током ионов калия. Другим током, участвующим в формировании диастолического потенциала кардиомиоцитов, является активный ток, непосредственно создаваемый K + /Na + -насосом. При работе этого насоса происходит неэквивалентный (электрогенный) обмен ионов: на каждые 2 иона К + , введенных в клетку, выводится 3 иона Na + . В результате возникает выходящий из клетки ток положительных зарядов - насосный ток, который увеличивает отрицательный внутриклеточный заряд. Доля насосного тока в формировании мембранного потенциала тем больше, чем больше электрическое сопротивление сарколеммы, и может составлять 25% и более (особенно в клетках Пуркинье). Активность K + /Na + - АТФ-азы и величина насосного тока регулируются концентрациями переносимых ионов, усиливаясь при увеличении внеклеточной концентрации ионов К + и внутриклеточной концентрации ионов Na + .
В клетках водителей ритма сердца диастолический потенциал нестабилен и самопроизвольно отклоняется от максимального отрицательного уровня в сторону деполяризации. Этот феномен, называемый спонтанной (медленной) диастолической деполяризацией, свойственен всем проводящим кардиомиоцитам и является основой их автоматии. Когда в результате спонтанной деполяризации мембранный потенциал достигает критического уровня (порогового потенциала), пейсмекер генерирует потенциал действия. При развитии потенциала действия в одном кардиомиоците деполяризация распространяется на соседние невозбужденные клетки. В результате мембранный потенциал этих клеток достигает порогового уровня, и в них также возникает потенциал действия. Таким образом, распространение возбуждения по миокарду происходит согласно «принципу домино»: предыдущая клетка возбуждает последующую.
В норме с наибольшей скоростью спонтанная диастолическая деполяризация протекает в пейсмекерах синоатриального узла. Поэтому в этих клетках пороговый потенциал достигается наиболее быстро, что и обусловливает самую большую частоту генерации потенциалов действия (60-80 в 1 мин). В изолированных кардиомиоцитах атриовентрикулярного соединения и проводящей системы желудочков (латентные пейсмекеры) спонтанная диастолическая деполяризация протекает с меньшей скоростью, а в сократительных кардиомиоцитах отсутствует. Таким образом, в норме перечисленные клетки возбуждаются не спонтанно, а лишь под влиянием импульсов, поступающих от синоатриального узла. Пусковыми стимулами для возбуждения кардиомиоцитов могут явиться и внешние электрические импульсы, получаемые от искусственных водителей ритма (кардиостимуляторов), а также механическое раздражение, например, сильный удар в область грудины при остановке сердца.
Основная роль в формировании потенциала действия кардиомиоцита принадлежит входящему в клетку току ионов Na + и (или) Са 2+ . Сила этого тока регулируется потенциалзависимыми ионными каналами, которые при диастолическом уровне мембранного потенциала закрыты, а при деполяризации открываются (активируются). Когда деполяризация достигает величины порогового потенциала, количество активированных каналов становится достаточным для замыкания положительной обратной связи: «деполяризация - активация ионных каналов - усиление входящего тока - деполяризация». В результате возникает самоподдерживаемый, лавинообразный вход катионов в клетку, составляющий суть формирования потенциала действия. Описанный процесс приводит к сильной деполяризации мембраны, при которой каналы входящего тока инактивируются, т. е. закрываются и временно теряют способность к активации. Сила входящего тока при этом падает, а мембранный потенциал вследствие выхода из клетки ионов К + возвращается к диастолическому уровню. В межимпульсный период каналы входящего тока вновь обретают способность к активации, а исходное распределение ионов по обе стороны сарколеммы восстанавливается за счет работы ионных насосов.
В 1975 г. американский электрофизиолог П. Крейнфилд предложил классифицировать кардиомиоциты по скорости развития фазы деполяризации потенциала действия на клетки с « медленным ответом» и клетки с «быстрым ответом». В сарколемме клеток первого типа представлены, в основном, «медленные» каналы входящего тока, которые характеризуются низкими скоростями активации и инактивации. Эти каналы проницаемы для ионов Са 2+ и Na + (в соотношении примерно 1000 : 1) и обозначаются как Ca 2+ ,Na + - или просто Са 2+ -каналы. В поверхностной мембране кардиомиоцитов с «быстрым ответом», кроме указанного вида каналов, находятся также «быстрые» натриевые каналы ,проницаемые исключительно для ионов Na + и обладающие высокими скоростями активации и инактивации. Именно свойства ионных каналов входящего тока и определяют особенности формирования потенциала действия и электрофизиологические параметры «медленных» и «быстрых» кардиомиоцитов, сравнительная характеристика которых представлена в табл.1 .
Электрофизиология сердца: мембранный (монофазный) потенциал действия
Фазы потенциала действия кардиомиоцитов (КМЦ) сердца
Трансмембранный потенциал кардиомиоцитов (КМЦ) состоит из 5 фаз: фаза 0 — возбуждение или быстрая деполяризация; фаза 1 — ранняя быстрая реполяризация; фаза 2 — плато; фаза 3 — конечная быстрая реполяризация; фаза 4 — мембранный потенциал покоя и диастолическая деполяризация. Эти фазы являются результатом пассивного тока ионов по электрохимическому градиенту, обусловленному активным ионным насосом и обменными механизмами.
Каждый ион первоначально движется через свой специфичный канал. Импульсы распространяются от одной клетки к другой без участия нервной системы. Этот факт можно наглядно продемонстрировать на трансплантированном сердце. Изложенное далее обсуждение объясняет электрогенез каждой из этих фаз.
Ионные потоки регулируют мембранный потенциал (МП) кардиомиоцитов (КМЦ) следующим образом. Когда открыт только один тип ионных каналов, предположим, что этот канал идеально селективен для данного иона, МП всей клетки будет эквивалентен РП для этого иона. При решении уравнения Nernst для 4 основных ионов но обе стороны плазматической мембраны были получены следующие равновесные потенциалы: натрий +60 мВ; калий -94 мВ; кальций +129 мВ; хлор от -80 до -35 мВ.
В связи с этим, если открывается одиночный селективный К + -канал, как, например, К + -канал, выпрямляющий входящий ток, МП становится равным Еk (-94 мВ). Если открывается одиночный селективный Na + -каиал, трансмембранный потенциал равняется ENa (+60 мВ). В покоящемся КМЦ (фаза 4) открытых калиевых каналов значительно больше, чем натриевых, при этом величина трансмембранного потенциала клетки близка Еk.
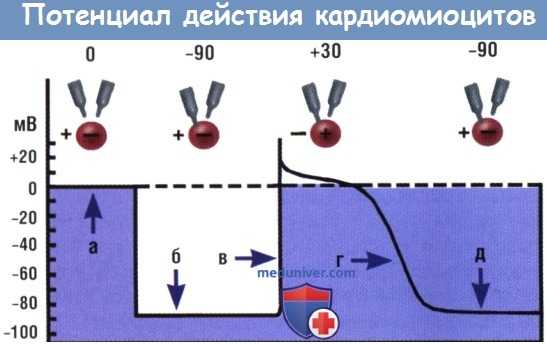
Потенциал действия, записанный во время регистрации сердечных клеток.
Клетка (сфера), два микроэлектрода и стадии: помещение электродов в клетку, активация и восстановление.
Когда оба микроэлектрода находятся вне клетки (а), разность потенциалов между ними отсутствует (потенциал 0).
Внутри клетки среда отрицательная, а вне клетки —положительная, поскольку клетка поляризована.
Один микроэлектрод проколол клеточную мембрану (б) для записи внутриклеточного мембранного потенциала покоя, который составляет -90 мВ относительно внеклеточной среды.
Клетка реполяризована (в), записано восходящее колено ПД.
ПД на пике внутри клетки составляет +30 мВ относительно внеклеточной среды.
Показана фаза реполяризации (г) с возвратом мембраны к исходному потенциалу покоя (д).
Когда одновременно открываются ионные каналы > 2 типов, каждый из них пытается привести МП к равновесному потенциалу для этого канала. Вклад каждого типа ионов в суммарный МП в каждый конкретный момент определяется мгновенной проницаемостью плазматической мембраны для этого иона. Например, по степени отклонения мембранного потенциала покоя от Еk можно предположить, что другие типы ионов с равновесным потенциалом, положительным по отношению к Еk, будут вносить вклад в величину мембранного потенциала покоя КМЦ.
Если предположить, что ионы Na + , К + и Cl - проникают через мембрану при создании потенциала покоя, индивидуальный вклад каждого из них в величину мембранного потенциала покоя (V) можно рассчитать по формуле (значения символов указаны ранее):
Если имеется только один проникающий ион, V становится равным РП для этого иона. Для нескольких типов проникающих ионов V является средневзвешенным значением всех РП.
Внутриклеточную электрическую активность можно измерить посредством помещения в одиночную клетку стеклянного микроэлектрода, заполненного раствором электролита, с наконечником диаметром < 0,5 мкм. Электрод обусловливает минимальное повреждение, точка его вхождения частично «запечатывается» клеткой.
Трансмембранный потенциал регистрируется при использовании этого электрода относительно внеклеточного электрода заземления, помещенного в культуру ткани вблизи клеточной мембраны, и представляет собой разность потенциалов между внутренней и наружной средой клетки. В качестве альтернативы в подобных условиях для измерения трансмембранного потенциала может быть использован метод пэтч-кламп.
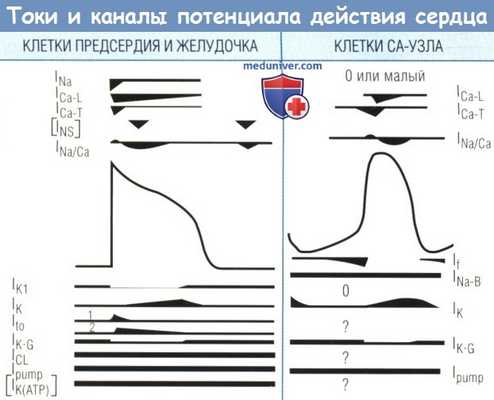
Токи и каналы, вовлеченные в генерацию ПД и потенциала покоя;
для клеток предсердия и желудочка — слева, для клеток синоатриального узла — справа.
Вверху и внизу — токи, обусловливающие электрические свойства клетки (символы и описания каналов и потоков).
Где это возможно, примерная длительность потоков, связанных с каналами или насосами, показана символично,
без попытки представить их взаимодействие друг с другом.
IK включается в состав по меньшей мере двух токов — IKR и IKS.
Оказалось, что также существует сверхбыстрый компонент IKUR.
Толстые линии для ICL, IPUMP и IK(ATP) означают только наличие этих каналов или насосов, т.к. они варьируют в зависимости от физиологических и патофизиологических условий.
Каналы в квадратных скобках (INS и IK(ATP)) активны только в патологических условиях.
INS могут представлять свеллинг-индуцируемый катионный поток.
Для клеток синусового узла (СУ) INS и IK1 небольшие или отсутствуют.
Вопросительные знаки означают еще не предоставленные экспериментальные данные для определения этих каналов в мембранах клеток СУ.
Существуют и другие механизмы ионных потоков, но они здесь не показаны, т.к. их роль в электрогенезе недостаточно определена.
Читайте также:
- Антигены псевдотуберкулеза. Антигенная структура псевдотуберкулеза. Клиника псевдотуберкулеза. Микробиологическая диагностика возбудителя псевдотуберкулеза. Реакция Фогеса-Проскауэра.
- Эффективность раннего обнаружения рака кожи.
- Мой муж перфекционист в быту. Что делать?
- КТ, МРТ при метастатической нейробластоме головы и шеи
- Примеры экстрапапиллярного типа комбинированной гамартомы сетчатки и пигментного эпителия