Передача тактильных сигналов в нервных волокнах. Восприятие вибрации
Добавил пользователь Алексей Ф. Обновлено: 21.01.2026
В соматосенсорную систему входят система кожной чувствительности и чувствительная система скелетно-мышечного аппарата, главная роль в которой принадлежит проприорецепции.
5.1. Кожная рецепция
Кожные рецепторы сосредоточены на огромной кожной поверхности (1,4-2,1 м 2 ). В коже находится множество рецепторов, чувствительных к прикосновению, давлению, вибрации, теплу и холоду, а также к болевым раздражениям. Они весьма различны по строению, локализуются на разной глубине кожи и распределены неравномерно по её поверхности. Больше всего их в коже пальцев рук, ладоней, подошв, губ и половых органов. У человека в коже с волосяным покровом (90% всей кожной поверхности) основным типом рецепторов являются свободные окончания нервных волокон, идущих вдоль мелких сосудов, а также более глубоко локализованные разветвления тонких нервных волокон, оплетающих волосяную сумку. Эти окончания обеспечивают высокую чувствительность волос к прикосновению. Рецепторами прикосновения являются также осязательные мениски (диски Меркеля), образованные в нижней части эпидермиса контактом свободных нервных окончаний с модифицированными эпителиальными структурами. Их особенно много в коже пальцев рук.
В коже, лишённой волосяного покрова, находят много осязательных телец (телец Мейснера). Они локализованы в сосочковом слое кожи пальцев рук и ног, ладонях, подошвах, губах, языке, половых органах и сосках груди. Другими инкапсулированными нервными окончаниями, но более глубоко расположенными, являются пластинчатые тельца, или тельца Пачини (рецепторы давления и вибрации). Они имеются также в сухожилиях, связках, брыжейке.
Теории кожной чувствительности многочисленны и во многом противоречивы. Наиболее распространено представление о наличии специфических рецепторов для четырёх основных видов кожной чувствительности: тактильной, тепловой, холодовой и болевой. Исследования электрической активности одиночных нервных окончаний и волокон свидетельствуют о том, что многие из них воспринимают лишь механические или температурные стимулы.
Механизмы возбуждения кожных рецепторов. Механический стимул приводит к деформации мембраны рецептора. В результате этого электрическое сопротивление мембраны уменьшается, т.е. увеличивается её проницаемость для ионов. Через мембрану рецептора начинает течь ионный ток, приводящий к генерации рецепторного потенциала. При достижении рецепторным потенциалом критического уровня деполяризации генерируются импульсы, распространяющиеся по волокну в ЦНС.
Адаптация кожных рецепторов. По скорости адаптации при длящемся действии раздражителя большинство кожных рецепторов подразделяются на быстрои медленноадаптирующиеся. Наиболее быстро адаптируются тактильные рецепторы, расположенные в волосяных фолликулах, а также пластинчатые тельца. Адаптация кожных механорецепторов приводит к тому, что мы перестаём ощущать постоянное давление одежды или привыкаем носить на роговице глаз контактные линзы.
5.2. Свойства тактильного восприятия
Ощущение прикосновения и давления на кожу довольно точно локализуется человеком на определённом участке кожной поверхности. Эта локализация вырабатывается и закрепляется в онтогенезе при участии зрения и проприорецепции. Абсолютная тактильная чувствительность существенно различается в разных частях кожи: от 50 мг до 10 г. Пространственное различение на кожной поверхности, т.е. способность человека раздельно воспринимать прикосновения к двум соседним точкам кожи, также сильно отличается в разных её участках. На языке порог пространственного различия равен 0,5 мм, а на коже спины более 60 мм. Эти отличия связаны главным образом с различными размерами кожных рецептивных полей (от 0,5 мм 2 до 3 см 2 ) и со степенью их перекрытия [Кейдель, 1975; Физиол. сенсорн. систем, 1971, 1972, 1975].
5.3. Температурная рецепция
Температура тела человека колеблется в сравнительно узких пределах. Именно поэтому информация о температуре внешней среды, необходимой для деятельности механизмов терморегуляции, имеет особо важное значение. Терморецепторы располагаются в коже, на роговице глаза, в слизистых оболочках, а также в гипоталамусе. Они подразделяются на два вида: холодовые и тепловые (последних намного меньше). Больше всего терморецепторов в коже лица и шеи. Полагают, что терморецепторами могут быть немиелинизированные окончания дендритов афферентных нейронов.
Терморецепторы подразделяются на специфические и неспецифические. Первые отвечают лишь на температурное воздействие, вторые реагируют и на механическое раздражение. Рецептивные поля большинства терморецепторов локальны. Терморецепторы реагируют на изменение температуры повышением частоты импульсов, устойчиво длящимся в течение всего времени действия стимула. Это повышение пропорционально изменению температуры, причём постоянная частота разрядов у тепловых рецепторов наблюдается в диапазоне температур от +20 до +50°С, а у холодовых от +10 до +41°С. Дифференциальная чувствительность терморецепторов велика: достаточно изменить температуру на 0,2°С, чтобы вызвать длительные изменения их импульсации.
Температурное ощущение человека зависит как от абсолютного значения температуры, так и от разницы температуры кожи и действующего раздражителя, его площади и места приложения. Так, если руку держали в воде с температурой +27°С, то в первый момент после переноса руки в воду, нагретую до +25°С, она кажется холодной. Однако уже через несколько секунд становится возможной оценка абсолютной температуры.
5.4. Болевая рецепция
Болевая, или ноцицептивная, чувствительность имеет особое значение для выживания организма, так как сигнализирует о действии чрезмерно сильных и вредных факторов. В симптомокомплексе многих заболеваний боль одно из первых, а иногда и единственное проявление патологии и важный показатель для диагностики. Несмотря на интенсивные исследования, до сих пор не удаётся решить вопрос о существовании специфических болевых рецепторов и адекватных им болевых раздражителей. Сформулированы две альтернативные гипотезы об организации болевого восприятия: а) существуют специфические болевые рецепторы (свободные нервные окончания с высоким порогом реакции) и б) специфических болевых рецепторов не существует, и боль возникает при сверхсильном раздражении любых рецепторов.
В электрофизиологических опытах на одиночных нервных волокнах типа С обнаружено, что некоторые из них реагируют преимущественно на чрезмерные механические, а другие на чрезмерные тепловые воздействия. При болевых раздражениях импульсы возникают также в волокнах типа А. Соответственно разной скорости проведения импульсов в волокнах типа С и А отмечается двойное ощущение боли: вначале чёткое по локализации и короткое, а затем длительное, разлитое и сильное (жгучее).
Механизм возбуждения рецепторов при болевых воздействиях пока не выяснен. Предполагают, что значимыми являются изменения рН ткани в области нервного окончания, так как этот фактор обладает болевым эффектом при концентрациях водородных ионов, встречающихся в реальных условиях. Таким образом, наиболее общей причиной возникновения боли можно считать изменение концентрации этих ионов при токсическом воздействии на дыхательные ферменты или при механическом или термическом повреждении клеточных мембран.
Адаптация болевых рецепторов возможна: ощущение укола от продолжающей оставаться в коже иглы быстро проходит. Однако в очень многих случаях болевые рецепторы не обнаруживают существенной адаптации, что делает страдания больного особенно длительными и требует применения анальгетиков.
Болевые раздражения вызывают ряд рефлекторных соматических и вегетативных реакций. Если эти реакции умеренно выражены, то они имеют приспособительное значение, но могут привести к тяжёлым патологическим эффектам, например к шоку. При болевых воздействиях на кожу человек локализует их достаточно точно, но при заболеваниях внутренних органов часты так называемые отражённые боли, проецирующиеся в определённые части кожной поверхности (зоны Захарьина-Геда). Так, при стенокардии кроме болей в области сердца ощущается боль в левой руке и лопатке. При тактильных, температурных и болевых раздражениях «активных» точек кожной поверхности (иглоукалывание или акупунктура) включаются цепи рефлекторных реакций, опосредуемых центральной и вегетативной нервной системой. Они могут избирательно изменять кровоснабжение и трофику тех или иных органов и тканей.
5.5. Мышечная и суставная рецепция (проприорецепция)
В мышцах человека содержатся три типа специализированных рецепторов: первичные окончания веретён, вторичные окончания веретён и сухожильные рецепторы Гольджи. Эти рецепторы реагируют на механические раздражения и участвуют в координации движений, являясь источником информации о состоянии двигательного аппарата (см. также гл. 5).
Мышечное веретено имеет длину в несколько миллиметров, ширину в десятые доли миллиметра, одето капсулой и расположено в толще мышцы. Внутри капсулы находится пучок интрафузальных мышечных волокон. Веретена расположены параллельно внешним по отношению к капсуле экстрафузальным волокнам, поэтому при растяжении мышцы нагрузка на веретена увеличивается, а при сокращении уменьшается. В расслабленной мышце импульсация, идущая от веретён, невелика, но они реагируют повышением частоты разрядов на удлинение мышцы. Таким образом, веретена дают мозгу информацию о длине мышцы и её изменениях. Импульсация, идущая от веретён, в спинном мозге возбуждает мотонейроны своей мышцы и тормозит мотонейроны мышцы-антагониста, а также возбуждает мотонейроны сгибателей и тормозит мотонейроны разгибателей.
Сухожильные рецепторы Гольджи находятся в зоне соединения мышечных волокон с сухожилием и расположены последовательно по отношению к мышечным волокнам. Они слабо реагируют на растяжение мышцы, но возбуждаются при её сокращении, причём их импульсация пропорциональна силе сокращения. Поэтому сухожильные рецепторы информируют мозг о силе, развиваемой мышцей. Идущие от этих рецепторов волокна в спинном мозге вызывают торможение мотонейронов собственной мышцы и возбуждение мотонейронов мышцы-антагониста. Информация от мышечных рецепторов по восходящим путям спинного мозга поступает в высшие отделы ЦНС, включая кору большого мозга.
Суставные рецепторы изучены меньше, чем мышечные. Известно, что они реагируют на положение сустава и на изменения суставного угла, участвуя таким образом в системе обратных связей от двигательного аппарата.
5.6. Передача и переработка соматосенсорной информации
Чувствительность кожи и ощущение движения связаны с проведением в мозг сигналов от рецепторов по двум основным путям (трактам): лемнисковому и спиноталамическому, значительно различающимся по своим свойствам.
Лемнисковый путь передаёт в мозг сигналы о прикосновении к коже, давлении на неё и движениях в суставах. Отличительная особенность этого пути быстрая передача в мозг наиболее точной информации, дифференцированной по силе и месту воздействия.
По мере перехода на всё более высокие уровни изменяются некоторые важные свойства нейронов лемнискового пути. Значительно увеличиваются рецептивные поля нейронов (в продолговатом мозге в 2-30, а в коре большого мозга в 15-100 раз). Несмотря на увеличение размеров рецептивных полей, нейроны остаются достаточно специфичными (нейроны поверхностного прикосновения, глубокого прикосновения, нейроны движения в суставах и нейроны положения или угла сгибания суставов). Для корковой части лемнискового пути характерна чёткая топографическая организация, т.е. проекция кожной поверхности осуществляется в центры мозга по принципу «точка в точку» [Кейдель, 1975; Сомьен, 1975; Дудел и др., 1985]. При этом площадь коркового представительства той или иной части тела определяется её функциональной значимостью: формируется так называемый «сенсорный гомункулюс» (рис. 4.10). Роль соматосенсорной коры состоит в интегральной оценке соматосенсорных сигналов, включении их в сферу сознания и в сенсорное обеспечение выработки новых двигательных навыков.
Спиноталамический путь значительно отличается от лемнискового сравнительно медленной передачей афферентных сигналов, нечётко дифференцированной информацией о свойствах раздражителя и не очень чёткой её топографической локализацией; он служит для передачи температурной, всей болевой и в значительной мере тактильной чувствительности.
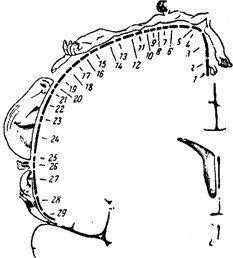
Рис. 4.10. Корковое представительство кожной чувствительности. Расположение в соматосенсорной зоне коры больших полушарий мозга человека проекций различных частей тела:
1- половые органы; 2 пальцы ноги; 3 ступня; 4 голень; 5 бедро; 6 туловище; 7- шея; 8 голова; 9 плечо; 10-11 локоть; 12 предплечье; 13 запястье; 14 кисть; 15-19 пальцы руки; 20 глаза; 21 нос; 22 лицо; 23 верхняя губа; 24,26 зубы; 25 нижняя губа; 27 язык; 28 глотка; 29 внутренние органы. Размеры изображений частей тела соответствуют размерам их сенсорного представительства
Читайте еще:
Каждый этап развития человека характеризуется совокупностью специфических закономерностей развития - основными достижениями, сопутствующим образованием, особенностями развития самосознания. А.С. Выготский в качестве критерия возрастной периодизации рассматривал психические новообразования.
К аналогичному заключению пришли Говод и В.Тимм (1999). Ими было установлено, что влияние постгипнотического внушения на точность полиграфных обследований при приеме на работу ничтожно.
Из всех членов Комитета Абрахам и Ференци, по моему мнению, являлись лучшими аналитиками. Абрахам обладал очень четким суждением, хотя ему и не хватало некоторого интуитивного проникновения Ференци. В те дни не существовало и мысли об учебном анализе. Мне кажется, я стал первым психоаналитиком.
-- Погоди. В табличке одни только еоглаеные звуки. Кели соединить их с гласными — ведь гласные сами по себе не означают циФро--то можно составить слова; которые в то же время булут вьражать чиєла.
9. Вы все делаете чрезмерно. Чрезмерно заботитесь о нас, чрезмерно настаиваете на браке, чрезмерно кормите нас.
В книге крупного американского специалиста по психологической войне П. Лайнбарджера приводится неожиданный факт из истории России. В русских летописях и даже в современном русском языке сохранились свидетельства необычайной многочисленности монголо-татарских войск («тьма тьмущая»), хотя.
А те, в комнате которых был и проволочный каркас, и чучело, все свободное время сидели на чучеле — тут же спали, прижимаясь к нему, играли. К проволочной «матери» приходили только тогда, когда хотели есть. Эти шимпанзе развивались вполне нормально.
Мудрецы утверждают: «Дорогу осилит идущий». У каждого, кому выпадет счастливая карта идти вместе с Натальей, шансы на достижение успеха удваиваются. Она и обнадежит, и поддержит, и окрылит.
Как-то Шура, уборщица детсадика нашего, где я сторожевал, руку мою взяла и говорит: «Дай, Иван Афанасич, по ладошке тебе погадаю…» - «Ну, ну, погадай, говорю, на счастье». - «Ой, ой, Иван Афанасич, ну и счастливый же ты человек будешь». - «Почему буду? Уже есть». - «А будешь-то какой счастливый.
С помощью несложного теста попробуйте определить стиль ваших отношений с людьми, его достоинства и недостатки.
И так они расслабляли мышцы другой руки, грудные мышцы, мышцы живота, спины, ног, шеи до тех пор, пока все тело не стало мягким, податливым и расслабленным.
ствии влиятельными учеными. Этому немало способствовала общительная натура Уорнера и его преподавательский дар «Уорнер всегда поощрял студентов к новаторству и работал в тесном контакте с ними. Он был живым учителем, который исчерчивал доски диаграммами и охотно обменивался с аудиторией идеями.
ТАКТИЛЬНЫЙ АНАЛИЗАТОР
Тактильный анализатор (лат. tactilis осязаемый) — совокупность периферических и центральных нервных образований, обеспечивающих восприятие и переработку информации о действии на наружные покровы организма различных неболевых механических раздражителей.
Тактильная чувствительность является составной частью осязательных ощущений (см. Осязание), а Тактильный анализатор — частью соматосенсорного анализатора. Механические воздействия, обусловливающие возникновение тактильных ощущений, обычно подразделяют на прикосновение и давление. Часто в качестве самостоятельного вида тактильных раздражителей выделяют вибрацию, фактически представляющую собой разновидность ритмического прикосновения.
Периферический отдел Тактильного анализатора образуют различные виды механорецепторов (см.) и связанные с ними афферентные нервные волокна (см.). Основной вид механорецепторов в волосистой части кожи (90% всей поверхности кожи) составляют свободные нервные окончания (см.), нервные окончания вокруг волосяных фолликулов, диски (тельца) Меркеля (осязательные мениски), тактильные корпускулы Игго. В коже, лишенной волосяного покрова, основными рецепторными структурами являются свободные нервные окончания, диски Меркеля, тельца Мейсснера (осязательные тельца) и Пачини (пластинчатые тельца). Тактильные рецепторы на поверхности тела распределены весьма неравномерно, что обусловливает неодинаковую тактильную чувствительность ее разных участков. Наиболее тонкая тактильная чувствительность отмечается на кончиках пальцев рук и языка, наименьшая — на коже брюшной и ягодичной областей. Очень богатой афферентной иннервацией и соответственно тактильной чувствительностью обладают вибриссы — специальные чувствительные волоски, встречающиеся у многих приматов и других млекопитающих, но отсутствующие у человека. Активность тактильных рецепторов совместно с рецепторами опорнодвигательного аппарата лежит в основе сложных осязательных ощущений.
Восприятие прикосновения (и вибраций) осуществляется с помощью фазных (быстро адаптирующихся) рецепторов, а давления — благодаря возбуждению тонических (медленно адаптирующихся) рецепторов. К первым относятся тельца Пачини, ряд рецепторных структур волосяной сумки (так наз. Т-, G-, D-рецепторы, являющиеся окончаниями афферентных волокон Аа и Аа групп), ко вторым — тактильные корпускулы, тельца Руффини (окончания волокон Аа группы), свободные нервные окончания (разветвления волокон группы С). Абсолютный порог тактильной чувствительности определяется активностью наиболее чувствительных механорецепторов, к-ры-ми являются тельца Пачини, способные возбуждаться уже при смещениях порядка 10 -4 —10 -5 мм. Вибрационные раздражения воспринимаются в диапазоне от долей до 1 000—1 200 гц, но наибольшая чувствительность к вибрациям наблюдается в диапазоне от 200 до 400 гц (см. Вибрационная чувствительность). Помимо абсолютного порога тактильную чувствительность характеризуют дифференциальный порог и порог пространства. Дифференциальный порог — величина, на к-рую нужно изменить действующий раздражитель, чтобы вызвать минимальное изменение ощущения. Дифференциальный порог характеризует процесс адаптации тактильных рецепторов к действию раздражителя и используется для выявления нарушений тактильной чувствительности. Порог пространства — наименьшее расстояние между двумя точками кожи, при одновременном раздражении к-рых возникает ощущение действия двух раздельных стимулов. Пороги пространства зависят от интенсивности ветвления нервного волокна на периферии и числа рецепторов, передающих возбуждение одному нервному волокну. Пороги пространства значительно различаются на разных участках кожи: они минимальны (1 — 2,5 мм) на кончиках пальцев, губах, языке и максимальны (более 60 мм) на коже бедер, плеч, средней линии спины. Для определения дифференциальных порогов и порогов пространства используют те же методические приемы, что и для исследования осязания (см.) и чувствительности кожи (см. Кожа), применяя только тактильные раздражители.
Анализ механизмов возбуждения тактильных рецепторов, составляющих биофизическую основу тактильной рецепции, осуществлен пока еще в малой степени, что связано с большими техническими сложностями исследования одиночных рецепторов, расположенных в плотных тканях тела. Поэтому почти все сведения о деятельности этих рецепторов получены при изучении потоков импульсов, регистрируемых от одиночных афферентных нервных волокон. Единственным исключением являются крупные пластинчатые тельца Пачини, исследование к-рых позволило выяснить назначение и механизмы деятельности всех образующих их структур: капсулы, не-миелинизированной терминали и мякотного нервного волокна. Капсула рецептора (как и окружающая его ткань) выполняет роль вспомогательной структуры, проводящей механическое раздражение извне к нервной терминали. Она является механическим фильтром, определяющим скорость адаптации рецептора, а также его частотные характеристики (при действии вибраций). Благодаря особому строению внутренней части капсулы тельце Пачини, как и ряд других тактильных рецепторов, обладает чувствительностью к направлению действия раздражителя. Собственно процесс рецепции осуществляется мембраной нервной терминали. При одном направлении стимула происходит растяжение поверхностной мембраны терминали, что приводит к возбуждению рецептора, проявляющемуся возникновением деполяризационного рецепторного потенциала и вслед за этим генерацией импульсных разрядов в первом перехвате Ранвье нервного волокна. При другом направлении стимула мембрана терминали подвергается сжатию, что ведет к появлению тормозных гиперполяризационных рецепторных потенциалов. Рецепторные потенциалы представляют собой местные ответы поверхностной мембраны нервной терминали. Известно, что деполяризационные рецепторные потенциалы вызываются увеличением проницаемости мембраны преимущественно к ионам натрия, к-рые переносят основную (до 90%) часть зарядов.
Афферентные нейроны первого порядка от тактильных рецепторов туловища и конечностей располагаются в спинномозговых узлах, а от рецепторов лица и головы — в полулунных (гассеровых) узлах тройничных нервов (тройничных узлах, Т.). Сведения о восходящих путях тактильной афферентации получены гл. обр. в экспериментах на животных, преимущественно кошках. Основная часть быстропроводящих (Аа) волокон, занимающих медиальную часть задних корешков, после вхождения в спинной мозг направляется в составе задних столбов белого вещества к nuci, gracilis и nuci, cuneatus продолговатого мозга. Расположенные в этих ядрах тела афферентных нейронов второго порядка дают начало волокнам медиальной петли, образующим перекрест на уровне олив продолговатого мозга и заканчивающимся в специфических вентробазальных ядрах таламуса. Информация от тактильных рецепторов головы также достигает этих структур. Здесь локализуются нейроны третьего порядка, к-рые посылают свои волокна в первую соматосенсорную область коры полушарий большого мозга (постцентральную извилину). По быстро-проводящим волокнам от тактильных рецепторов тела в таламус поступает также информация по латеральному тракту Морина. Начавшись от клеток задних рогов спинного мозга, этот спиноталамический путь переключается в боковом шейном ядре продолговатого мозга, а затем после перекреста сливается с медиальной петлей. Медленно проводящие волокна от тактильных рецепторов тела проходят в латеральной части задних корешков и заканчиваются на крупных канатиковых клетках задних рогов спинного мозга. Отсюда начинаются спиноталамические пути (передние и боковые). На спинальном уровне эти пути осуществляют частичный перекрест и дают диффузную проекцию в комплекс вентробазальных ядер таламуса (более вентрально, чем медиальная петля), в неспецифические ядра таламуса и в крупноклеточный отдел медиального коленчатого тела. Отсюда начинаются волокна, идущие во вторую соматосенсорную область коры (у кошки — в переднюю экто-сильвиеву извилину, у человека и обезьян — кзади и вентральнее первой соматосенсорной области). Туда же поступают волокна, связанные с трактом Морина. Благодаря этим волокнам сигналы достигают второй соматосенсорной области коры с коротким латентным периодом. В коре выделяют иногда и третью соматосенсорную зону, к-рая у животных располагается между передней эктосильвиевой и ансатной извилинами. Ответы ее нейронов имеют большие латентные периоды, чем ответы нейронов других двух зон, и вызываются специфическими раздражителями с небольших рецептивных полей.
Система медиальной петли обеспечивает передачу информации о специфических тактильных раздражениях небольшой интенсивности (прикосновении, легком давлении); латеральный тракт Морина передает сведения о более сильных деформациях кожи, а спиноталамическая система является путем, по к-рому поступают сигналы о значительных механических воздействиях, причем этот путь является неспецифическим, обеспечивающим кодирование информации и о температурных воздействиях. У нейронов системы медиальной петли рецептивные поля невелики, а для всей системы характерна строгая топографическая организация проекций областей поверхности кожи. У нейронов же спиноталамической системы рецептивные поля очень большие, покрывающие иногда все тело, но в ее ядрах нет четкого топического представительства поверхности тела. В целом система задние столбы — медиальная петля осуществляет специфические формы тактильного различения, а спиноталамическая система дает значительно более генерализованные формы ощущений и связана с передачей информации о качественной природе влияний с периферии, а не о локализации и форме раздражителя, его изменении во времени.
Совместно с другими видами чувствительности кожи (см. Кожа) тактильная чувствительность может в какой-то степени компенсировать отсутствие или недостаточность функции других органов чувств (зрения, слуха). У слепых количество телец Пачини в коже пальцев и ладоней резко возрастает, и тактильная чувствительность этих областей значительно превышает таковую у зрячих. Нарушения тактильной чувствительности (см. Осязание) тесно связаны с нарушениями других видов чувствительности кожи и в клинике исследуются вместе с ними.
Библиография: Есаков А. И. и Дмитриева Т. М. Нейрофизиологические основы тактильного восприятия, М., 1971, библиогр.; Ильинский О. Б. Физиология механорецепторов, Л., 1975; Физиология сенсорных систем, под ред. А. С. Батуева, Л., 1976.
ВИБРАЦИОННАЯ ЧУВСТВИТЕЛЬНОСТЬ
Вибрационная чувствительность — свойство организма воспринимать действие ритмических колебаний, возникающих в окружающей среде и передаваемых через предметы, с которыми организм находится в контакте, а также через почву, воздух, воду. Источниками вибрации в среде служат естественно существующие в природе явления (колебание водных и воздушных масс, земной коры) и созданные человеком различные моторы, двигатели и т. п. Широкое распространение вибрации (см.) определяет важную роль Вибрационной чувствительности в жизнедеятельности организмов. Ощущения, основанные на Вибрационной чувствительности, зависят от особенностей раздражения (частоты колебаний, амплитуды, продолжительности) и от особенностей области тела, на к-рую раздражение действует с наибольшей силой.
Основные структуры, обеспечивающие Вибрационную чувствительность,— это преимущественно механорецепторы, расположенные в поверхностных и глубоких слоях кожи. Нервные окончания, локализованные в глубоких слоях кожи, возбуждаются при малых амплитудах раздражения (от долей микрона до нескольких микрон). Рецепторы поверхностных слоев кожи возбуждаются лишь при достаточно больших амплитудах воздействия (десятки микрон) и в меньшей степени обусловливают В. ч. В развитии В. ч. могут участвовать и механорецепторы, заложенные в фасциях, надкостнице, тканях суставов; определенной чувствительностью к вибрации обладает и вестибулярный аппарат.
Рецепторы, воспринимающие вибрацию,— это, как правило, фазные механорецепторы типа пластинчатых телец, которые отвечают лишь одним — двумя импульсами на действие каждого вибрационного колебания. Они же определяют и порог восприятия вибрации, являясь наиболее чувствительными механорецепторами кожи. Если кожа не покрыта волосами, диапазон воспринимаемых ею частот колебаний шире, а порог восприятия ниже. Наибольшей В. ч. обладает кожа пальцев рук, где порог возбуждения рецепторов наименьший (амплитуда 0,02 мк при частотах 200— 450 гц). В целом же организмы могут воспринимать вибрацию в достаточно широком диапазоне частот (от долей герц до 1000—1200 гц). При воздействии вибраций импульсы от рецепторов проводятся преимущественно в задних столбах спинного мозга и достигают сомато-сенсорных областей коры головного мозга.
При ряде патологических состояний, обусловленных, напр., поражением спинного мозга или длительным воздействием вибраций, наблюдаются различные расстройства В. ч. (см. Вибрационная болезнь, Вибротравма).
Вибрационная чувствительность составляет физиологическую основу действия вибрационного массажа, применяемого в лечебных целях.
Приборы для определения вибрационной чувствительности
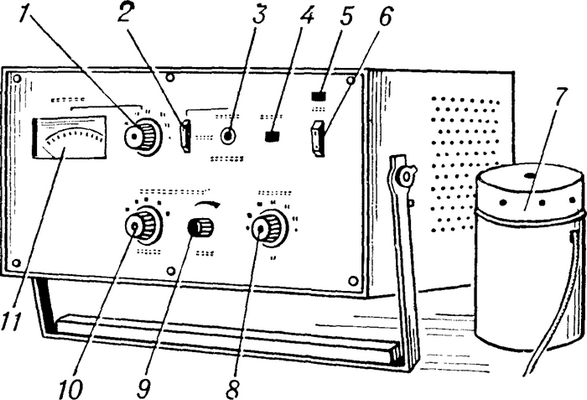
Измеритель вибрационной чувствительности ИВЧ-02: 1 — переключатель диапазонов отсчета; 2 — переключатель подачи прерывания сигнала; 3 — кнопка прерывания сигнала; 4 — индикаторная лампа ответа пациента; 5 — индикатор включения; 6 — переключатель сети; 7 — вибратор; 8 — переключатель частот; 9 и 10 — регуляторы амплитуды вибрации; 11 — измерительный прибор.
Приборы для определения вибрационной чувствительности предназначены для измерения чувствительности к вибрации кожи различных участков тела человека. Принцип действия этих приборов состоит в возбуждении механических колебаний какого-либо участка тела, исследуемого с помощью вибратора, и измерении (отсчете) порогового уровня вибрации при речевом ответе пациента.
Прибор для определения вибрационной чувствительности состоит из вибратора, генератора электрических сигналов и измерительного, или отсчетного, устройства. В качестве вибратора, позволяющего преобразовать электрические колебания в механические, используют пьезоэлектрические, магнитострикционные и электродинамические преобразователи. Генераторы электрических сигналов работают на фиксированных частотах предпочтительно следующего ряда: 16; 32; 63; 125; 250; 500; 1000 гц. Измерительное, или отсчетное, устройство градуируется в определенных единицах. Первые приборы градуировались по величине электрического напряжения, подаваемого с генератора на вибратор; позднее измерялась величина вибросмещения штока вибратора. В современных приборах производят отсчет виброскорости штока вибратора в сантиметрах в 1 сек. или в децибелах относительно физиологического нулевого уровня, являющегося среднестатистическим значением для молодых практически здоровых лиц. Последний тип отсчета наиболее удобен. Величина физиологического нулевого уровня на каждой из рабочих частот задается в децибелах относительно 5-10“6 см/сек. Разность между пороговым и нулевым уровнем характеризует величину потери вибрационной чувствительности для данного испытуемого аналогично определению степени потери слуха при аудиометрии (см.).
В СССР разработаны приборы для определения чувствительности к вибрации ВТ-2 и ИВЧ-02 (см. рисунок и таблицу).
Прибор ИВЧ-02 (стационарный) применяется в клинике и в эксперименте; прибор ВТ-2 (переносный) может быть использован в медсанчастях, поликлиниках.
Глава 3 ПЕРЕДАЧА И ПЕРЕРАБОТКА СЕНСОРНЫХ СИГНАЛОВ
Сенсорные сигналы несут в мозг внешнюю информацию, необходимую для ориентации во внешней среде и для оценки состояния самого организма. Эти сигналы возникают в воспринимающих элементах (рецепторах) и передаются в мозг через цепи нейронов и связывающих их нервных волокон сенсорной системы. Процесс передачи сенсорных сигналов сопровождается их многократными преобразованиями и перекодированием на всех уровнях сенсорной системы и завершается опознанием сенсорного образа. Этим процессам посвящён ряд обзорных работ [Черниговский, 1960; Физиол. сенсорн. систем, 1971, 1972, 1975; Кейдель, 1975; Сомьен, 1975; Тамар, 1976; Батуев, Куликов, 1983; Глезер, 1985; Дудел и др., 1985, Хьюбел, 1990; Физиол. зрения, 1992].
Основные функции сенсорной системы. Каждая сенсорная система выполняет ряд основных функций, или операций с сенсорными сигналами. Эти функции таковы: обнаружение сигналов, их различение, передача, преобразование и кодирование, а также детектирование признаков сенсорного образа и его опознание. Обнаружение и первичное различение сигналов обеспечивается уже рецепторами, а их детектирование и опознание - нейронами корковых уровней сенсорной системы. Передачу, преобразование и кодирование сигналов осуществляют нейроны всех уровней системы.
1. ОБНАРУЖЕНИЕ И РАЗЛИЧЕНИЕ СИГНАЛОВ
1. ОБНАРУЖЕНИЕ И РАЗЛИЧЕНИЕ СИГНАЛОВ Сенсорная рецепция Рецептором называют специализированную клетку, эволюционно приспособленную к восприятию из внешней или внутренней среды определённого раздражителя и к преобразованию его энергии из физической или химической
2. ПЕРЕДАЧА И ПРЕОБРАЗОВАНИЕ СИГНАЛОВ
2. ПЕРЕДАЧА И ПРЕОБРАЗОВАНИЕ СИГНАЛОВ Процессы передачи и преобразования сигналов обеспечивают поступление в высшие сенсорные центры наиболее важной (существенной) информации о сенсорном событии в такой форме, которая удобна для надёжного и быстрого анализа. Что
4. ДЕТЕКТИРОВАНИЕ СИГНАЛОВ
4. ДЕТЕКТИРОВАНИЕ СИГНАЛОВ Детектированием называют избирательное выделение сенсорным нейроном того или иного признака раздражителя, имеющего поведенческое значение. Осуществляют такой анализ нейроны-детекторы, избирательно реагирующие лишь на определённые свойства
7. ВЗАИМОДЕЙСТВИЕ СЕНСОРНЫХ СИСТЕМ
7. ВЗАИМОДЕЙСТВИЕ СЕНСОРНЫХ СИСТЕМ Взаимодействие сенсорных систем осуществляется на спинальном, ретикулярном, таламическом и корковом уровне. Особенно широка интеграция сигналов в ретикулярной формации. В коре мозга происходит интеграция сигналов высшего порядка. В
Глава 4 ПСИХОФИЗИОЛОГИЯ СЕНСОРНЫХ ПРОЦЕССОВ
Глава 4 ПСИХОФИЗИОЛОГИЯ СЕНСОРНЫХ ПРОЦЕССОВ 1. ОБЩИЕ СВОЙСТВА СЕНСОРНЫХ СИСТЕМ Сенсорной системой называют часть нервной системы, воспринимающую внешнюю для мозга информацию, передающую её в мозг и анализирующую её. Сенсорная система состоит из воспринимающих
1. ОБЩИЕ СВОЙСТВА СЕНСОРНЫХ СИСТЕМ
1. ОБЩИЕ СВОЙСТВА СЕНСОРНЫХ СИСТЕМ Сенсорной системой называют часть нервной системы, воспринимающую внешнюю для мозга информацию, передающую её в мозг и анализирующую её. Сенсорная система состоит из воспринимающих элементов - рецепторов, нервных путей, передающих
1.1. Методы исследования сенсорных систем
1.1. Методы исследования сенсорных систем Функции сенсорных систем исследуют в электрофизиологических, нейрохимических и поведенческих опытах на животных, проводят психофизиологический анализ восприятия у здорового и больного человека, а также с помощью ряда
Общие принципы организации сенсорных систем
Общие принципы организации сенсорных систем Все сенсорные системы человека организованы по некоторым общим принципам. Важнейшие из них следующие: многослойность, многоканальность, наличие так называемых «сенсорных воронок», а также дифференциация систем по вертикали
4.4. Основные афферентные пути и проекции вестибулярных сигналов
4.4. Основные афферентные пути и проекции вестибулярных сигналов Два основных пути поступления вестибулярных сигналов в кору мозга обезьян следующие: прямой - через вентральное постлатеральное ядро и непрямой - через вентролатеральное ядро. В коре основные афферентные
5.6. Передача и переработка соматосенсорной информации Чувствительность кожи и ощущение движения связаны с проведением в мозг сигналов от рецепторов по двум основным путям (трактам): лемнисковому и спиноталамическому, значительно различающимся по своим
НЕЙРОФИЗИОЛОГИЯ СЕНСОРНЫХ ПУТЕЙ СПИННОГО МОЗГА
НЕЙРОФИЗИОЛОГИЯ СЕНСОРНЫХ ПУТЕЙ СПИННОГО МОЗГА 1. Дорсальные (задние) столбы спинного мозга представляют собой мощную систему волокон различного происхождения (рис. 6.21). Задние столбы у наземных млекопитающих в эволюции формируются в связи с развитием конечностей, когда
Механизм передачи гормональных сигналов через мембранные рецепторы
Механизм передачи гормональных сигналов через мембранные рецепторы Гормоны (первичные посредники) связываются с рецепторами на поверхности клеточной мембраны и образуют комплекс гормон-рецептор. Этот комплекс трансформирует сигнал первичного посредника путем
Передача сигналов через рецепторы, сопряженные с ионными каналами
Передача сигналов через рецепторы, сопряженные с ионными каналами Рецепторы, сопряженные с ионными каналами, являются интегральными мембранными белками, состоящими из нескольких субъединиц. Они действуют одновременно как ионные каналы и как рецепторы, которые
Передача гормонального сигнала: синтез, секреция, транспорт гормонов, их действие на клетки-мишени и инактивация
Передача гормонального сигнала: синтез, секреция, транспорт гормонов, их действие на клетки-мишени и инактивация В определении понятия «гормон» было указано несколько этапов распространения гормонального сигнала (рис. 2.6). Рис. 2.6. Этапы распространения гормонального
Глава 22. ПЕРЕДАЧА ЭЛЕКТРОНОВ
Глава 22. ПЕРЕДАЧА ЭЛЕКТРОНОВ Львиная доля производимой в процессе обмена веществ энергии выделяется в ходе реакций, в которых участвует атмосферный кислород. Перед тем как мы сможем свободно рассуждать об этих реакциях, надо сначала обговорить основные
Передача тактильных сигналов в нервных волокнах. Восприятие вибрации
Понимание физико-химической природы генерации нервного сигнала, путей передачи информации с одной нервной клетки на другую или на мышечную клетку позволит вплотную подойти к объяснению механизма деятельности нервной системы. Нервные клетки передают информацию с помощью сигналов, представляющие собой электрические токи, генерируемой поверхностной мембраной нейрона. Эти токи возникают благодаря движению зарядов, принадлежащих ионам натрия, калия, кальция и хлора.
От наружной среды внутреннее пространство нейрона отделено клеточной мембраной, которая является плохим изолятором и допускает некоторую утечку ионов в обоих направлениях. Внутри нейронов относительно высока концентрация ионов К + , но мало ионов Na + и CI - , снаружи больше натрия и хлора и мало калия. В состоянии покоя мембрана более проницаема для ионов К + чем для ионов Na + . Поэтому ионы калия стремятся выйти из клетки по градиенту концентрации, заставляющий выходить ионы К + наружу. Если бы мембрана была проницаема только для ионов калия, разность потенциалов на ней могла бы достигать величин, определяемой уравнением Нернста (1) для калиевого электрода.
где VK - равновесный потенциал для ионов К + , КН и КВ - активность калия снаружи и внутри волокна, R - газовая постоянная, Т - абсолютная температура и F - постоянная Фарадея. По данным различных авторов, эта величина соответствует 70-75 мВ. Для объяснения реверсии мембранного потенциала (МП) во время развития импульса предполагается, что на гребне спайка мембрана избирательно проницаема для ионов Nа + . Разность потенциалов при этом выражается формулой Нернста для натриевого электрода:
RT [Na] Н
Снижение потенциала покоя (ПП) до определенной величины ведет к проницаемости мембраны к ионам натрия, которые входя в клетку вызывают дальнейшее снижение ПП. Повышенная проницаемость мембраны к ионам Nа + сменяется повышением ее проницаемости к ионам К + . При этом последние выходят из клетки и в результате чего происходит восстановление ПП клетки. Эти изменения разности потенциалов и создают электрический импульс, распространяющийся по нервному волокну. Эксперимент с двумя электродами, введенными в одиночное волокно аксона кальмара, позволил вплотную подойти к вопросу о природе энергии, необходимой для изменения знака потенциала на мембране. Один электрод служит для пропускания тока, другой - для измерения разности потенциалов на мембране. Показано, что если ток течет через мембрану внутрь волокна, то разность потенциалов увеличивается, и возбуждения нет. Ток, направленный наружу, также не вызывает возбуждения. Однако, генератор срабатывает каждый раз, когда напряжение на мембране уменьшается ниже определенной величины, которую принято называть порогом возбуждения. Нервный импульс возникает только в том случае, если вызванное возбуждение любым способом изменяет напряжение мембраны за пороговую величину, которая обычно равна 10-15 мВ. Суммируя вышесказанное можно предположить, что передача электрических сигналов в нервных сетях основан на изменении МП в результате прохождения относительно небольшого числа ионов через мембранные каналы. В результате открывания и закрывания натриевых каналов нервный импульс распространяется вдоль нервного волокна, пока не достигнет его окончания - места контакта с мышечной клеткой или, как принято называть, «концевой пластинкой». В концевой пластинке под действием нервного импульса открываются потенциал-зависимые кальциевые каналы, и ионы Са 2+ входят в нервное окончание, в результате чего нервная клетка освобождает медиатор - ацетилхолин (АХ).
Применение микроэлектродной техники отведения спонтанных биопотенциалов концевой пластинки позволило определить пороговую чувствительность синаптической области мышечной мембраны путем нанесения незначительного количества АХ. Показано, что АХ в количестве 10 8 -10 9 молекул уже вызывает деполяризацию мышечной мембраны в области наружной поверхности синапса. Сама же мембрана является непроницаемой для АХ. При введении АХ внутрь мышечных волокон в районе концевой пластинки, никаких электрических изменений не наблюдалось.
Многочисленными работами показана необходимость притока ионов Са 2+ в окончание аксона для синаптической передачи. Показано, что если во внеклеточной среде кальций отсутствует, АХ не освобождается и передача сигнала не происходит и, во-вторых, если искусственно ввести Са 2+ в наружную среду, омывающий нервно-мышечный препарат при помощи микроаппликации, выход нейромедиатора происходит спонтанно.
t
Рt = е --- (3)
tх
По нашим данным амплитуда МПКП имеет величину порядка 1 мВ и заметно колеблется от 0,1 до 4 мВ. Этот разброс связан, прежде всего с тем, что места возникновения МПКП находятся на разном расстоянии от регистрирующего электрода. МПКП регистрируются внеклеточным микроэлектродом от наружной поверхности мышечных мембран, от различных, но строго локальных участков синапса, что свидетельствует о выделении АХ не диффузно, а в определенных активных точках. При изучении возникновения постсинаптического потенциала концевой пластинки (ПКП) многие исследователи пришли к выводу, что ПКП возникает вследствие резкого увеличения частоты МПКП и, что между частотой и силой поляризующего тока имеется линейная зависимость. Деполяризация пресинаптических окончаний на 60 мВ увеличивает частоту в 10 4 раз, что вызывает появление ПКП. В нормальных условиях такой ПКП состоит из более чем из сотни наложенных друг на друга МПКП. Если предположить, что ПКП состоит из спонтанно возникающих МПКП, то их число в одиночном ПКП должно испытывать отклонения от среднего значения, которые описываются формулой Пуассона. Допустив, что среднее число МПКП в ПКП равно m, тогда вероятность Рх наблюдать ПКП, содержащей х МПКП, будет
m x
Рх = ------ e - m , (4)
X!
где х - порция и m - среднее число порций, освобождаемых при одном импульсе. В виду того, что вероятность отклонений х от m для больших значений мала, возникла необходимость снизить квантовый состав ПКП за счет снижения концентрации Са 2+ и повышения концентрации Mg 2+ . Однако, в последние годы появилось много убедительных данных, в которых показано, что временное распределение интервалов не подчиняется закону Пуассона. Обнаружено существование низко- и высокоамплитудных МПКП, которые возникали в той же самой концевой пластинке. Анализ встречаемости обоих видов МПКП в односекундные и 100миллисекундные непрерывающиеся интервалы показал, что имеются существенные отклонения от пуассоновского распределения, тем большие, чем меньше диаметр волокна и частота МПКП. Этот статистический подход представляет интерес, поскольку позволяет подтвердить предположение о квантовом характере освобождения медиатора.
Читайте также: