Прорастание спор бактерий. Активация споры. Покоящиеся ( некультивируемые ) формы бактерий.
Добавил пользователь Alex Обновлено: 21.01.2026
Процесс спорообразования контролируется более чем 40 оперонами, которые представляют собой как бы дополнительный геном у спорообразующих бактерий. В составе этого генома насчитывается более 60 генов. Инициация споруляции связана с геном spoO, мутации в котором делают невозможным образование споры с самых начальных стадий. Транскрипция гена spoO запускает последовательную транскрипцию всех оперонов спорового генома. При этом их транскрипция носит строго регулируемый характер: выражение более поздних генов зависит от транскрипции более ранних генов. Это обусловливает четкую временнwую последовательность биохимических и морфологических процессов, лежащих в основе споруляции. Спорообразующие бактерии обладают механизмами, с помощью которых они распознают определенные изменения в окружающей среде, например, уменьшение содержания источников энергии, некоторых аминокислот и оснований. В ответ на это в клетке происходят метаболические изменения, которые и запускают споруляцию. Эти изменения приводят прежде всего к изменению субъединичного состава РНКполимераз. Индукция транскрипции спорового генома приводит к синтезу особых ?-единиц РНК-полимераз, которые и обеспечивают распознавание промоторов генов, контролирующих споруляцию. Вместе с тем наличие множественных промоторов у жизненно важных для клетки генов, распознаваемых разными ?-факторами, обеспечивает их выражение на всех этапах роста этих клеток, споруляции и прорастания спор. Одна из особенностей споруляции состоит в том, что на определенном ее этапе (приблизительно на 3-м часу) происходит синтез небольших кислоторастворимых белков. На их долю приходится около 10 - 12 % всех белков споры. В спорах они связываются с ДНК, обеспечивая устойчивость их к УФ-облучению. В момент прорастания споры эти белки гидролизуются и тем самым обеспечивают прорастающую спору необходимыми аминокислотами. У некоторых представителей рода Clostridium выявлена функциональная зависимость токсинообразования от споруляции: наиболее интенсивно экзотоксин вырабатывается во время активной споруляции; причинная связь этих процессов не ясна.
Прорастание споры происходит после получения соответствующего химического сигнала. Различные виды спорообразующих бактерий располагают рецепторами, распознающими наличие в среде источников энергии, L-аланина, аденозина и других веществ. Связывание с такими эффекторами активирует содержащийся в споре автолизин (лизоцим), который быстро разрушает пептидогликан кортекса.
Прорастание спор включает три стадии: активацию, начальную стадию и стадию роста.
Активация является обязательным условием прорастания спор. Она осуществляется различными воздействиями - кислой рН; веществами, содержащими свободные сульфгидрильные группы; повышением температуры; механическим повреждением спор.
Начальная стадия. Под влиянием внешних эффекторов происходит активация автолизина, последний разрушает пептидогликан кортекса, в спору поступает вода, спора высвобождается от дипиколината кальция, под воздействием гидролитических ферментов разрушаются другие ее компоненты.
Стадия роста. После разрушения кортекса и наружных слоев споры из нее появляется («выклевывается») растущая новая вегетативная клетка. Она состоит из протопласта споры и ее клеточной стенки. В ней активизируются биосинтетические процессы; в результате новая вегетативная клетка, при наличии необходимых питательных веществ, удваивает свою биомассу и делится на две дочерние клетки, которые далее активно размножаются, пока этому способствуют условия среды. Процесс прорастания споры контролируется генами как спорового, так и вегетативного геномов.
2. Другие покоящиеся формы бактерий
К другим покоящимся формам бактерий относятся: цисты, экзоспоры, миксоспоры. Как и эндоспоры, все эти покоящиеся формы предназначены для выживания бактерий при неблагоприятных условиях.
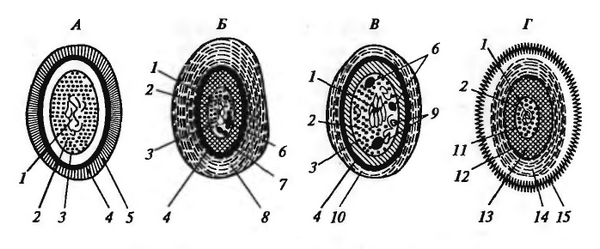
Рис. 5 - Строение покоящихся форм прокариот: А - миксоспоры миксобактерий; Б - цисты азотобактера; В - акинеты цианобактерий; Г - эндоспоры; 1 - нуклеоид; 2 - цитоплазма; 3 - ЦПМ; 4 - клеточная стенка; 5 капсула; 6 - гранулы запасных веществ; 7 - внутренние покровы; 8 - внешние покровы;, 9 - тилакоиды; 10 - чехол; 11 - внутренняя мембрана споры; 12 - наружная мембрана споры; 13 - кортекс; 14 - многослойные покровы споры; 15 - экзоспориум
Покоящиеся клетки прокариот
При переходе к неблагоприятным условиям существования многие прокариоты образуют особые дифференцированные формы - покоящиеся клетки, основным назначением которых является обеспечение переживания вида в неблагоприятных условиях. Их общей морфологической особенностью являются утолщенные клеточные стенки, а также частое образование дополнительных поверхностных структур на фоне одновременной дегидратации и компактизации метаболического и генетического аппаратов. В результате этого покоящиеся клетки характеризуются чрезвычайно низким (фоновым) уровнем обмена веществ и энергии, но приобретают повышенную устойчивость к действию разнообразных повреждающих факторов: высоких и низких температур, кислотности среды, радиации и пр. При попадании же в благоприятные условия покоящиеся клетки прорастают с образованием активно метаболизирующих вегетативных клеток.
Покоящиеся формы прокариот можно условно разделить на несколько видов: эндоспоры, экзоспоры, цисты, акинеты. Эндоспоры - это особый тип покоящихся клеток эубактерий, формирующихся внутри цитоплазмы родительской клетки и обладающих специфическими структурами: внутренней и наружной мембранами, кортексом и многослойными белковыми покровами. К спорообразующим относится большое число эубактерий, относящихся к 15 родам, и имеющих клеточную стенку типичную для грамположительных бактерий. У аэробных микроорганизмов процесс спорообразования лучше всего изучен у р. Bacillus, и у анаэробов р. Clostridium. Переход к процессу спорообразования является дефицит питательных веществ, накопление в среде продуктов метаболизма, действующих по принципу кворум-зависимых регуляторных факторов. В упрощенном виде у бациллюс субтилис споруляция регулируется системой из сигнального олигопептида, фосфатазы и белковых факторов споруляции. При достижении высокой плотности популяции, олигопептид в большом количестве поступает в клетку, инактивирует фосфатазу,активирует таким образом белковые факторы споруляции и запускает программу споруляции.
Последующий процесс споруляции можно рассматривать как неравновеликое бинарное деление особого типа, при котором одна из образующихся клеток формирует отмирающий спорангий, а вторая превращается в покоящуюся эндоспору. Спорообразованию всегда предшествует репликация ДНК, часть которого компактизируется и в составе т.н. «ядерного тяжа» переходит в ту часть клетки, где в последующем и будет формироваться спора. Компактизированный материал теряет свою метаболическую активность на ранних стадиях спорообразования, а сам процесс формирования споры контролируется геномом спорангия.
Помимо разделения генетического материала, в клетке активно идут процессы дегидратации (в споре остается 15% воды), ДНК переходит из В-формы в А-форму, что существенно повышает устойчивость к УФ лучам, белки распадаются до специфичного для спор и отсутствующего у вегетативных клеток вещества - дипиколиновой кислоты (пиридин-2,6-дикарбоновой) кислоты. В ходе ее образования происходит также интенсивное поглощение клеткой ионов кальция, образующего с кислотой хелатное соединение, составляющее 10-15% сухого вещества зрелых спор. Визуально же это проявляется как блестящие тела на фоне темных клеток.
Далее начинается процесс обособления протопласта: ЦПМ впячивается вовнутрь как при делении, но потом протопласт обрастает ЦПМ родительской клетки вторым слоем. Между внутренней и внешней мембраной начинает формироваться кортекс - многослойный пептидогликан, отличающийся высокой степенью поперечной сшивки. Вслед за этим поверх наружной мембраны синтезируются дополнительные многослойные «споровые покровы», состоящие из белков, липидов и гликолипидов. Белки эндоспор богаты цистеином и гидрофобными аминокислотами, что обеспечивает их дополнительную устойчивость, в частности радиорезистентность. Далее у некоторых бактерий формируется еще одна структура - «экзоспориум», окружающий спору в виде свободного чехла и имеющий характерное для каждого вида тонкое многослойное строение.
После завершения процесса спорообразования происходит лизис спорангия, после чего спора высвобождается в окружающую среду. Споры могут сохраняться в течении сотен лет в окружающей среде. При наступлении благоприятных условий происходит прорастание споры, проходя три стадии: активацию, инициацию и вырастание. Экзоспоры фототрофных бактерий формируются в результате отпочковывания от одного из полюсов материнской клетки. Они не имеют каких-либо особенных поверхностных структур, однако клеточная стенка значительно плотнее и толще аналогичной структуры вегетативной клетки. Экзоспоры миксобактерий - миксоспоры - формируются в т.н. «плодовом теле» псевдоплазмодия, клетки переходят в покоящееся состояние, их клеточная стенка утолщается и окружается капсулой. Цисты распространенный тип покоящихся клеток, образуемые представителями разных групп эубактерий, виды Azotobacter и скользящие бактерии.При истощении пищевых ресурсов в цисту превращается вся палочковидная клетка, а не только какая-то ее часть, что сопровождается изменением ее морфологии. Происходящие изменения включают потерю жгутиков, накопление в цитоплазме гранул поли-бета- гидроксимаслянной кислоты и образованием дополнительных клеточных покровов. Между ЦПМ и КС - формируется «интина», а поверх КС - «экзина». Возникающие толстостенные шарообразные покоящиеся клетки - цисты - устойчивы к высушиванию, механическим нагрузкам и облучению, но не к дейтсвию высоких температур. Акинеты - это особая форма сохранения у планктонных нитчатых цианобактерий, обладающая повышенной устойчивостью к ряду пониженным температурам, высушиванию, но не к повышенным температурам. В акинету превращается вся вегетативная клетка, которая начинает этот процесс с увеличения своих размеров и накопленияв цитоплазме гранул запасных веществ (гликогеновых, полифосфатных, цианофициновых), а также карбоксисом при одновременном уменьшении количества хлорофилла и фикобилиновых пигментов. Пептидогликановый слой КС утолщается, окружающий слизистый чехол уплотняется за счет полисахаридных фибрилл. Под действием силы тяжести акиеты опускаются на дно водоемов и перезимовывают, а весной прорастают, после чего молодые, снабженные газовыми везикулами вегетативные клетки всплывают к поверхности воды.
Заверщая рассмотрение данного вопроса, следует также указать, что многие бактерии способны некоторое время сохранятся в неблагоприятных условиях существования и без каких-либо явных признаков структурной дифференцировки. Однако в этом случае в них происходят существенные физиологические перестройки, заключающиесяв резком угнетении активности метаболических процессов, снижении содержании АТФ и абсолютного значения мембранного потенциала. В результате микроорганизмы впадают в так называемое «некультивируемое» состояние, аналогичное переходу в анабиоз.
Покоящиеся формы бактерий. Споры и самообразования, прорастание спор. Свойства спор.
Специализированные анабиотические формы (такие как эндо- и экзоспоры, миксоспоры, цисты) известны лишь для ограниченного круга микроорганизмов. Однако как у спорообразующих микроорганизмов (Bacillus cereus) на фоне подавления спорообразования, так и у споронеобразующих (Pseudomonas carboxidoflava, Micrococcus luteus, Escherichia coli) бактерий внеклеточные микробные метаболиты, проявляющие свойства индукторов анабиоза, вызывают образование цистоподобных рефрактерных клеток (термин связан с переходом целой клетки в покоящуюся форму и увеличенной способностью данных клеток по сравнению с обычными сильно преломлять свет). В этих клетках снижение метаболической активности (гипометаболизм) сопровождается развитием резистентности к экстремальным воздействиям. Эти формы отличаются нерегистрируемым уровнем эндогенного дыхания, повышенной терморезистентностью и специфической ультраструктурой. По сравнению с вегетативными клетками, цистоподобные рефрактерные клетки обладают рядом особенностей ультраструктурной организации: у них снижена плотность рибосом в цитоплазме, цитоплазма приобретает мелкозернистую структуру, увеличена толщина клеточной стенки, в самой клеточной стенке появляются слоистость и некоторые плотные включения. Для споронеобразующих микроорганизмов цистоподобные рефрактерные клетки — единственная покоящаяся форма, способная сохраняться в течение нескольких лет, а для спорообразующих бактерий — альтернатива спорообразования. Поскольку рефрактерные клетки невозможно выявить с помощью традиционных бактериологических подходов, то в самое ближайшее время они могут создать серьёзную проблему при оценке их эпидемиологической и экологической значимости.
Прокариотические организмы бактерии обладают способностью к спорообразованию, которая заключается в том, что при наступлении условий, неблагоприятных для жизни, клетка частично теряет воду, объём и форму; под внешней мембраной образуется плотная сферическая оболочка.
В виде споры бактерия может выдерживать огромные механические, температурные и химические нагрузки. Например, некоторые споры выдерживают трёхчасовое кипячение или температуру жидкого азота. Также в виде споры более эффективно проходит расселение, потому что частично обезвоженная клетка имеет меньшую массу. Споры неустойчивы к ультрафиолету, как и вообще бактерии, и быстро погибают под таким излучением. Поэтому меньше всего бактерий — в высокогорной местности, а некоторые методы лечения инфекционных заболеваний, вызванных бактериальным возбудителем, предусматривают облучение пациента. Следует помнить, что споры у бактерий, в отличие от растений и грибов, служат не для размножения.
Споры бактерий окрашивают по методу Пешкова или по методу Марцелли.
Спорообразование у бактерий наблюдается в неблагоприятных условиях существования: при недостатке питания, высыхании среды, накоплении в ней вредных продуктов обмена, недостатке воздуха, резких колебаниях температуры и др. Сущность спорообразования заключается в уменьшении интенсивности процессов обмена у микробов и снижении их активности.
В процессе спорообразования цитоплазма теряет влагу, сгущается, собирается в определенном участие тела бактерии, облачается плотной непроницаемой для растворов оболочкой, в результате чего содержимое клетки принимает вид округлого или овального образования, которое называется спорой. В зависимости от размеров и расположения споры внешний вид микробной клетки либо не изменяется, либо принимает другую форму. Если спора образуется в центре клетки, то последняя принимает форму бочонка, лимона, веретена.
У столбнячной палочки спора образуется на конце, и поэтому клетка приобретает форму барабанной палочки. Споровые бактерии отличаются от вегетативных форм большой устойчивостью, легко переносят высушива-ние, замораживание, кратковременное или длительное кипячение, воздействие различных химических веществ. К спорообразующим бактериям относятся: возбудители сибирской язвы, столбняка, ботулизма, а также некоторые сапрофитные обитатели почвы. Спорообразующие патогенные бактерии часто встречаются в унавоженной почве. Они могут попадать в рану с землей.
Прорастание споры в оптимальных условиях осуществляется в течение 2-3 ч; процент проросших спор увеличивается после соответствующей предварительной обработки. Например, споры могут быть активированы кратковременным прогреванием.
• Прорастанию предшествует активация споры. Её инициируют различные химические вещества, повышение температуры и влажности. Под воздействием автолизинов происходит расщепление кортекса, поглощение воды и набухание. Внешне процесс проявляется увеличением («вздутием») споры и уменьшением коэффициента светопреломления. При этом в споре происходят глубокие физиологические изменения: усиливается дыхание, увеличивается активность ферментов, происходит выделение аминокислот, дипиколиновой кислоты и пептидов (потеря сухой массы споры может достигать 20-30%). В тот период спора утрачивает терморезистентность. Затем спора лопается в произвольном месте и из неё выходит вегетативная клетка, снабжённая у подвижных видов жгутиковым аппаратом.
Выявление спор бактерий
Окрашивание спор удается только в мазках.
1. Мазки окрашивают в растворе карболового фуксина или фуксина на анилиновой воде в термостате при 37 °С в закрытой посуде в течение нескольких часов.
Если вам нужна помощь в написании работы, то рекомендуем обратиться к профессионалам. Более 70 000 авторов готовы помочь вам прямо сейчас. Бесплатные корректировки и доработки. Узнайте стоимость своей работы.
2. Обесцвечивают 2,5 % раствором серной кислоты в течение 3 — 5 с и споласкивают 96 % спиртом, пока не перестанут отходить облачка красителя.
3. Докрашивают метиленовым синим 3 — 5 мин.
4. Ополаскивают в проточной воде, сушат на воздухе и изучают при масляной иммерсии.
Можно окрашивать карболовым фуксином, подогревая на огне мазок с красителем до появления паров 5 — 8 мин, но тогда дифференцировать следует 2,5 % серной кислотой в течение 10-20 сек.
Результат: споры окрашиваются в красный цвет, бактерии - в синий.
1. Фиксированные мазки помещают на 2 мин. в хлороформ.
2. Переносят в 5 % водный раствор хромовой кислоты на 2 — 10 мин.
3. Промывают в проточной воде и окрашивают карболовым фуксином 1 мин (стекло при этом подогревают на горелке до появления паров).
4. Дифференцируют в 5 % серной кислоте 5 с.
5. Хорошо промывают в проточной воде и подкрашивают водным раствором метиленового синего 3 мин.
6. Промывают в проточной воде, сушат и изучают при иммерсии.
Результат: опоры окрашиваются в красный цвет, бактерии — в синий.
Окраска малахитовым зеленым Шеффера-Фултона
1. Мазки, фиксированные над огнем, окрашивают 5 % водным раствором малахитового зеленого 5—10 мин.
2. Промывают в проточной воде.
3. Докрашивают 1 % водным раствором сафранина 30 с.
4. Промывают водой, сушат и изучают при масляной иммерсии.
Результат: споры окрашиваются в зеленый цвет, бактерии — в красный.
На нефиксированный мазок наливают 0,5 % раствор соляной кислоты и подогревают над огнем 1 — 2 мин. С препарата сливают кислоту, тщательно промывают проточной водой, высушивают и после высыхания фиксируют на пламени. Фиксированный препарат окрашивают по методу Циля — Нильсена.
Результат: тела бактерий окрашиваются в синий цвет, споры — в красный.
Нефиксированный мазок обрабатывают 10 % формалином 10 мин, промывают проточной водой и высушивают. Окрашивают 3 мин аммиачным метиленовым синим (20 мл насыщенного спиртового раствора метиленового синего + 3 мл 98 % раствора аммиака + 80 мл дистиллированной воды), нагревая стекло над огнем до закипания красителя. Промывают проточной водой, докрашивают 0,5 % раствором сафранина 3—5 мин, промывают водой, высушивают и изучают при масляной иммерсии.
Результат: бактерии красные, споры синие.
Споры бактерий - это приспособление к выживанию в неблагоприятных условиях.
Плодовые тела несут споры, которыми грибы размножаются.
Споры бактериальные овальные или округлые образования, возникающие внутри палочковидных клеток — спороносных бактерий. Внутри каждой С. б. имеются компактное скопление дезоксирибонуклеиновой кислоты (См. Дезоксирибонуклеиновая кислота) (ДНК), Рибонуклеиновые кислоты (РНК) и белок. С. б. устойчивы к действию ядовитых веществ и др. отрицательных внешних факторов, что объясняется меньшим содержанием в них воды (15—20%), чем в вегетативных клетках, наличием 4—5 плотных труднопроницаемых оболочек и переходом ферментов в неактивное состояние. Устойчивость спор к высокой температуре (некоторые споры выдерживают кипячение в течение 30 и более мин) определяется присутствием в оболочках значительного количества кальциевой соли дипиколиновой кислоты. Попадая в свежую питательную среду, споры прорастают (полярно или экваториально), давая начало новым бактериальным клеткам.
Поможем написать любую работу на аналогичную тему
Покоящиеся формы бактерий. Споры и самообразования, прорастание спор. Свойства спор.
Споры, капсулы у бактерий, некультивируемые формы бактерий. Методы изучения
Капсула бактерий - это утолщенный наружный слой клеточной стенки. Капсулы могут быть построены из полисахаридов (пневмококк) или белков (возбудитель сибирской язвы). Большинство бактерий, особенно патогенных, образуют капсулу только в организме человека или животных. Однако, существует род истинно капсульных бактерий (Klebsiella), представители которого образуют капсулу и при культивировании на искусственно питательных средах. Некоторые бактерии могут иметь микрокапсулу, например, эшерихии, или неявно выраженную способность к капсулообразованию - так называемую «нежную капсулу», например, золотистые стафилококки, менингококки. Основное предназначение капсулы - защита бактерий от фагоцитоза.
Споры бактерий представляют собой бактериальные клетки в состоянии анабиоза и образуются при неблагоприятных условиях внешней среды. Располагаться могут внутри клетки терминально, субтерминально или центрально. В процессе спорообразования клетка почти полностью теряет воду, смарщивается, клеточная стенка уплотняется, появляется новое вещество дипиколинат кальция, которое образует комплексы с биополимерами клетки, устойчивые к действию температуры и ультрофиолетовых лучей. В окружающей среде споры бактерий могут сохраняться годами, но при попадании в благоприятные условия спора впитывает влагу, комплексы распадаются, дипиколинат разрушается и спора превращается в вегетативную клетку.. При этом надо запомнить - 1 клетка- 1спора- 1 клетка. Спорообразование характерно в основном для Гр(+) бактерий.
У Гр(-) эквивалентом спорообразования является переход в так называемое некультивируемое состояние. В такой форме они могут длительно сохраняться в окружающей среде.
Выявление капсулы по методу Гинса:
1. На предметное стекло наносят каплю туши, а рядом - каплю исследуемого материала. Обе капли тщательно перемешивают и с помощью шлифованного стекла готовят мазок.
2. Мазок высушивают на воздухе и фиксируют на пламени горелки.
3. Мазок окрашивают фуксином в разведении 1:3 или сафранином. При этом бактерии окрашиваются в красный цвет, капсулы остаются неокрашенными и выделяются на темном фоне препарата
Окраска спор по методу Ожешки:
1. На нефиксированный мазок наносят 0,5% раствор хлористоводородной кислоты и подогревают на пламени горелки в течение 2-3 минут.
2. Кислоту сливают, препарат промывают водой, просушивают и фиксируют над пламенем горелки.
3. Окрашивают препарат по Цилю-Нильсену. Споры бактерий при этом приобретают красный цвет, а вегетативные формы - синий.
5. Особенности метаболизма у бактерий, методы его изучения.
Особенности метаболизма у бактерий.
· Метаболизм бактерий характеризуется ярко выраженным разнообразием.
· Напряженность (интенсивность) процессов метаболизма.
· Все процессы направлены на рост и размножение клетки.
· Преобладают процессы распада (катаболизма).
· В метаболизме у бактерий принимают участие как эндоферменты, так и экзоферменты.
Различают конститутивные и индуцибельные ферменты. Конститутивные ферменты синтезируются клеткой непрерывно, вне зависимости от наличия субстратов в
питательной среде. Индуцибельные (адаптивные) ферменты синтезируются только при наличии в среде субстрата данного фермента. Например, кишечная палочка на среде с глюкозой практически не образует Р-галактозидазу, но резко увеличивает ее синтез при выращивании на среде с лактозой или другим Р-галактозидом.
Некоторые ферменты (так называемые ферменты агрессии) разрушают ткань и клетки, обусловливая широкое распространение в инфицированной ткани микроорганизмов и их токсинов. К таким ферментам относят гиалуронидазу, коллагеназу, дезоксирибонуклеазу, нейраминидазу, лецитовителлазу и др.
Изучается углеводный обмен в процессе культивирования исследуемых бактерий на средах Гисса (полужидких средах, содержащих углевод и индикатор, реагирующий на появление кислых продуктов распада углеводов).
Пептолитические свойства - способность расщеплять только промежуточные продукты распада белков - пептоны, до элементарных радикалов) изучается при посеве бактерий в мясо - пептонный бульон. Под крышку пробирки помещают индикаторные полоски, способные изменять свой цвет в присутствии летучих продуктов распада пептонов(аммиака, сероводорода, индола).
Протео - и пептолитические свойства изучают для идентификации возбудителей.

Споры, капсулы бактерий, некультивируемые формы бактерий. Методы изучения.
Капсула - это структура, чаще всего полисахаридной природы, расположенная кнаружи от клеточной стенки. Её толщина в среднем более 0,2 мкм. Капсула может состоять и из белка (бациллы сибирской язвы).
Это очень гидрофильная структура, поглощающая много воды, и её роль здесь защитная, т.е. определяет устойчивость бактериальных клетках при высушивании.
С другой стороны, капсула защищает микроорганизм от фагоцитоза, а также имеет антигены.
Микрокапсула - это слизистое образование толщиной менее 0,2 мкм, выявляемое только при электронной микроскопии.
От капсулы и микрокапсулы следует отличать слизь - отдельные экзополисахариды, выделяемые бактериальной клеткой во внешнюю среду, и которые с клеткой не связаны. Большое количество слизи выделяют некоторые штаммы синегнойной палочки, выделенных у больных муковисцидозом.
Капсулы выявляют по методу Бурри-Гинса. Суть этого метода заключается в последовательности действий:
- на предметное стекло капают каплю туши, в неё добавляют чистую культуру;
- другими стеклом размазывают каплю по поверхности (равномерно, как при приготовлении мазка крови);
- далее к мазку туши добавляют фуксин
При микроскопировании общий фон чёрный, клетки бактерий розовые, а капсулы остались неокрашенными.
В большинстве случаев капсулы образуются только в макроорганизме, но есть бактерии рода Klebsiella, которые иначе называют истинно капсульными бактериями, потому что они формируют капсулы и в колониях на питательных средах.
Споры - это особые структуры, образуемые бактериальной клеткой при попадании её в неблагоприятные условия.
Споры образуют палочки рода Bacillus и также рода Clostridium. У первых диаметр споры не превышает размер клетки, а у вторых он несколько больше.
Споры являются кислотоустойчивыми, наиболее часто их идентифицируют по методу Ожешко:
- к мазку добавляют 0,5-% HCl;
- окрашивают по методу Циля-Нильсена: через фильтровальную бумагу добавляют фуксин Циля, затем подогреваем до отхождения паров. Далее промываем и погружаем в 5-% раствор H2SO4, затем вновь промываем и к мазку добавляем метиленовый синий.
При микроскопии споры окрашиваются в красный цвет, клетки бактерий в голубой.
Процесс спорообразования начинается с отторжением участка хромосомы и цитоплазмы с инвагинацией цитоплазмы - образуется проспора. Проспору окружают 2 мембраны, слой, прилежащий к внутренней мембране, называется стенкой споры (состоит из пептидогликана).
Между наружной мембраной и стенкой споры расположен слой пептидогликана с большим количеством сшивок - кортекс.
Кнаружи от внешней мембраны расположена оболочка споры из кератиноподобных белков, а в некоторых случаях кнаружи от оболочки споры расположен слой липопротеина - экзоспориум.
Таким образом, спора - многослойная структура, при образовании которой поглощается много дипиколиновой кислоты и кальция, и кислотоустойчивость споре по большей части придаёт именно дипиколинат кальция.
Некультивируемые формы бактерий - бактериальные клетки, имеющие низкую метаболическую активность, не способные размножаться, не культивируются на питательных средах.
Такие формы образуются у некоторых неспорообразующих бактерий при попадании в неблагоприятные условия существования. Основной метод их выявления - ПЦР.
Особенности метаболизма у бактерий. Конститутивные и индуцибельные ферменты бактерий.
Метаболизм бактерий направлен прежде всего на обеспечение клетки нужными питательными веществами, необходимых для их роста и размножения.
По тому, какой углерод используют для своего метаболизма бактерии, их делят на 2 группы:
- автотрофы (используют неорганический углерод CO2);
- гетеротрофы (используют в качестве источника углерода органические соединения: лактат, жирные кислоты, многоатомные спирты, сахара, аминокислоты).
По источнику получаемой энергии все микроорганизмы делятся также на 2 группы: фототрофы (используют энергию солнечного света) и хемотрофы (используют энергию ОВР).
В свою очередь, хемотрофы делятся на литотрофов (в качестве донора электронов выступают неорганические молекулы типа H2S, NH3, Fe 3+ ) и органотрофов (используют в качестве донора электронов органические молекулы). Медицинская микробиология изучает гетерохемоорганотрофов.
Степень гетеротрофности различных микроорганизмов неодинакова. Различают сапрофиты - бактерии, питающиеся мёртвыми органическими остатками и паразиты - бактерии, осуществляющие свой рост и размножения за счёт клеток хозяина.
Паразитизм - фундаментальное биологическое явление, но оказалось, что и группа паразитов является разнородной. Так, выделяют облигатные паразиты и факультативные паразиты. В первом случае бактериальные клетки существуют только в клетках хозяина. Это в том числе представители рода Rickettsia и Chlamydia.
Ферменты бактерий.
В микробной клетке осуществляется большое количество метаболических превращений, которые осуществляются при участии большого количества ферментов. Все ферменты бактериальной клетки принадлежат к 6 основным классам: оксидоредуктазы, гидролазы, трансферазы, изомеразы, лигазы и лиазы.
Ферменты по их локализации в клетке могут быть экзоферментами и эндоферментами.
Экзоферменты выделяются в окружающую среду, большинство этих ферментов - гидролазы. С одной стороны, эти ферменты играют важную роль в питании микробной клетке, поскольку они расщепляют сложные биополимеры до простых фрагментов - сахара, аминокислоты, которые легко попадают в микробную клетку. К экзоферментам относят так называемые ферменты агрессии и защиты. Это те ферменты, которые обеспечивают проникновение бактерий в клетки хозяина (IgA-протеаза, нейраминидаза, коллагеназа и др.).
Одни ферменты присутствуют в относительно постоянных количествах в бактериальной клетке всегда. Их называют конститутивные ферменты. Другие же ферменты начинают усиленно синтезироваться только при наличии соответствующего субстрата - индуцибельные ферменты. Так, микробная клетка начинает расщеплять лактозу в соответствующем субстрате, однако ферменты для расщепления этого сахара отсутствуют у неё при отсутствии лактозы.
© 2014-2022 — Студопедия.Нет — Информационный студенческий ресурс. Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав (0.006)
Читайте также: