Функции фосфокреатина. Анаэробный механизм получения энергии
Добавил пользователь Alex Обновлено: 01.02.2026
Среди высокоэнергетических фосфори-лированных соединений имеется одно, играющее особую роль в энергетике возбудимых тканей, таких, как мышечная и нервная. Это соединение, креатинфос-фат, или фосфокреатин ( рис. 14 - 13), служит резервуаром высокоэнергетических фосфатных групп. [32]
Креатинин также является конечным продуктом азотистого обмена. Он образуется в мышечной ткани из фосфокреатина . Суточное выделение креатинина для каждого человека-величина довольно постоянная и отражает в основном его мышечную массу. У мужчин на каждый 1 кг массы тела за сутки выделяется с мочой от 18 до 32 мг креатинина, а у женщин-от 10 до 25 мг. Эти цифры мало зависят от белкового питания. В связи с этим определение суточной экскреции креатинина с мочой во многих случаях может быть использовано для контроля полноты сбора суточной мочи. [33]
Среди экстрактивных веществ, полученных из ткани мозга, найдены как азотистые, так и безазотистые соединения, которые встречаются также и в других органах и тканях, например в печени и мышцах. Из азотистых экстрактивных веществ в мозгу обнаружены: фосфокреатин , креатин, аденозинтр и фосфорная кислота, холин, аце-тилхолин, пуринов ые основания, мочевая кислота, свободные аминокислоты, гнетами н, г л ю-тамин, аспарагин и др. Мозг принадлежит к числу органов, особенно богатых креатином. Из безазотистых экстрактивных веществ в мозгу найдены глюкоза, инозит и молочная кислота. Каких-либо экстрактивных веществ, специфических только для нервной ткани, не обнаружено. [34]
При сокращении скелетной мышцы в ней снижается концентрация фосфокреатина , тогда как концентрация АТР остается практически постоянной. [35]
Также очень быстро происходил обмен в фосфопротеинах и фосфокреатине мозга . В рибонуклеиновой кислоте он был значительно более медленным: за 3 часа относительная удельная активность достигала 13 5 %, а в дезоксирибонуклеиновой кислоте она была равна лишь 2 % через 24 часа. [36]
Значение резонансной стабилизации особенно велико, когда продукты гидролиза обладают относительно высокой резонансной стабилизацией. Этот эффект приложим к ангидридам нуклеотидов с кислотами, иными, чем фосфорная кислота, такими, как карбоновые кислоты, сульфо - и аминокислоты, а также к таким фосфоамидам, как фосфокреатин , фосфоаргинин и аденозин-5 - фосфоамид. [37]
Возможность работы мышцы за счет дефосфорилирования фосфагена была установлена в 1930 г. Лундсгардом. Этот автор установил, что мышца, отравленная монойодуксусной кислотой и таким путем лишенная способности расщеплять углеводы ( стр. При этом наблюдается распад фосфокреатина в большем размере, чем в неотравленной мышце. После исчерпания запасов фосфагена и АТФ отравленная монойодацетатом мышца быстро утрачивает в анаэробных условиях способность отвечать сокращением на раздражение с нерва. [38]
Важнейшими из них, как в настоящее время установлено, являются: 1) гликогенолиз или гликолиз ( расщепление гликогена или глюкозы с образованием молочной кислоты, стр. Фосфорилирование самого креатина в мышечной ткани с образованием фосфокреатина в конечном счете осуществляется за счет энергии двух первых процессов. [39]
Ион металла может участвовать в ферментативной реакции, не оказывая существенного влияния на прочность связи фермент-субстрат. В этой реакции реагенты и ион металла взаимодействуют с ферментом независимо друг от друга, и ни один из компонентов реакции практически не влияет на прочность связи фермента с остальными компонентами. Вероятно, ион металла в данном случае выполняет роль активатора фермента и, возможно, фосфокреатина , облегчая его взаимодействие с нуклеофильным реагентом АДФ. [40]
Креатин ( метилгуанидинуксусная кислота) является обязательной составной частью поперечнополосатой мускулатуры. Содержание креатина в скелетных мышцах достигает 400 - 500 мг %, в сердечной мышце креатина в 2 - 3 раза меньше. В мышечной ткани креатин содержится как в свободном виде, так и в виде фосфорилированного производного ( креатинфосфата, фосфокреатина ), который образуется в результате обратимого переноса фосфорильного остатка с АТФ на креатин. [41]
