Пути передачи сигналов от мозжечка. Клетка Пуркинье мозжечка
Добавил пользователь Валентин П. Обновлено: 21.01.2026
Ознакомьтесь с Условиями пребывания на сайте Форнит Игнорирование означет безусловное согласие. СОГЛАСЕН
Физиология мозжечка
Мозжечок, или малый мозг, представляет собой надсегментарную структуру, расположенную над продолговатым мозгом и мостом, позади больших полушарий мозга. Мозжечок состоит из нескольких частей, различных по происхождению в эволюции позвоночных животных.
У человека мозжечок состоит из двух полушарий, находящихся по бокам от червя. К филогенетически более древней части мозжечка млекопитающих относят переднюю долю и флоккулонодулярную часть задней доли. Эти структуры мозжечка преимущественно связаны со спинным мозгом и вестибулярным аппаратом, тогда как полушария в основном получают информацию от мышечных и суставных рецептор ов, а также от зрительного и слухового анализаторов. На рис. 5.16 представлена схема мозжечка млекопитающего (см. приложение 6), отражающая плотность вестибулярных, проприоцептивных (от мышц, сухожилий и суставов) и корковых афферентных проекций в различные зоны мозжечка. Согласно этой классификации кора мозжечка делится на три области:
1) архицеребеллум (старый мозжечок) - флоккулонодулярная доля (долька X); в ней оканчиваются преимущественно вестибулярные афференты и волокна от вестибулярных ядер; вестибулярные волокна проецируются также частично в язычок (lingula - долька I) и каудальную часть втулочки (uvula - долька IX), которые обычно относят также к архицеребеллуму;
2) палеоцеребеллум (древний мозжечок) включает переднюю долю (дольки II - V), простую дольку (долька VI) и заднюю часть корпуса мозжечка (дольки VIII-IX); палеоцеребеллум тесно связан со спинным мозгом, а также имеет двусторонние связи с сенсомоторной областью коры больших полушарий;
3) неоцеребеллум (новый мозжечок) включает среднюю часть корпуса мозжечка (долька VII и частично дольки VI и VIII), которая получает информацию от коры больших полушарий, а также от слуховых и зрительных рецептор ов. Обратите внимание, что основная часть полушарий мозжечка принадлежит новому мозжечку, который лучше всего развит у человека.
В толще мозжечка находятся три пары ядер: зубчатое, расположенное латерал ьно; ядро шатра - медиально; пробковидное и округлое ядра - между ними.
.
Единственным эфферентным выходом из коры мозжечка являются аксон ы клеток Пуркинье, образующие синапс ы с нейрон ами внутримозжечковых ядер и нейрон ами латерал ьного вестибулярного ядра (рис. 5.17). Тесная связь ядра Дейтерса с корой мозжечка дает основание рассматривать его функционально как внутримозжечковое ядро. Все остальные образования головного и спинного мозга не получают прямых эфферентов из коры мозжечка. Ядра шатра посылают волокна к ядрам Дейтерса и к ретикулярной формации продолговатого мозга. Из области ретикулярной формации, где оканчиваются пути от мозжечка, берет начало ретикулоспинальный путь. Промежуточные ядра посылают аксон ы в средний мозг, в том числе к красному ядру. Мощные пучки волокон, образованные преимущественно аксон ами нейрон ов зубчатого ядра, направляются к вентро латерал ьному ядру таламуса, где происходит синаптическое переключение, и аксон ы постсинаптических нейрон ов идут в моторную область коры больших полушарий; часть аксон ов направляется к базальным ядрам. Таким образом, мозжечок не имеет самостоятельных двигательных систем, но образует обширные связи со всеми моторными системами: кортикоспинальной (пирамидной), руброспинальной, ретику-лоспинальной, вестибулоспинальной, а также с полосатым телом.
Афферентные и эфферентные волокна мозжечка собраны в три пары массивных волокнистых пучков, известных как мозжечковые ножки. Афферентные волокна входят в мозжечок в основном через нижние и средние ножки мозжечка. Эфферентные волокна проходят преимущественно через верхние ножки. Однако имеются исключения: некоторая часть спинно-мозжечковых путей входит через верхние ножки, а некоторые эфферентные волокна от флоккулонодулярной доли и ядра шатра проходят через нижние ножки.
Ядра шатра направляют волокна через нижние ножки к вестибулярным ядрам и ретикулярной формации продолговатого мозга и моста. Промежуточные и зубчатые ядра посылают волокна через верхние ножки преимущественно к среднему мозгу и таламусу, особенно к красному ядру. Основная часть церебелло-таламических волокон отходит от зубчатых ядер. Эти волокна проецируются также к красному ядру, полосатому телу. Таким образом, влияния мозжечка на спинальные мото нейрон ы осуществляются через вестибулоспинальные и ретикулоспинальные пути, а на прецентральную область коры - через вентро латерал ьное ядро таламуса.
Строение коры мозжечка. Кора мозжечка различных представителей позвоночных, включая человека, построена по единому плану и состоит из трех слоев (см. приложение 6). Поверхностный, или молекулярный, слой содержит разветвления дендрит ов клеток Пуркинье и параллельные волокна. Клетки Пуркинье имеют уплощенный дендрит , ориентированный параллельно сагиттальным зонам долек (folia) мозжечка. Дендрит ы и аксон ы звездчатых клеток в молекулярном слое расположены таким же образом, тогда как параллельные волокна ориентированы строго трансверзально (перпендикулярно) по отношению к фоллиуму и сагиттальному направлению мшистых волокон. У кошки, например, параллельные волокна, имеют среднюю длину 2 мм (диапазон до 5-7 мм). Одно параллельное волокно пересекает около 700 дендрит ов клеток Пуркинье.
Проекции мшистых и лазающих волокон в коре мозжечка организованы в виде сагиттальных полосок. В нижней части молекулярного слоя находятся также тела корзинчатых клеток, аксон ы которых идут перпендикулярно направлению листка коры мозжечка и оплетают тела и начальные сегменты аксон ов клеток Пуркинье. В молекулярном слое имеется также небольшое число звездчатых клеток.
Самый нижний, гранулярный слой содержит клетки-зерна, или гранулярные клетки. От тела клетки-зерна отходит 4-7 коротких дендрит ных отростков, с которыми мшистые волокна образуют синапс ы. Аксон клетки-зерна поднимается вертикально вверх и в молекулярном слое Т-образно ветвится, образуя параллельные волокна. Плоскость его ветвления перпендикулярна плоскости ветвления дендрит ов клеток Пуркинье. В гранулярном слое находятся клетки Гольджи, аксон ы которых восходят в молекулярный слой. Ганглиозный слой находится между описанными выше молекулярным и гранулярным слоями и содержит тела клеток Пуркинье.
Афферентный вход в кору мозжечка осуществляется в основном через две системы волокон: лазающие и моховидные, или мшистые. Каждая клетка Пуркинье получает вход только от одного лазающего волокна (медиатор - аспартат), которое заканчивается на начальной (проксимальной) части дендрит а. Лазающие волокна представляют собой аксон ы нейрон ов, тела которых находятся в нижних оливах. По этому входу оказывается мощное возбуждающее действие: клетка Пуркинье деполяризуется на 10-15 мс, и в ней развиваются кальцийзависимые потенциал ы действия. За потенциал ом действия наступает следовая деполяризация, которая возникает вследствие активации кальцийзависимой калиевой проводимости соматической мембраны. После следовой деполяризации наступает следовая гиперполяризация. По этой причине клетка Пуркинье может возбуждаться по этому входу не более 1-2 раз в 1 с.
Моховидные волокна характеризуются обширной дивергенцией (одно волокно образует синапс ы примерно на 20 клетках-зернах) и оказывают как тормозное, так и возбуждающее действие на клетки Пуркинье. Возбудительные влияния на клетки Пуркинье от моховидных волокон переключаются через клетки-зерна, аксон ы которых поднимаются к поверхности коры мозжечка и, разветвляясь в молекулярном слое, образуют параллельные волокна. Последние оканчиваются возбудительными синапс ами (медиатор - глутамат) на дистальных участках дендрит ов клеток Пуркинье. По этому входу клетка Пуркинье может активироваться в среднем 30-40 раз в 1 с. Важным свойством этого пути является то, что активные клетки-зерна преимущественно активируют клетки Пуркинье, лежащие непосредственно над ними, т.е. эта активность незначительно распространяется по системе параллельных волокон. Отсюда можно сделать вывод о двойственной функции аксон ов клеток-зерен. С одной стороны, их восходящая часть образует радиальную организацию, благодаря которой может осуществляться локальное возбудительное влияние на клетки Пуркинье. Другая функция - модулирующая - связана собственно с параллельными волокнами. Стимуляция мшистых волокон вызывает через аксон ы клеток-зерен кортикальные ВПСП в клетках Пуркинье; они характеризуются градуальностью и последующими ТПСП. Показано, что эти ТПСП генерируются за счет активации тормозных интер нейрон ов в молекулярном слое. Синаптическая активация через параллельные волокна вызывает генерацию натриевых ПД в соме, а при большой амплитуде - генерацию дендрит ных кальциевых ПД.
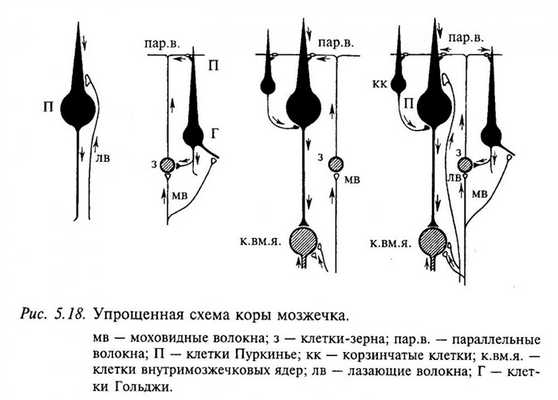
На рис. 5.18 представлена упрощенная морфофункциональная схема коры мозжечка ( нейрон ы, оказывающие тормозное действие, закрашены черным). Видно, что все входы в кору превращаются в тормозные в большинстве случаев через два переключения. Возбуждающее действие оказывают только два типа нейрон ов: клетки-зерна и нейрон ы внутримозжечковых ядер. Оба типа входных волокон (моховидные и лазающие) также являются возбуждающими. Предполагают, что такое большое количество тормозных нейрон ов в коре мозжечка необходимо для предотвращения длительной циркуляции импульсов по нервным цепям. Благодаря этому свойству мозжечок может участвовать в оперативном управлении движениями.
Функция клеток Гольджи состоит в подавлении разряда всех гранулярных клеток, которые слабо возбуждены. За счет этого осуществляется своего рода «фокусирование» ответа на те гранулярные клетки, которые сильно возбуждены через моховидные волокна. Контроль ответа гранулярной клетки осуществляется как за счет отрицательной обратной связи через клетки Гольджи, так и через синаптический вход на когтевидные дендрит ы гранулярной клетки. Как правило, каждый «коготок» дендрит а активируется отдельным моховидным волокном. Чтобы гранулярная клетка возбудилась, необходима суммация по крайней мере двух «коготков». Следовательно, только при «концентрированной» активности в моховидных волокнах можно вызвать разряд в гранулярной клетке. Аксон ы клеток Пуркинье образуют единственный выход из коры мозжечка.
.
Афферентные связи мозжечка делят на следующие группы: восходящие от спинного мозга, вестибулярные - от нижней оливы, ретикулярной формации и ядер моста. В кору мозжечка проецируются также зрительные, слуховые и вегетативные афференты. Все эти пути заканчиваются мшистыми и лазающими волокнами в коре мозжечка. Основная часть этих путей передает информацию о состоянии интер нейрон ного аппарата спинного мозга.
Кора больших полушарий, особенно кора вокруг центральной борозды, образует многочисленные прямые проекции на нейрон ы, залегающие в толще моста (так называемые мостовые ядра). Аксон ы этих нейрон ов проецируются на кору мозжечка. Эти проекции у человека очень обширны, что выражается в образовании валика на вентральной стороне моста (см. приложение 6). Эта система является основным каналом, по которому импульсация от коры больших полушарий достигает коры мозжечка, образуя проекцию как к червю, так и к полушариям мозжечка.
Влияние мозжечка на сегментарный аппарат спинного мозга.
Охлаждение передней доли мозжечка вызывает увеличение как разгибательных, так и сгибательных моносинаптических рефлексов вследствие высвобождения альфа-мото нейрон ов спинного мозга из-под тонического тормозного контроля со стороны мозжечка. Альфа-ригидность, появляющаяся после удаления мозжечка, исчезает после перерезки вестибулоспинального тракта. Это говорит о том, что клетки Пуркинье оказывают на сегментарный аппарат спинного мозга тормозное влияние. Действительно, нейрон ы ядра Дейтерса увеличивают частоту спонтанного разряда примерно в 2 раза после удаления мозжечка. Поскольку вестибулоспинальный тракт организован соматотопически, таким же образом организована регуляция мозжечком антигравитационного тонуса. Напротив, активность гамма-мото нейрон ов при охлаждении передней доли снижается. Это, в свою очередь, снижает частоту фонового разряда первичных и вторичных окончаний мышечных афферентов, а также подавляет их реакции на растяжение мышцы.
На нейрон ах вестибулярных ядер заканчиваются как аксон ы клеток Пуркинье коры червя, так и волокна, берущие начало от ядра шатра. Аксон ы клеток Пуркинье заканчиваются преимущественно на нейрон ах крупноклеточной части латерал ьного вестибулярного ядра (ядро Дейтерса), тогда как аксон ы ядра шатра - на других ядрах вестибулярного комплекса. Через вестибулоспинальные пути осуществляется фазный и тонический контроль экстензорной (антигравитационной) мускулатуры.
Удаление задней части мозжечка (флоккулюса или нодулюса) вызывает у животных дефицит вестибулярного контроля позы и движения. При этом сохраняется ощущение тяжести или движений в пространстве, отсутствуют галлюцинации движения или головокружения. Нарушаются движения глаз: появляется нистагм (обычно горизонтальный с быстрой фазой, направленной к стороне повреждения).
Установлено, что мозжечок модулирует активность нейрон ов пирамидного тракта. Основные пути к коре больших полушарий от мозжечка образованы аксон ами зубчатых и частично промежуточных ядер (через верхние ножки мозжечка). Они переключаются через вентро латерал ьное ядро таламуса, нейрон ы которого проецируются на моторную кору (см. выше). Этот путь организован соматотопически. Другой восходящий путь проходит через ретикулярную формацию ствола, куда поступают волокна от верхних и нижних ножек мозга.
Нарушения позы и движений, вызванные повреждением мозжечка.
Систематические исследования мозжечковых расстройств у животных были начаты итальянским ученым Л. Лючиани в конце XIX века. Его наблюдения были в дальнейшем многократно подтверждены, расширены и уточнены, в том числе клиницистами при изучении симптомов поражения мозжечка у людей.
Удаление мозжечка растормаживает ряд вестибулярных рефлексов и рефлексов, регулируемых сегментарными механизмами спинного мозга. У хронических собак и кошек, у которых удален мозжечок, сразу после операции усиливаются сухожильные рефлексы, гипертонус разгибателей туловища и конечностей (так называемый опистотонус). Примерно через 2 нед после операции собака без мозжечка приобретает способность стоять, и в этот период гипертонус начинает сменяться гипотонией. У обезьян удаление мозжечка сразу вызывает падение мышечного тонуса. Аналогичная картина наблюдается у больных с поврежденным мозжечком. В связи с этим больные с пораженным мозжечком не способны поддерживать определенную позу. Например, если руки больного вытянуты вперед и опираются на подставку, которую внезапно убирают, то руки пассивно падают вниз, в то время как у здорового человека они или сохраняют прежнее положение, или только слегка опускаются. По-видимому, с гипотонусом мускулатуры связано и возникновение мышечной слабости, которая проявляется в быстром наступлении усталости. Примерно в 2 раза уменьшается вес, который животное без мозжечка может нести на спине. Симптомы гипотонии объясняют подавлением активности гамма-мото нейрон ов после удаления мозжечка.
Одно из характерных проявлений мозжечковой недостаточности - это появление тремора, который проявляется в виде колебательных движений конечностей и головы в покое (статический тремор) и во время движения (кинетический, или интенционный, тремор). Обычно тремор более выражен во время движений или при эмоциональном возбуждении. При расслаблении мускулатуры, особенно во время сна, он исчезает. Тремор, который проявляется только во время мышечной активности, связывают с деятельностью гамма-системы. Кинетический тремор наиболее ярко выражен в начале движения и при его завершении, а также при перемене направления движения.
Под атаксией понимают нарушение координации, порядка (лат. taxia - порядок) движений. Наиболее сильно атаксия проявляется у приматов и особенно у человека. Принято различать статическую атаксию - нарушение равновесия при стоянии и динамическую атаксию -нарушение координации двигательных навыков. Больной с пораженным мозжечком не способен быстро сменить одно движение другим, например, барабанить пальцами.
Дисметрия (нарушение размерности движения) проявляется при совершении целенаправленных движений, когда конечность либо не достигает цели (гипометрия), либо проносится мимо нее (гиперметрия). Человек, страдающий дисметрией, не способен выполнить пальценосовую пробу (с закрытыми глазами дотронуться указательным пальцем до собственного носа). У обезьяны, обученной передвигать рычаг между двумя ограничителями, после холодового выключения зубчатого ядра резко ухудшается выполнение этого навыка: рычаг в конце каждого движения упирается в ограничитель, и проходит некоторое время, прежде чем животное начинает передвигать его в противоположном направлении (дисметрия).
Установлено, что поражение полушарных структур (кора полушарий и зубчатое ядро) приводит к атаксии конечностей, тогда как поражение червя, включая ядра шатра, вызывает атаксию туловища. Мозжечковая атаксия хорошо изучена у животных - кошек, собак, обезьян. Так, у «безмозжечковых» собак и кошек лапы широко расставлены, иногда перекрещиваются, при этом животное теряет равновесие и падает. При ходьбе или беге на тредбане амплитуда движений у них сильно варьирует от шага к шагу.
Под асинергией понимают отсутствие дополнительных содружественных движений при выполнении данного двигательного акта. Например, при попытке ходить больной заносит ногу вперед, не переместив центра тяжести, и это приводит к падению назад; при попытке сесть без помощи рук из положения, лежа изолированно сокращаются сгибатели бедра, ноги поднимаются вверх, и больной не может подняться. Больной не способен подняться со стула без помощи рук. Здоровый человек это делает в следующей последовательности: отклоняет туловище вперед, перенося тем самым центр тяжести в площадь будущей опоры, и затем встает. В результате проявления асинергии движение как бы распадается на ряд выполняемых последовательно простых движений. Так, обезьяна, у которой холодовой блокадой выключено зубчатое ядро, выполняет движение рукой, чтобы нажать кнопку, в виде последовательной серии коротких движений. Больной с поражением мозжечка, если рука поднята вверх, а его просят дотронуться до кончика носа, сначала опускает руку, затем сгибает в локте и только после этого подносит палец к носу.
Удаление флоккулонодулярной доли мозжечка у приматов, включая человека, вызывает комплекс расстройств движений глаз: гиперметрию саккад, ухудшение плавных прослеживающих движений глаз, неспособность удерживать взор в эксцентричном положении, осцилляции (тремор) глазных яблок. Из этого следует, что флоккулонодулярная доля участвует в контроле мускулатуры глаз, конечностей и туловища в ситуациях, в которых используется вестибулярный аппарат. Передняя доля мозжечка участвует в контроле локомоции, а полушария - в произвольн ых движениях. Передняя доля получает сигналы от тех областей переднего мозга, в которых формируются двигательные программы. Сюда поступает также информация от головы, шеи, туловища и конечностей. Несмотря на то что мозжечок не имеет своей собственной двигательной системы, он участвует в коррекции движений всех частей тела.
Источник: Шульговский В. В. Основы нейрофизиологии
Дата создания: 13.03.2008
Последнее редактирование: 27.03.2017
Анатомия и функции мозжечка
Тенториум был когда-то магической линией мозга. Супратенториальные регионы управляли «высшими корковыми функциями», тогда как смиренный субтенториальный мозжечок реализовал «более низкие» функции, не связанные с процессом познания. Недавние данные иллюстрируют возможную ложность этой дихотомии и привели к тому, что растущая группа нейробиологов переосмысливает мозжечок в качестве ключевого игрока в высших когнитивных функциях.
Одна линия доказательств значимости мозжечка в познании возникает из положений эволюционной нейробиологии. Две области человеческого мозга значительно больше (примерно на 1/3), чем у других высших приматов, у которых нет способностей сложного языка, формирования абстрактного понятия высокого уровня, создания произведений искусств в различных формах и сложных социальных конструкций. Один регион очевиден: префронтальная кора, другой не всегда предсказан большинством исследователей: мозжечок, причем, эти два региона работают вместе, чтобы решать множество «более высоких познавательных» задач, выполняемых мозгом.
Анатомическая поддержка «мозжечковой когнитивный теория» была представлена группой исследований , так называемой трассировки . В частности, одна из групп исследователей применяла ретроградное и антероградное трактовое картирование с вирусами герпеса и бешенства более десяти лет. В изящной серии исследований они продемонстрировали двухстороннюю связь между несколькими регионами коры ( поля 46, 12, 9 и 40, т.е. включая лобную и теменную кору) и мозжечок, связанный через мост и таламус. Эти исследования дают доказательства того, что мозжечок участвует в нейронных цепях, которые выполняют более высокие когнитивные функции , опосредованные гетеромальными ассоциативными связями. Сегодня мы знаем, что мозжечок связан со многими областями коры головного мозга кортико-мозжечковой таламо-кортикальной цепью (CCTCC).
Мозжечок имеет относительно простую архитектуру: трехслойная мозжечковая кора (молекулярная, клеточная и зернистая), корпускулярная оболочка (белое вещество) и глубокие ядра (фастигмические, шаровидные и эмболические и зубчатые ядра). Он имеет пять типов нейронов. Четыре являются ингибирующими ГАМКергическими нейронами (Purkinje, Golgi, клетками корзинчатых и звездчатых клеток), а один представляет собой возбуждающий глутаматергический нейрон (клетки гранулы).
Многие исследования показали, что основным эффектом поражений мозжечка является ухудшение двигательной координации или моторное обучение, предполагалось , что это основные задачи мозжечка. Современные исследования поражений мозжечка помогли определить и познавательную роль мозжечка, указав, что восприятие времени и нарушаются у пациентов с травмами мозжечка, что указывает на то, что мозжечок может облегчить познание, контролируя психические события в контексте времени. Кроме того, исследования по поражению мозжечка также продемонстрировали, что последний играет важную роль в ассоциативном обучении, о чем свидетельствуют исследования кондиционирования движений глазных яблок. Другие исследования показали, что поражения мозжечка могут вызывать симптомы, сходные с симптомами психических расстройств, например, такими, как мутизм. Однако, церебральные поражения обычно не вызывают психотических симптомов. Это положение может привести у мысли, что мозжечок не играет никакой роли в патогенезе шизофрении. Тем не менее, многие эксперты подтверждают классическое мнение E. Bleuler о том, что нарушения в познании являются при шизофрении первичными симптомами и что психотические симптомы являются здесь вторичными. Это изменение перспективы взгляда на мозжечок , а также растущее признание того, что мозжечок "занимается" основными когнитивными функциями, такими как синхронизация и ассоциативное обучение, привело к возникновению нового интереса к роли мозжечка при шизофрении.
Обширная работа, проделанная за последнее десятилетие с использованием инструментов in vivo нейровизуализации (neuroimaging), также продемонстрировала, что мозжечок играет значительную роль в процессах познания. Неоднократно было показано, что мозжечок активируется при множестве когнитивных процессов , даже когда двигательная активность хорошо контролируется, включая распознавание лица, атрибуцию эмоций, направленное внимание и многие типы памяти.
Мозжечок при шизофрении
Сегодня известно, что мозжечок играет роль в более высоких функциях коры. Мозжечок, понимаемый в контексте современных знаний о его связях и клеточной архитектуре, представляет интересную структуру для объяснения разнообразных симптомов шизофрении. Роль мозжечка, вероятно, не является первичной при шизофрении , в том смысле, что это единственная область, которая дисфункциональна при этом психическом расстройстве. Скорее, шизофрения, является болезнью, связанной с нарушением взаимодействий между разными структурами мозга в распределенных мозговых цепях нейронов.
Свидетельства об ухудшении возбуждающего / тормозного тонуса в мозжечке при шизофрении, в сочетании с данными исследований функциональной визуализации, показывающих нарушения мозгового кровообращения при многих типах умственной деятельности, включают мозжечок в качестве одного из важнейших сайтов где может развиться патологический процесс при шизофрении. Существует точка зрения, что аномалии мозжечка возникают при шизофрении и медленно накапливается в течение нескольких десятилетий.
Неврологические исследования предоставили важные данные о нарушениях мозжечка при шизофрении . Исследователи сообщили о сниженной линейной плотности в клетках Пуркинье или о конкретном уменьшении их размера (на 8,3%) . Последнее обнаружение особенно важно, поскольку клетки Пуркинье играют ключевую роль в модулировании выхода ( проекций) мозжечка в кору головного мозга, так как они обеспечивают вход в «глубокие ядра», такие как зубчатое ядро. Глубокие ядра, в свою очередь, обеспечивают единственный выход мозжечка в кору головного мозга.
Аномалии мозжечка при шизофрении наблюдаются как в черве, так и в полушариях мозжечка в моделях, связанных с решением задач. Пациенты с шизофренией демонстрируют снижение кровотока в мозжечке при решении широкого спектра задач, которые затрагивают различные функциональные системы мозга, включая память, внимание, социальное познание и эмоции.
Несмотря на растущие данные об аномалиях мозжечка при шизофрении, они гораздо менее обширны, чем поражения других областей мозга (например, лобной и височной коры). В настоящее время роль мозжечка в шизофрении остается несколько противоречивой, хотя противоречие частично возникает из-за приверженности консервативным моделям , касающимся функциональной активности мозжечка.
Мозжечок и психические расстройства
Мозжечок долгое время изучался исключительно в плане его роли в координации движений. Однако, исследования проведенные за последние два десятилетия показали, что мозжечок также играет ключевую роль при многих двигательных, когнитивных и эмоциональных процессах. В литературе описано несколько функций мозжечка , включая регулирование эмоций, ингибирующие импульсивное влияние на принятие решений, внимание и рабочую память. Многие экспериментальные исследования показывают , что мозжечок играет роль в процессах неконтролируемого обучения.
Кроме того, было показано, что мозжечок участвует в патогенезе многих психических расстройствах, включая расстройство гиперактивности дефицита внимания, расстройства аутистического спектра, шизофрению, биполярное расстройство, большое депрессивное расстройство и тревожные расстройства. Было высказано предположение, что двигательные, когнитивные и эмоциональные аномалии могут быть вызваны повреждением частей мозжечка, взаимодействущих с двигательной областью, префронтальной корой и лимбической системой соответственно. Некоторые авторы предполагают, что роль мозжечка в когнитивном функционировании аналогична его контролю над целенаправленными двигательными навыками во время соверешения движений.
Из мозжечка выходят три тракта: (1) пространство мозжечка - его червь , косвенно , имеет отношение к мосту мозгу и ретикулярной формации; (2) промежуточная зона мозжечка направляется к красному ядру и таламусу; и (3) боковая зона мозжечкового полушарий мозжечка идет по направлению к таламусу. После входа в зрительный бугор ( таламической связи) эти волокна проецируются в разные части коры головного мозга, включая лобную, моторную и теменную области коры. Кортико-понто-мозжечковые и мозжечково-таламо-корковые пути позволяют мозжечку влиять на обработку информации в областях коры, ответственные за когнитивные и эмоциональные процессы. Эти сложные связи между мозжечком и другими структурами могут объяснить, почему повреждение мозжечка может привести к различным психическим расстройствам.
До сих пор мало что известно о том, как мозг развивается у пациентов с СДВГ в ходе течения этого психического расстройства. Исследователи обнаружили объемные аномалии с уменьшенным размером головного мозга и мозжечка, которые увеличивались с возрастом, в то время как изменения объема хвостатого ядра исчезали по мере того, как испытуемые становились старше. Было установлено, что эти результаты не связаны с лечением психостимуляторами. В то же время было показано , что у пациентов, получающих лечение стимуляторами, отмечается больший общий объем мозжечка, который превышает размеры мозжечка детей с СДВГ , не принимавших стимуляторы. Размер червя (vermis) также может предсказать результаты терапии ( меньшие объемы прогнозировали более плохие результаты). Кроме того, у пациентов с меньшими по размеру дольками червя, вызванных инсультом или другими аномалиями развития, также проявляется способность к плохой концентраци внимания и ориентировке в окружающей среде. В целом, сокращение объема мозжечка является распространенной находкой в исследованиях, посвященных расстройству мозжечка и СДВГ. Однако, мы не можем определить, присутствовали ли аномалии в мозжечке с рождения или развивались во время роста ребенка, и как это влияет на этиологию СДВГ.
Мозжечок способен влиять на моторную кору и область префронтальной коры, две области, которые отвечают за моторный контроль и социальное познание, поэтому неудивительно, что аномалии в мозжечке могут вызвать симптомы, наблюдаемые при расстройствах аутистического спектра (ASD). Уменьшение активности клеток Пуркинье приводит к поведению, подобному ASD, включая аномальные социальные и двигательные формы поведения. Посмертные исследования также показали снижение плотности клеток Пуркинье у пациентов с ASD. Являясь гамкергическими клетки Пуркинье влияют на активность тракта " мозжечок-кора", что может объяснить возникновение стереотипных (повторных) движений при ASD. Это, однако, должно быть подтверждено или неподтверждено в исследованиях, которые связывают потерю клеток Пуркинье с точными доменами симптомов (двигательная и социальная дисфункция) при ASD. Снижение активности в ножках мозжечка также связано с более слабыми двигательными способностями ( навыками) у пациентов с ASD. Существует предположение о дополнительном дефекте в формировании сетей нейронов мозжечка и фронтальной доли при синдроме Аспергера. Эти дефициты могут быть причиной моторных и когнитивных нарушений, наблюдаемых у пациентов с ASD. Нарушение адаптации социального поведения у пациентов с ASD может быть вызвано нарушением путей обратной связи от мозжечка до коры головного мозга. Кроме того, были выявлены аномально организованные волокна среднего и нижнего мозжечковых стеблей ( ножек) , соединяющих мозжечок с лобной долей. Это может быть либо прямой причиной, либо следствием изменений в коре головного мозга и мозжечковых ядрах у пациентов с аутизмом. Эти патологические изменения объясняют дефицит координации и атаксию при некоторых формах аутизма. Таким образом, в настоящее время наблюдаются три основных нарушения мозжечка у пациентов с ASD: уменьшенные в размерах клетки Пуркинье, уменьшение объема мозжечка и нарушения ( разрывы) обратной связи между мозжечком и областями коры мозга. Поскольку клетки Пуркинье являются ингибирующими по своей природе, недостаток этих клеток уменьшает ингибирование, которое мозжечок направляет в кортикальные и подкорковые области, что приводит к гиперчувствительности этих областей мозга, обнаруженных у большинства детей с аутизмом.
Большинство исследований на сегодняшний день сосредоточено на клетках Пуркинье при ASD , в частности, на синдроме Аспергера или аутизме; однако было бы полезнопонять, как плотность клеток Пуркинье связана с выраженностью аутизма. Поскольку клетки Пуркинье ингибируют области коры головного мозга и области среднего мозга, можно предположить , что у пациентов с выраженным аутизмом также будет наблюдаться гораздо более низкая плотность клеток Пуркинье.
Структурные исследования изображений головного мозга выявили снижение объема мозжечка у больных шизофренией, в том числе уменьшенный объем червя мозжечка. Изменения объема мозжечка у пациентов с шизофренией были связаны: с нарушениями поведения, возникающими в перинатальном периоде; мужским полом, ранним дебютом и манифестацией шизофренией , хроническим течением заболевания и клинической картиной с преимуществено позитивными симптомами. Функциональные исследования изображений мозга у пациентов с шизофренией свидетельствуют о снижении кровотока в коре головного мозга и черве мозжечка во время решения многих когнитивных задач, на внимание, память, включая задачи как краткосрочной, так и рабочей памяти и социальная активность.
Исследования, касающиеся роли мозжечка в моторных побочных эффектах, наблюдаемых у пациентов с шизофренией, находящихся на терапии антипсихотическими препаратами, ограничены. Например, одно исследование показало снижение активности мозжечка у больных шизофренией с симптомами акатезии во время лечения оланзапином, однако не известно, как изменения функции мозжечка могут привести к этому осложению терапии нейролептиками. Меньший объем мозжечка в ASD можно объяснить уменьшением числа клеток Пуркинье; однако, количество клеток Пуркинье не отличаются у здоровых людей и пациентов с шизофренией. Это означает, что уменьшение объема мозжечка при шизофрении, возможно, связана с уменьшением или отсутствием различных его частей.
Многие исследования демонстрируют изменения мозжечка с уменьшением его объема и атрофией у пациентов с биполярным расстройством. Интересно, что объем V3-красной зоны субрегиона мозжечка значительно снижается у пациентов с множественными эпизодами биполярного расстройства по сравнению с данными , полученными на здоровом контролем, в то время как объем V2-красной зоны субрегиона меньше у пациентов с множественными эпизодами, чем у пациентов с первым эпизодом. Также было обнаружено, что уменьшение объема мозжечка выражено больше у пациентов редко принимавшими лекарства по сравнению с пациентами, находящимися на активном лечении препаратами в течение длительного периода времени. В исследовании, использующем функциональную МРТ у пациентов с BD, в мозжечке больных, которые были устойчивы к лечению, был обнаружен повышенный метаболизм глюкозы. Однако неясно, являются ли эти изменения в церебральном кровотоке и метаболизме первичными или вторичными по отношению к BD . Из-за ингибирующего характера активности мозжечка можно ожидать , что активация этой структуры будет уменьшаться во время маниакальных фаз и увеличиваться во время фаз депрессии. Альтернативно, активация из мозжечка может оставаться постоянной, в то время как остальная часть мозга работает как бы "на автомате", пытаясь компенсировать отклоняющую ингибирующую активацию, исходящую из мозжечка.
В литературе сообщается о нарушениях функций мозжечка при тревожных расстройствах , что может быть связано с увеличением возбуждения при посттравматическом стрессовом расстройстве (PTSR), генерализованном тревожном расстройстве (GAD) и социальном тревожном расстройстве (SAD).
Исследование , проведенное с помощью однофотонная эмиссионная компьютерная томография (SPECT) Bonne et al.( 2003) выявило повышенную активность мозжечка при повторном травматическом событии у пациентов с посттравматическим стрессовым расстройством ( PTSR). Гиперчувствительность мозжечка положительно коррелировала с повышенным кровяным давлением и частотой сердечных сокращений, указывая на возможную роль мозжечка в регуляции симпатической активности, что может объяснить его роль в патогенезе тревожных расстройств. Исследования пациентов с паническим расстройством выявили значительные изменения уровни метаболизма глюкозы в мосту, среднем мозге, таламусе, гиппокампе, миндалине и мозжечке.
Было обнаружено, что мозжечок ассоциируется не только с психическими и когнитивными симптомами при различных психических и неврологических расстройствах, но также с особенностями фармакологической и когнитивно - поведенческой терапии. Однако до сих пор неясно, как дисфункция мозжечка связана с различными симптомами при психических расстройствах. Большинство результатов исследований неубедительны при рассмотрении конкретных анатомических аномалий в мозжечке, которые присутствуют при психических расстройствах. Тем не менее, некоторые из расстройств, обсуждавшихся с аналогичными нарушениями мозжечка, например, ASD, шизофрения, биполярные и MDD, показывают снижение объема в области vermis; однако их симптомы значительно отличаются друг от друга. Поскольку каждая область мозжечка проецирует свое влияние на разные области коры головного мозга и в средние структуры мозга , разнообразие симптомов указывает на то, что отклонения от каждого расстройства сосредоточены на определенных областях, а не на мозжечке в целом. Это может объяснить широкий спектр симптомов, наблюдаемых при нарушениях , выявленных в мозжечке. Например, сильная связь между областями vbis VIIb и IX и визуальной сетью была отмечена Sang et.al ( 2012) . Известно также, что эта область влияет на кровоток у пациентов с шизофренией, что, в свою очередь, может быть фактором, связанным с визуальными галлюцинациями. То же самое можно сказать и о связи с полушариями VI, VIIb и VIII, которые показывают связь с "слуховой сетью" нейронов и могут объяснить слуховые галлюцинации, имеющиеся у некоторых пациентов с шизофренией.
Нейронауки для всех. Детали: клетки Пуркинье
Наша рубрика «Нейронауки для всех» уже имеет два подраздела. Первый — это большие статьи, посвящённые общим вопросам: как появляется нервная система, как она устроена, и так далее. Второй — это статьи, посвящённые инструментам и методам нейронаук. МРТ, КТ, ТМС и другие страшные аббревиатуры. Однако, в этом наборе совершенно точно не хватает ещё одной, самой толстой колоды карт: клеточно-анатомической. Ведь нервная система состоит из большого количества типов клеток, а мозг имеет много различных отделов, которые постоянно упоминаются в наших новостях и статьях. Поэтому мы начинаем и третий цикл статей, который мы условно назвали «Детали». И первая статья этого цикла будет посвящена одному из самых известных типов клеток в мозжечке (структуре, статья о которой еще впереди): клеткам Пуркинье.
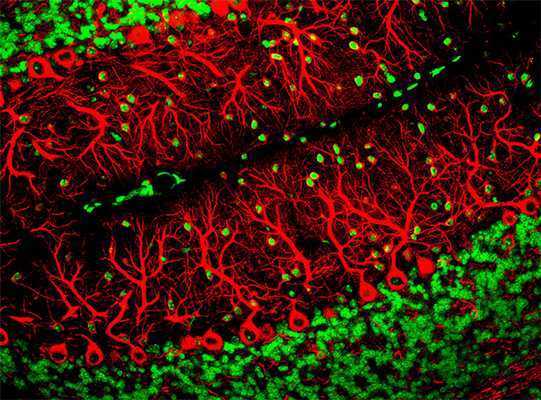
Клетки Пуркинье (красный). Фото Yinghua Ma and Timothy Vartanian, Cornell University, Ithaca, N.Y. Part of the exhibit Life:Magnified by ASCB and NIGM
История
Впервые, как несложно догадаться, клетки Пуркинье увидел человек по фамилии Пуркинье. Или Пуркине, как любят говорить пуристы. Чех Ян Эвангелиста Пуркинье был потрясающим человеком, учёным-энциклопедистом, переписывался с Гёте, состоял в ордене иллюминатов, открыл сумеречное зрение, стал одним из родоначальников дактилоскопии и создал прототип киноаппарата, несмотря на то, что был старшим современником Пушкина. Он прожил долгую жизнь, и анатомией занимался чуть более двух десятков лет. В 1837 году он описал «ганглиозные тельца» — клетки Пуркинье.

Ян Эвангелиста Пуркинье
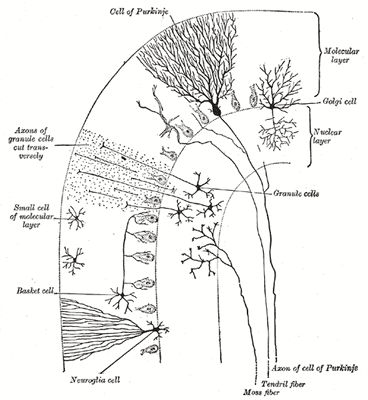
Самое известное изображение этих ветвистых клеток получил другой великий учёный — нобелевский лауреат 1906 года, Сантьяго Рамон-и-Кахаль. На его знаменитом рисунке, растиражированном во всех учебниках, изображены клетки Пуркинье и более глубокие гранулярные клетки в мозжечке голубя.
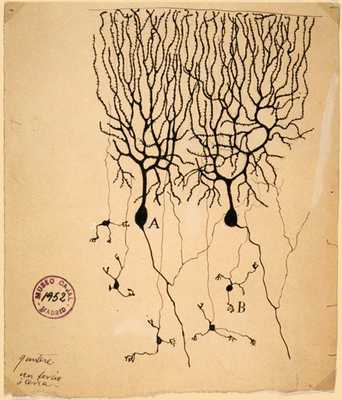
Клетки Пуркинье (А) и гранулярные клетки (В) мозжечка голубя. Рисунок Сантьяго Рамон-и-Кахаля
Только факты
Клетки Пуркинье — это ГАМК-эргические (передающие сигнал при помощи нейромедиатора гамма-аминомасляной кислоты) униполярные нейроны (с одним аксоном). Длина аксона у мышей - 2 миллиметра, у крыс — 3 миллиметра. Толщина дендритов 2-5 мкм — толстые ветви, 0.5-1 мкм — тонкие. Дендритная сетка у клеток Пуркинье очень развита, плотность дендритных шипиков тоже высокая. В итоге, каждая клетка Пуркинье способна образовать до двухсот тысяч синапсов! Это очень много для нейрона.
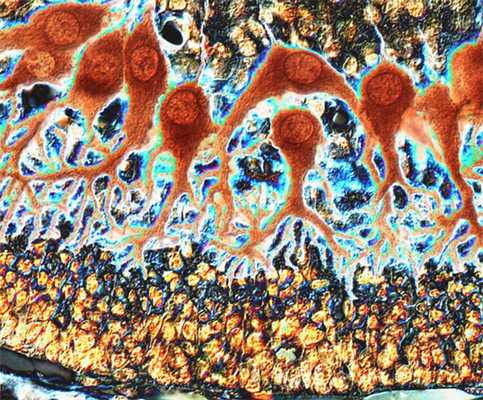
Клетки Пуркинье мозжечка мыши
Credit: Jakob Jankowski
University of Bonn
Department of Anatomy and Cell Biology
Bonn, Germany
Клетки Пуркинье — одни из самых крупных нейронов (если не считать длину аксонов) в человеческом мозге. Крупнее только клетки Беца, о которых речь ещё впереди.
Мозжечок. Credit: Anatomography maintained by Life Science Databases(LSDB).
Всего клеток Пуркинье в мозжечке 26 миллионов. Они, вместе с особыми глиальными клетками, так называемой глией Бергманна, образуют слой Пуркинье в мозжечке. Удивительно, что разветвлённая дендритная сеть каждой клетки почти двумерна, подобно опахалу или кораллу, и в слое Пуркинье одноименные клетки ориентированы параллельно, как костяшки домино в различных фокусах с падением последних. Сквозь эти двумерные слои дендритов проходят перпендикулярно им волокна аксонов гранулярных клеток из более глубоких слоев мозжечка и образуют с дендритами клеток Пуркинье синапсы (так называемые gcPc-cинапсы, от Granule-cell to Purkinje-cell synapses). Соединение гранулярных клеток и клеток Пуркинье изображено на том самом известном рисунке с клетками Пуркинье, выполненным Рамон-и-Кахалем.
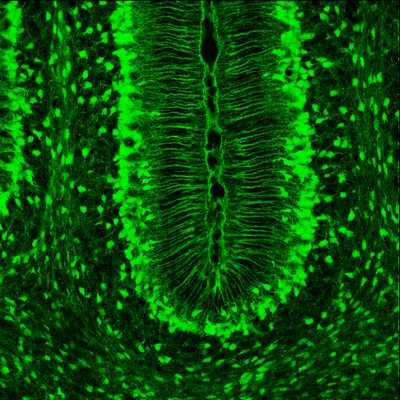
«Разрез» дольки мозжечка с типами клеток
Тело клетки Пуркинье — грушевидное, а длинный аксон уходит сквозь белое вещество к ядрам мозжечка и вестибулярным ядрам.
Происхождение
Если говорить о происхождении клеток Пуркинье во время формирования организма (у нас есть отдельная статья про то, как формируется нервная система), то есть данные, что эти клетки происходят от общего стволового «предка» вместе с совершенно непохожими на них B-лимфоцитами и альдостерон-продуцирующими клетками коры надпочечника (!).
Зачем они нужны
Роль клеток Пуркинье в том, как мы двигаемся, очень сложно переоценить. Они получают возбуждающие импульсы от лиановидного волокна и моховидных (мшистых) волокон мозжечка и отправляют тормозные импульсы (мы же помним, что ГАМК - основной «тормоз» головного мозга) в глубокие слои мозжечка - его ядра. Если перевести эту активность на простой язык, то клетки Пуркинье играют важнейшую роль в двигательном обучении, в равновесии и коодинации движений. Убедиться в этом просто, зная два факта: во-первых, у человека клетки Пуркинье вызревают сравнительно поздно, к восьми годам жизни человека, а во-вторых, они очень чувствительны к воздействию алкоголя. И именно поэтому дети и пьяные движутся так неуклюже. Кстати, самые развитые клетки Пуркинье — именно у людей, с детства занимающихся сложнокоординированным движением: акробатикой, гимнастикой, фигурным катанием или танцами.
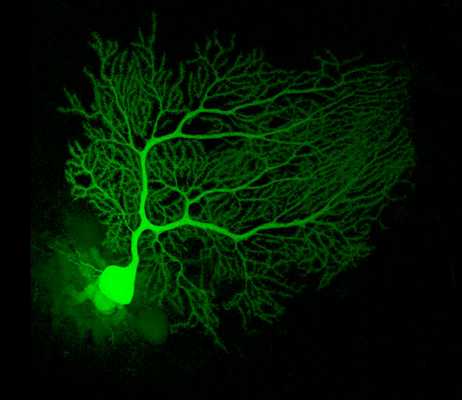
Одиночная клетка Пуркинье
Текст: Алексей Паевский
Читайте материалы нашего сайта в Facebook, ВКонтакте, Яндекс-Дзен и канале в Telegram, а также следите за новыми картинками дня в Instagram.
Читайте также:
- Резистентность к антибиотикотерапии. Эффективность пенициллина в лечении менингита в прошлом
- Хроническая головная боль при цереброваскулярных заболеваниях. Атеросклеротическая транзиторная ишемическая атака.
- Роль световой и темновой адаптации. Цветовое зрение
- Нейрофиброма у ребенка
- Рекомендации по химиотерапии у беременной