Итак, мы видим, что системы фосфокреатина и анаэробного дыхания поставляют энергию быстро, но только в течение короткого времени. Аэробная система способна служить источником энергии неограниченно долго при достаточном количестве дыхательного субстрата. В таких видах спорта, которые рассчитаны на короткое и резкое усиление мышечной активности, например в беге на короткую дистанцию или в поднятии штанги, энергию поставляет главным образом система фосфокреатина . При беге на 200 м анаэробное дыхание может служить дополнительным источником энергии. При беге на 400 м оно поставляет уже большую часть энергии, а при таких играх, как теннис, сквош или футбол, практически вся энергия в момент предельного напряжения поступает от этой системы. Те виды спорта, в которых главное - выносливость, например марафон, бег трусцой или бег на лыжах по пересеченной местности, зависят почти целиком от аэробного дыхания. [42]
Содержащегося в мышцах АТФ может хватить при максимальной мышечной нагрузке не более, чем примерно на три секунды. Фосфокреатин - это еще одно вещество, из которого при отщеплении его фосфатной группы высвобождается энергия. Этой энергии достаточно для синтеза АТФ из АДФ и Фн. Запас фосфокреатина в мышцах в 2 - 4 раза превышает запас АТФ, и при необходимости энергия из фосфокреатина может быть быстро передана АТФ. [43]
Содержащегося в мышцах АТФ может хватить при максимальной мышечной нагрузке не более, чем примерно на три секунды. Фосфокреатин - это еще одно вещество, из которого при отщеплении его фосфатной группы высвобождается энергия. Этой энергии достаточно для синтеза АТФ из АДФ и Фн. Запас фосфокреатина в мышцах в 2 - 4 раза превышает запас АТФ, и при необходимости энергия из фосфокреатина может быть быстро передана АТФ. [44]
Запасы гликогена в мышцах, однако, невелики, и потому существует верхний предел того количества энергии, которое вырабатывается в ходе гликолиза, в условиях максимальной ( например, при спринте) нагрузки. Более того, накопление молочной кислоты и связанное с этим снижение рН, а также повышение температуры, происходящее при очень высокой мышечной активности, снижают эффективность обмена в мышцах. Так, в период восстановления после максимальной мышечной нагрузки атлет продолжает еще некоторое время тяжело дышать. Потребляемый при этом дополнительный кислород используется для окисления пирувата, лактата и других субстратов, а также регенерации АТР и фосфокреатина в мышцах. [45]
34.Брожение. Получение энергии анаэробными микроорганизмами. Химизм процесса.
Брожение (субстратное фосфорилирование) - разновидность анаэробного дыхания, при котором и донором и акцептором водорода является органическое вещество. При анаэробном дыхании микроорганизмы получают энергию не путем окисления, а путем распада сложных органических веществ до более простых. Микроорганизмы, у которых имеет место анаэробное дыхание, называют анаэробами.
При брожении происходит расщепление сложных органических веществ до более просто устроенных с выделением небольшого количества энергии. При поступлении глюкозы в клетку, происходит гликолиз и образуется ПВК. Дальнейшие ее превращения зависят от набора ферментов анаэробных бактерий. В зависимости от того какие конечные продукты образуются, выделяют разные типы брожения:
· Молочнокислое брожение вызывается лактобактериями, бифидобактериями, стрептококками, образуя из ПВК молочную кислоту (гомоферментативное брожение) или молочную, янтарную, уксусную кислоты, ацетон (гетероферментативное брожение). Эти бактерии применяются в производстве молочно-кислых продуктов: ряженки, простокваши, кефира, йогуртов и творога.
· Маслянокислое брожение. Возбудителями этого вида брожения являются анаэробные бактерии рода клостридии, а также бактероиды, фузобактерии и другие микроорганизмы, вызывающие у человека опасные анаэробные инфекции. Основным продуктом брожения является масляная,изомасляная, уксусная, валериановая кислоты.
· Пропионовокислое брожение также вызывается анаэробами - пропионибактериями (обитатели кожи и слизистой оболочкичеловека и животных могут вызывать анаэробные инфекции), которые используются в производстве сыров. Конечный продукт брожения - пропионовая кислота.
· Спиртовое брожение. Вызывают дрожжи. В результате спиртового брожения образуется этиловый спирт, что издавна используется в пивоварении и виноделии.
· Бутиленгликолевое брожение. В результате брожения образуются бутиловый спирт, этиленгликоль, срероводород и другие токсические продукты. Этот вид брожения вызывают кишечная палочка и другие энтеробактерии, в том числе - возбудители кишечных инфекций - сальмонеллёза, дизентерии.
При субстратном фосфорилировании из глюкозы или других источников углерода выделяется незначительное количество энергии, так как образующиеся при этом продукты брожения (молочная кислота, спирты и др.) сохраняют в себе значительные количества энергии. Поэтому в анаэробных условиях бактериальная культура для получения необходимой энергии во много раз больше разлагает пищевого материала, чем в присутствии кислорода. Теплообразование при развитии бактериальной флоры в органическом материале (навоз, торф, мусор) может привести к его самовозгоранию.
Изучение ферментов бактерий имеет большое практическое значение для разработки методов диагностики (идентификации) возбудителей инфекционных заболеваний по набору ферментов, а также для создания современных биотехнологий получения продуктов питания в том числе молочнокислых продуктов, сыра, хлеба, вина, пива и т.д.
Химизм процесса брожения был расшифрован в 1933 г. в результате независимых исследований немецкого ученого Г. Г. Эмбдена, немецко-американского — О.Ф. Мейергофа и польско-советского — Я.О. Парнаса.
Было установлено, что брожение представляет собой последовательную цепочку реакций, протекающих в цитоплазме клетки бродящего организма под воздействием вырабатываемых здесь ферментов. В процессе анаэробной диссимиляции глюкозы различают три стадии: подготовительную, энергетическую и завершающую.
«Пусковой» реакцией катаболического распада глюкозы является ее фосфорилирование за счет АТФ с образованием глюкозо-6-фос- фата и АДФ, в результате чего молекула глюкозы активируется для участия в последующих превращениях. Катализирует реакцию фермент гексокиназа (КФ 2.7.1.1).
Затем образовавшийся глюкозо-6-фосфат (альдоза) превращается во фруктозо-6-фосфат (кетозу). Эта реакция происходит под воздействием фермента глюкозо-6-фосфат—изомеразы (КФ 5.3.1.9).
Третья реакция — фосфорилирование фруктозо-6-фосфата за счет АТФ с образованием фруктозо-1,6-бисфосфата и АДФ. Она катализируется ферментом фосфофрукгокиназой (КФ 2.7.1.11) и представляет собой еще одну «пусковую» реакцию брожения.
Четвертая реакция процесса брожения — расщепление фруктозо- 1,6-бисфосфата и образование двух молекул фосфотриоз: фосфоди- гидроксиацетона (ФДА) и 3-фосфоглицеринового альдегида (3-ФГА). Фермент альдолаза (КФ 4.1.2.13) катализирует разрыв углеродной цепочки фруктозо-1,6-бисфосфата посередине, чему способствует
Энергетическая стадия брожения также включает пять реакций, в ходе которых энергия, содержащаяся в глюкозе, постепенно высвобождается и запасается в форме АТФ (рис. 61).
Она начинается с того, что 3-фосфоглицериновый альдегид под воздействием фермента триозофосфатдегидрогеназы (КФ 1.1.1.12) при участии фосфорной кислоты превращается в 1,3-бисфосфогли- цериновую кислоту (1,3-бисФГК). Энергия, высвобождающаяся в результате окисления альдегидной группы 3-фосфоглицеринового альдегида, аккумулируется в образуемой высокоэнергетической связи, с помощью которой присоединяется фосфатная группа.
Затем фермент фосфоглицераткиназа (КФ 2.7.2.3) катализирует перенос остатка фосфорной кислоты, содержащего высокоэнергетическую связь, от карбоксильной группы 1,3-бисфосфоглицерино- вой кислоты на АДФ с образованием АТФ и 3-фосфоглицериновой кислоты (3-ФГК).
В результате этой реакции энергия, ранее высвободившаяся при окислении альдегидной группы 3-фосфоглицеринового альдегида до карбоксильной, оказывается запасенной в виде АТФ.
После этого 3-фосфоглицериновая кислота под воздействием фермента фосфоглицерат-фосфомутазы (КФ 5.4.2.1) превращается в 2-фосфоглицериновую кислоту (2-ФГК).
Образовавшаяся 2-фосфоглицериновая кислота под воздействием фермента енолазы (КФ 4.2.1.11) превращается в 2-фосфоенолпиро- виноградную кислоту (2-ФЕП). При этом молекула 2-фосфоглице- риновой кислоты отдает молекулу воды. Это вызывает перераспределение энергии внутри молекулы субстрата, и связь, с помощью которой к нему присоединен остаток фосфорной кислоты, становится высокоэнергетической.
На последнем этапе энергетической стадии брожения 2-фосфо- енолпировиноградная кислота с помощью фермента пируваткиназы (КФ 2.7.1.40) передает содержащий высокоэнергетическую связь остаток фосфорной кислоты молекуле АДФ, в результате чего образуются АТФ и енолпировиноградная кислота (енолПВК). Последняя весьма нестойка и быстро неферментативным путем превращается в устойчивую кетоформу — пировиноградную кислоту (ПВК).
Таким образом, в ходе брожения молекула глюкозы расщепляется ферментативным путем в десяти последовательных реакциях до двух молекул пировиноградной кислоты и двух пар атомов водорода.
Образовавшаяся в результате реакций гликолиза пировиноград- ная кислота может подвергаться дальнейшим превращениям, направление которых зависит от условий среды (аэробных или анаэробных) и от специфических особенностей данного организма. В аэробных условиях она может окисляться до диоксида углерода и воды в процессе дыхания, а при анаэробной диссимиляции последующие превращения пировиноградной кислоты составляют завершающий этап брожения.
3.12.1. Креатинфосфокиназный механизм ресинтеза атф
Алактатный анаэробный механизм ресинтеза АТФ включает использование имеющейся в мышцах АТФ и быстрый ее ресинтез за счет креатинфосфата, концентрация которого в мышцах в 3-4 раза выше по сравнению с АТФ. Креатинфосфат локализован на сократительных миофибриллах.
Креатинфосфат + AДФ = АТФ +Креатин
Обратная реакция может протекать в митохондриях с использованием АТФ, образовавшейся в процессе окислительного фосфорилирования. Мембрана митохондрий хорошо проницаема как для креатина, так и для креатинфосфата, а креатинфосфокиназа есть и в саркоплазме, и в межмембранном пространстве митохондрий.
В скелетных мышцах креатинфосфокиназа обладает высокой активностью, что приводит к усилению этой реакции в самом начале мышечной работы, когда начинает расщепляться АТФ и накапливаться АДФ. Максимальная мощность креатинфосфокиназной реакции развивается уже на 0,5-0,7 с интенсивной работы, что свидетельствует о большой скорости развертывания, и поддерживается в течение 10-15 с у нетренированных, а у высокотренированных спринтеров - 25-30 сек.
Креатинфосфокиназный механизм первым включается в процесс ресинтеза АТФ в начале интенсивной мышечной работы и протекает до тех пор, пока не исчерпаются запасы креатинфосфата. Максимальная мощность креатинфосфокиназной реакции в 1,5-2 раза выше мощности анаэробного гликолиза и в 3-4 раза аэробного процесса. Общие запасы креатинфосфата у нетренированных обеспечивают образование энергии в количестве 420 кДж/кг, а тренированных - в 2 раза больше. Метаболическая емкость невелика. Эффективность очень высокая. Запасы креатинфосфата зависят от запасов креатина. Введение креатина в виде добавок приводит к увеличению запасов креатинфосфата, а также к повышению работоспособности. Содержание креатинфосфата увеличивается при адаптации организма к скоростным и силовым нагрузкам в 1,5-2 раза.
Креатинфосфокиназный путь ресинтеза АТФ играет решающую роль в энергообеспечении кратковременной работы максимальной интенсивности в течение 15-30 с - бег на 100 м, плавание на короткие дистанции, прыжки, метания, тяжелая атлетика и др. Он обеспечивает возможность быстрого перехода от покоя к работе, внезапных изменений темпа по ходу ее выполнения, а также финишного ускорения. Функционирует креатинфосфокиназная система преимущественно в быстросокращающихся мышечных волокнах, поэтому составляет биохимическую основу скорости и локальной мышечной силы (выносливости).
3.12.2. Гликолитический механизм ресинтеза атф
Как только креатинфосфокиназа перестает обеспечивать необходимую скорость восстановления АТФ в мышцах, в энергообеспечение работы вовлекается анаэробный гликолитический механизм ресинтеза АТФ. В процессе гликолиза используются внутримышечные запасы гликогена и глюкоза крови. Они постепенно расщепляются до лактата. Активации гликолиза способствует также снижение концентрации креатинфосфата в мышцах и накопление аденозинмонофосфата (АМФ), образующейся в миокиназной реакции.
Максимальная мощность гликолиза у хорошо тренированных людей 3,1 кДж/кг, у нетренированных - 2,5 кДж/кг. Это ниже мощности креатинфосфокиназы, но в 2-3 раза выше аэробного процесса. На максимальную мощность механизм выходит на 20-30 сек после начала работы. К концу 1-ой минуты работы гликолиз становится основным процессом ресинтеза АТФ. При дальнейшей работе снижается под влиянием образования лактата и снижения рН. Обеспечивает поддержание анаэробной работы продолжительностью от 30 сек до 2-6 мин. Общее количество энергии, которое образуется в гликолитическом механизме у нетренированных людей - 840 Дж/кг, у спортсменов - 1760-2090 Дж/кг. Гликолитический механизм отливается невысокой эффективностью. При гликолитической работе повышается теплопродукция, температура мышц может повышаться до 41-42С.
Гликолиз играет важную роль при напряженной мышечной работе в условиях недостаточного снабжения тканей кислородом. Это основной путь образования энергии в упражнениях субмаксимальной мощности, предельная продолжительность которых составляет от 30 сек до 2,5 мин - бег на средние дистанции, плавание на 100-200 м, велосипедные гонки на треке и др. За счет гликолиза совершаются длительные ускорения по ходу упражнения и на финише дистанции. Гликолитический механизм энергообразования является биохимической основой специальной скоростной выносливости организма.
Умеренный сдвиг рН в кислую сторону активирует работу ферментов дыхательного цикла в митохондриях и усиливает аэробное энергообразование. Изменение рН от 7,1 до 6,5 при изнеможении угнетает ферменты гликолиза и сокращение мышц. При рН 6,4 прекращается расщепление гликогена, что вызывает резкое снижение уровня АТФ и развитие утомления. Увеличение лактата в мышцах сопровождается изменением осмотического давления. Вода поступает в мышцы и они набухают, возникают болевые ощущения.
Увеличение кислотности крови активирует дыхательный центр, в результате чего увеличивается легочная вентиляция и поставка кислорода к работающим мышцам. Все это происходит при увеличении интенсивности выполняемого упражнения более максимальной аэробной мощности. Этот уровень нагрузки - порог анаэробного обмена (ПАНО), или порог лактата. Величина ПАНО - показатель эффективности процессов энергообразования в мышцах, который используется при биохимическом контроле функционального состояния спортсменов.
9.2.1. Анаэробный гликолиз
Последовательность реакций анаэробного гликолиза представлена на рисунке 9.2:
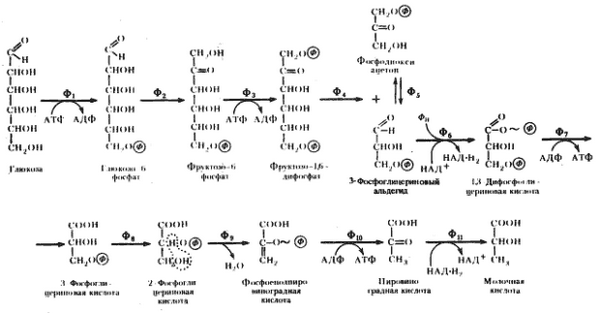
Рис. 9.2. Схема гликолиза в анаэробных условиях:Ф1 — гексокиназа; Ф2 — глюкозофосфатизомераза; Ф3 — фофсофруктокиназа; Ф4 — фруктозо-1,6-дифосфатальдолаза; Ф5 — триозофосфатизомераза; Ф6 — 3-ФГА-дегидрогеназа; Ф7 — фофсоглицерокиназа; Ф8 — фосфоглицеромутаза; Ф9 — енолаза; Ф10 — пируваткиназа; Ф11 — лактатдегидрогеназа
Условно гликолиз можно разделить на две стадии:
- первая стадия гликолиза - стадия активации глюкозы, которая включает пять реакций и завершается расщеплением углеродного скелета глюкозы на две молекулы трёхуглеродного скелета - глицероальдегидфосфата;
- вторая стадия - синтез молекул АТФ, в которой энергия окислительных реакций трансформируется в химическую энергию АТФ по механизму субстратного фосфорилирования.
I. Стадия активации глюкозы:
1) Необратимая реакция фосфорилирования глюкозы и образования глюкозо-6-фосфата, катализируемая ферментом гексокиназой.
2) Обратимая реакция кето-альдольной изомеризации глюкозо-6-фосфата во фруктозо-6-фосфат, катализируемая ферментом глюкозо-6-фосфатизомеразой.
3) Необратимая реакция фосфорилирования фруктозо-6-фосфата молекулой АТФ до фруктозо-1,6-дифосфата, катализируемая ферментом фосфофруктокиназой.
4) Обратимая реакция расщепления связи С-С во фруктозо-1,6- дифосфате на две триозы дигидрооксиацетон-3-фосфат и глицеральдегид-3-фосфат, катализируемые ферментом альдолазой.
5) Обратимая реакция кето-альдольной изомеризации дигидроксиацетон-3-фосфата в глицеральдегид-3-фосфат, катализируемая ферментом триозофосфатизомеразой.
II. Стадия синтеза атф:
6) Обратимая реакция окисления глицероальдегид-3-фосфата до 1,3-фосфоглицерата, которая катализируется ферментом глицеральдегид-3-фосфатдегидрогеназой.
. Эта реакция позволяет высокую энергию окисления альдегидной группы аккумулировать в виде макроэргической связи АТФ.
7) Обратимая реакция субстратного фосфорилирования АДФ и образования АТФ, при которой происходит перенос богатого энергией фосфорильного остатка с 1,3-дифосфоглицерата на АДФ. Реакция катализируется фосфоглицераткиназой.
8) Обратимая реакция изомериизации 3-фосфоглицерата в 2-фосфоглицерат, катализируемая ферментом фосфоглицератмутазой.
9) Обратимая реакция енолизации, в процессе которой отщепление молекулы воды от 2-фосфоглицерата приводит к образованию макроэргической связи в фосфоеноилпирувате. Реакция катализируется ферментом енолазой.
10) Необратимая реакция субстратного фосфорилирования АДФ и образования АТФ, при которой происходит разрыв макроэргитечской связи и перенос фосфорильного остатка от фосфоэнолпирувата на АДФ. Катализируется эта реакция ферментом пируваткиназой.
11) Обратимая реакция восстановления пирувата до лактката происходит в анаэробных условиях при участии фермента лактатдегидрогеназы, коферментом которой является восстановленная форма НАДН∙Н +
Специфические характеристики анаэробного гликолиза (отличие от аэробного гликолиза):
Во-первых, при анаэробном гликолизе окисление НАДН∙Н + осуществляется независимо от дыхательной цепи. В этом случае акцептором водорода от НАДН∙Н + является пируват, который восстанавливается в лактат.
Во-вторых, образование АТФ при анаэробном гликолизе идёт за счет субстратного фосфорилирования, когда для фосфорилирования АДФ используется энергия макроэргической связи субстрата (реакции 7, 10).
Ресинтез АТФ
Дано определение ресинтеза АТФ. Описаны основные пути ресинтеза АТФ в мышечных волокнах: креатинфосфатный, гликолитический, миокиназный и тканевое дыхание. Описаны количественные критерии путей ресинтеза АТФ, соотношение между различными путями ресинтеза АТФ при мышечной работе, а также между путями ресинтеза АТФ и зонами относительной мощности.
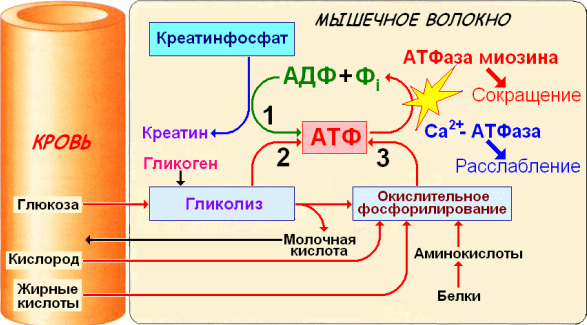
Ресинтез АТФ в мышечных волокнах
Определение
Ресинтез АТФ - синтез АТФ из различных энергетических субстратов во время физической работы в мышечных волокнах.
Формула ресинтеза АТФ выглядит следующим образом:
Пути ресинтеза АТФ
Ресинтез АТФ может осуществляться двумя путями:
- с участием кислорода (аэробный путь).
- без участия кислорода (анаэробный путь);
Аэробный путь (тканевое дыхание, аэробное или окислительное фосфорилирование) - основной способ образования АТФ в мышечных волокнах. Он протекает в митохондриях мышечных волокон. В результате тканевого дыхания выделяется 39 молекул АТФ. Окисляемое вещество распадается до углекислого газа и воды.
Анаэробный ресинтез АТФ
Анаэробные пути ресинтеза АТФ являются дополнительными способами образования АТФ в мышечных волокнах в тех случаях, когда основной путь получения АТФ - тканевое дыхание не может обеспечить мышечную деятельность необходимым количество кислорода. Эти механизмы ресинтеза АТФ активно функционируют в начале выполнения физических упражнений, когда тканевое дыхание не полностью «развернулось», а также при физических нагрузках высокой мощности.
Анаэробный ресинтез АТФ в мышечных волокнах возможен посредством нескольких механизмов:
- - ресинтез АТФ из креатинфосфата; - ресинтез АТФ из гликогена мышц;
- Миокиназный (аденилаткиназный) ресинтез АТФ - ресинтез АТФ из АДФ при значительном накоплении в мышечных волокнах АДФ. Рассматривается как аварийный механизм, обеспечивающий ресинтез АТФ, когда другие пути ресинтеза АТФ невозможны.
Количественные критерии путей ресинтеза АТФ
Существуют количественные критерии путей ресинтеза АТФ. К ним можно отнести: максимальную мощность, время развертывания, время сохранения или поддержания максимальной мощности, метаболическую ёмкость (табл. 1).
- Максимальная мощность - максимальное количество АТФ, которое может образоваться в единицу времени при функционировании данного пути ресинтеза АТФ.
- Время развертывания - минимальная длительность, необходимая для выхода ресинтеза АТФ на свою максимальную мощность.
- Время сохранения или поддержания максимальной скорости - длительность функционирования данного пути ресинтеза АТФ с максимальной мощностью.
- Метаболическая ёмкость - количество АТФ, которое может образоваться во время мышечной работы за счёт данного пути ресинтеза АТФ.
Таблица 1. Количественные критерии основных путей ресинтеза АТФ (С.С. Михайлов, 2009)
Соотношение между различными путями ресинтеза АТФ
При любой мышечной работе функционируют все три основных механизма ресинтеза АТФ, но включаются они последовательно. В первые секунды ресинтез АТФ осуществляется за счет креатинфосфатной реакции, затем включается гликолиз. По мере продолжения работы на смену гликолизу приходит тканевое дыхание (рис.1). Эта смена механизмов ресинтеза АТФ приводит к уменьшению суммарной выработки АТФ.
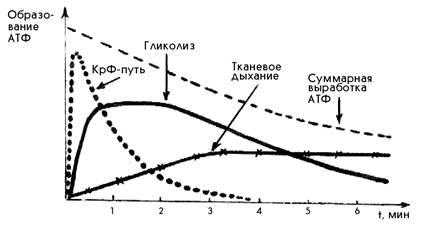
Рис.1. Включение путей ресинтеза АТФ при выполнении физической работы (С.С. Михайлов, 2009)
Пути ресинтеза АТФ и зоны относительной мощности
В.С. Фарфель приводит следующее соотношение мощности работы и основной системы энергообеспечения (табл.2)
Таблица 2. Зоны мощности работы и основная система энергообеспечения (В.С. Фарфель)
| Мощность работы | Основная система энергообеспечения | Типичное время работы |
| Максимальная | Креатинфосфатная реакция | до 20 с |
| Субмаксимальная | Гликолиз | до 5 мин. |
| Большая | Гликолиз+ тканевое дыхание | до 30 мин. |
| Умеренная | Тканевое дыхание | Более 30 мин. |
J.T. Cramer (2008) приводит несколько иное соотношение зон мощности и основных систем энергообеспечения (табл.3)
Таблица 3. — Зоны относительной мощности и основная ситема энергообеспечения (J.T. Cramer, 2008)
Читайте также: