Торможение и возбуждение ядер мозжечка. Физиология сигнальной системы мозжечка
Добавил пользователь Cypher Обновлено: 28.01.2026
Мозжечок регулирует силу и точность мышечных сокращений и их тонус как в покое, так и при движениях, а также синергию сокращений разных мышц при сложных движениях. При поражении мозжечка возникает целый ряд как двигательных расстройств, так и нарушений со стороны вегетативной нервной системы. К ним относятся астения - быстрая утомляемость, снижение силы мышечных сокращений, астазия - утрата способности к длительному сокращению мышц, поэтому больной не может долго стоять или сидеть, неточность движений; атаксия - нарушение координации движений, неуверенная походка; абазия - невозможность сохранить центр тяжести тела; атония или дистония - понижение или повышение тонуса мышц; тремор - дрожание пальцев рук, кистей и головы в покое, но его усиление при движении; дизартрия - нарушение кординации мышц лица, необходимое для четкого произнесения слов. Речь становится монотонной, медленной и невыразительной. Дисметрия - расстройство равномерности движения (гиперметрия или гипометрия).
Страдает кожная и проприоцептивная чувствительность нижних (пучок Флексига) и верхних конечностей (пучок Говерса). При поражении мозжечка тормозится процесс обучения, так как многие движения человек заучивает во время жизни (письмо, ходьба).
Нарушение функций мозжечка приводит к неточности движений, их разбросанности, негармоничности, а это говорит об участии мозжечка в деятельности коры больших полушарий, ответственной за организацию процессов высшей нервной деятельности.
Мозжечок влияет на возбудимость сенсомоторной коры больших полушарий и контролирует тем самым уровень тактильной, температурной и зрительной чувствительности.
Мозжечок играет адаптационно-трофическую роль в регуляции не только мышечной деятельнсти, но и ее вегетативного обеспечения. Удаление мозжечка приводит к нарушению целого ряда вегетативных функций (Л. А. Орбели). Мозжечок оказывает как угнетающее, так и стимулирующее влияние на работу сердечно-сосудистой системы. При раздражении последнего высокое артериальное давление снижается, а исходное низкое - повышается. Снижается частота дыхания, повышается тонус гладких мышц кишечника. При повреждении мозжечка нарушается углеводный, белковый и минеральный обмен, а также процессы энергообразования, терморегуляции и кроветворения. Стимуляция мозжечка приводит к нарушению репродуктивной функции, к сокращению матки у беременных кошек. Удаление мозжечка провоцирует рождение нежизнеспособного потомства или препятствует зачатию.
6. Строение и функции мозжечка
6. Строение и функции мозжечка Мозжечок является центром координации движения. Он расположен в задней черепной ямке вместе со стволом мозга. Крышей задней черепной ямки служит намет мозжечка. Мозжечок имеет три пары ножек.Эти ножки образованы мозжечковыми проводящими
7. Виды поражения мозжечка
7. Виды поражения мозжечка При поражении архицеребеллума отмечается нарушение равновесия при стоянии и ходьбе - мозжечковая атаксия. При закрытии глаз мозжечковая атаксия не изменяется. Причиной ее возникновения является асинергия мышц. Эфферентные импульсы от
19. Поражение артерии продолговатого мозга и нижней задней артерии мозжечка
19. Поражение артерии продолговатого мозга и нижней задней артерии мозжечка Парамедианные артерии в оральном отделе продолговатого мозга отходят от позвоночных артерий, в каудальном отделе - от передней спинальной артерии. Они снабжают кровью пирамидный путь,
НАМЁТ МОЗЖЕЧКА
НАМЁТ МОЗЖЕЧКА Ряд патологий, связанных с нарушениями в высшей нервной деятельности, может зависеть от намёта мозжечка, изменение натяжения которого оказывает патологическое влияние на прилегающие ткани, мозговые оболочки и, опосредованно, на организм человека в
Опухоли головного мозга и мозжечка
Опухоли головного мозга и мозжечка Опухоли головного мозга и мозжечка, создавая дополнительный объем внутри замкнутой полости, вызывают нарушения венозного оттока и оттока спинномозговой жидкости из полости черепа — возникает головная боль диффузного характера
Опухоли головного мозга и мозжечка Опухоли головного мозга и мозжечка, создавая дополнительный объем внутри замкнутой полости, вызывают нарушения венозного оттока и оттока спинномозговой жидкости из полости черепа - возникает головная боль диффузного характера
Максимизируйте функции мышц, минимизируя функции жировых тканей
Максимизируйте функции мышц, минимизируя функции жировых тканей Этот принцип можно применить к обширному комплексу метаболических процессов, которые принимают решение, произойдут ли рост мускулов и потеря жира. Этот принцип приводит к пониманию, какие процессы следует
Строение коры мозжечка. Процессы возбуждения и торможения в нейронных сетях мозжечка
Над четвертым желудочком, являющимся по существу полостью моста и продолговатого мозга, находится мозжечок, или, как его называют, «малый мозг».
Мозжечок выполняет функции управления движениями — как целенаправленными произвольными, так и быстрыми автоматизированными — регуляция позы, локомоции, мышечного тонуса, поддержание равновесия тела. У мозжечка различают два выпуклых полушария и червь — непарную срединную часть. Поверхности полушарий и червя разделяют поперечные параллельные борозды (щели), между которыми расположены узкие и длинные листки мозжечка. Благодаря этому его поверхность у взрослого человека составляет в среднем 850 см2. У мозжечка различают верхнюю и нижнюю поверхности, между которыми по заднему краю проходит глубокая горизонтальная щель. В боковых отделах горизонтальная щель берет начало у места вхождения в мозжечок его средних ножек. Группы листков, разделенные глубокими бороздами, образуют дольки мозжечка. Поскольку борозды мозжечка сплошные и переходят с червя на полушария, каждая долька червя связана с правой и левой стороны с дольками полушарий. На разрезе мозжечок состоит из серого и белого вещества. Серое вещество (кора мозжечка) находится на поверхности и тонким слоем (1—2,5 мм) покрывает белое вещество. Белое вещество находится внутри мозжечка.
У коры мозжечка три слоя: наружный — молекулярный, средний — ганглионарный (слой грушевидных нейронов) и внутренний — зернистый. В молекулярном и зернистом слоях залегают в основном мелкие нейроны. Крупные грушевидные нейроны (клетки Пуркинье), размерами до 40 мкм, располагаются в среднем слое в один ряд. Это эфферентные нейроны коры мозжечка. Их аксоны направляются к нейронам ядер мозжечка, а дендриты располагаются в поверхностном молекулярном слое. Остальные нейроны коры мозжечка являются вставочными, ассоциативными, которые передают импульсы грушевидным нейронам. В толще белого вещества мозжечка имеются скопления серого вещества — парные ядра. Самое крупное, зубчатое ядро расположено в пределах полушария мозжечка и получает сигналы от клеток Пуркинье новой коры мозжечка. Медиальнее зубчатого ядра лежит пробковидное, еще медиальнее — шаровидное, которые объединяют в промежуточное ядро мозжечка, получающее проекции от его старой коры. Наиболее медиально находится ядро шатра, связанное с клетками Пуркинье древней коры мозжечка. Афферентные и эфферентные волокна, связывающие мозжечок с другими отделами мозга, образуют три пары мозжечковых ножек. Нижние ножки соединяют мозжечок с продолговатым мозгом, средние — с мостом, верхние — со средним мозгом.
К мозжечку направляются восходящие (чувствительные) проводящие пути, по которым идут импульсы от мышц, сухожилий, капсул суставов, связок (спинно-мозжечковые пути, а также волокна от чувствительных ядер тройничного нерва).
В мозжечок приходят также импульсы от вестибулярных ядер, из коры полушарий большого мозга (через собственные ядра моста), а также из олив. Из мозжечка выходят пучки нервных волокон ко многим отделам ЦНС.
Имея обширные нервные связи с различными отделами мозга, мозжечок участвует в регуляции движений, делает их плавными, точными, целенаправленными. При этом древняя кора и ядра шатра связаны с поддержанием равновесия; старая кора и промежуточные ядра — со сгибанием и разгибанием конечностей и локомоцией; новая кора и зубчатые ядра — с тонкими движениями кисти и пальцев. При повторении движений все эти зоны способны запоминать их параметры. В результате движения, исходно осуществляемые как произвольные и требующие контроля коры больших полушарий, превращаются в автоматизированные, идущие в основном под контролем мозжечка (двигательное обучение).
При повреждении мозжечка, выпадении его функции нарушается распределение тонуса мышц — сгибателей и разгибателей; движения становятся несоразмерными, резкими, размашистыми. Нарушается анализ сигналов от рецепторов мышц и сухожилий, страдают вегетативные функции органов сердечно-сосудистой системы, пищеварительных и других органов (Относится также к билету 16, вопрос 3)
Взаимодействие входящих волокон с клетками коры мозжечка
В кору мозжечка входят два типа волокон: Лазающие (лиановидные) - аксоны клеток, тела которых находятся в нижней оливе (промежуточное вестибулярное ядро продолговатого мозга), проходят через зернистый слой и оплетают дендриты клеток Пуркинье в молекулярном слое.
Мшистые волокна (основные афференты) -аксоны различных стволовых структур, которые сами иннервируют разные клетки коры мозжечка (дивергенция) и посылают сигналы через множество параллельных волокон зернистых клеток к каждой клетке Пуркинье (конвергенция)
Вопрос 32. Физиология мозжечка, его влияние на моторику и вегетативные функции организма.
Анатомически в мозжечке можно выделить среднюю часть - червь, полушария мозжечка и боковые флокулонодулярные доли.
В мозжечке выделяют три слоя:
I - молекулярный слой. В этом слое располагаются дендритные разветвления грушевидных нейронов. В молекулярного слоя находятся тела корзинчатых клеток, аксоны которых образуют синапсы с телами грушевидных нейронов, и некоторое число звездчатых клеток.
II - ганглиозный слой. Здесь располагаются тела грушевидных нейронов.
III - гранулярный слой. В этом слое находятся тела вставочных нейронов (клеток-зерен). Аксоны клеток-зерен поднимаются в молекулярный слой, где они Т-образно разветвляются. Здесь же находятся клетки Гольджи, аксоны которых также направляются в молекулярный слой.
Мшистые волокна возбуждают клетки-зерна, а те через параллельные волокна оказывают возбуждающее действие на все остальные нейроны, которые, однако, всегда являются тормозными. Таким образом, все клетки, нейроны, тела которых лежат в коре мозжечка, кроме клеток-зерен, выполняют тормозные функции.
В состав мозжечка входят следующие ядра: ядро Шатра, вставочное ядро (у человека это ядро состоит из пробкового и шаровидного ядер) и зубчатое ядро.
Афферентные и эфферентные связи мозжечка. Афферентные и эфферентные связи мозжечка очень многочисленные. Мозжечок получает афферентные связи от вестибулярных ядер, соматосенсорные пути, главным образом от спинного мозга, нисходящие пути - в основном от коры головного мозга.
От спинного мозга к мозжечку подходят два спиномозжечковых пути - вентральный и дорсальный (Говерса и Кларка).
Эфферентные связи. Кора червя мозжечка посылает пути к ядру Шатра, средняя часть коры мозжечка - к вставочному ядру, а кора полушарий мозжечка - к зубчатому ядру. Пути от каждого из этих ядер поступают к различным образованиям ствола мозга и больших полушарий. Таким образом, ядра мозжечка являются выходными структурами мозжечка.
Функции мозжечка
В результате сложилось мнение, что основное значение мозжечка состоит в дополнении и коррекции деятельности остальных двигательных центров. Мозжечок отвечает за регуляцию позы и мышечного тонуса, за исправление медленных целенаправленных движений в ходе их выполнения и координацию этих движений с рефлексами поддержания позы, правильное выполнение быстрых целенаправленных движений, команда к которым поступает от коры больших полушарий головного мозга.
Торможение и возбуждение ядер мозжечка. Физиология сигнальной системы мозжечка
Ознакомьтесь с Условиями пребывания на сайте Форнит Игнорирование означет безусловное согласие. СОГЛАСЕН
Физиология мозжечка
Мозжечок, или малый мозг, представляет собой надсегментарную структуру, расположенную над продолговатым мозгом и мостом, позади больших полушарий мозга. Мозжечок состоит из нескольких частей, различных по происхождению в эволюции позвоночных животных.
У человека мозжечок состоит из двух полушарий, находящихся по бокам от червя. К филогенетически более древней части мозжечка млекопитающих относят переднюю долю и флоккулонодулярную часть задней доли. Эти структуры мозжечка преимущественно связаны со спинным мозгом и вестибулярным аппаратом, тогда как полушария в основном получают информацию от мышечных и суставных рецептор ов, а также от зрительного и слухового анализаторов. На рис. 5.16 представлена схема мозжечка млекопитающего (см. приложение 6), отражающая плотность вестибулярных, проприоцептивных (от мышц, сухожилий и суставов) и корковых афферентных проекций в различные зоны мозжечка. Согласно этой классификации кора мозжечка делится на три области:
1) архицеребеллум (старый мозжечок) - флоккулонодулярная доля (долька X); в ней оканчиваются преимущественно вестибулярные афференты и волокна от вестибулярных ядер; вестибулярные волокна проецируются также частично в язычок (lingula - долька I) и каудальную часть втулочки (uvula - долька IX), которые обычно относят также к архицеребеллуму;
2) палеоцеребеллум (древний мозжечок) включает переднюю долю (дольки II - V), простую дольку (долька VI) и заднюю часть корпуса мозжечка (дольки VIII-IX); палеоцеребеллум тесно связан со спинным мозгом, а также имеет двусторонние связи с сенсомоторной областью коры больших полушарий;
3) неоцеребеллум (новый мозжечок) включает среднюю часть корпуса мозжечка (долька VII и частично дольки VI и VIII), которая получает информацию от коры больших полушарий, а также от слуховых и зрительных рецептор ов. Обратите внимание, что основная часть полушарий мозжечка принадлежит новому мозжечку, который лучше всего развит у человека.
В толще мозжечка находятся три пары ядер: зубчатое, расположенное латерал ьно; ядро шатра - медиально; пробковидное и округлое ядра - между ними.
.
Единственным эфферентным выходом из коры мозжечка являются аксон ы клеток Пуркинье, образующие синапс ы с нейрон ами внутримозжечковых ядер и нейрон ами латерал ьного вестибулярного ядра (рис. 5.17). Тесная связь ядра Дейтерса с корой мозжечка дает основание рассматривать его функционально как внутримозжечковое ядро. Все остальные образования головного и спинного мозга не получают прямых эфферентов из коры мозжечка. Ядра шатра посылают волокна к ядрам Дейтерса и к ретикулярной формации продолговатого мозга. Из области ретикулярной формации, где оканчиваются пути от мозжечка, берет начало ретикулоспинальный путь. Промежуточные ядра посылают аксон ы в средний мозг, в том числе к красному ядру. Мощные пучки волокон, образованные преимущественно аксон ами нейрон ов зубчатого ядра, направляются к вентро латерал ьному ядру таламуса, где происходит синаптическое переключение, и аксон ы постсинаптических нейрон ов идут в моторную область коры больших полушарий; часть аксон ов направляется к базальным ядрам. Таким образом, мозжечок не имеет самостоятельных двигательных систем, но образует обширные связи со всеми моторными системами: кортикоспинальной (пирамидной), руброспинальной, ретику-лоспинальной, вестибулоспинальной, а также с полосатым телом.
Афферентные и эфферентные волокна мозжечка собраны в три пары массивных волокнистых пучков, известных как мозжечковые ножки. Афферентные волокна входят в мозжечок в основном через нижние и средние ножки мозжечка. Эфферентные волокна проходят преимущественно через верхние ножки. Однако имеются исключения: некоторая часть спинно-мозжечковых путей входит через верхние ножки, а некоторые эфферентные волокна от флоккулонодулярной доли и ядра шатра проходят через нижние ножки.
Ядра шатра направляют волокна через нижние ножки к вестибулярным ядрам и ретикулярной формации продолговатого мозга и моста. Промежуточные и зубчатые ядра посылают волокна через верхние ножки преимущественно к среднему мозгу и таламусу, особенно к красному ядру. Основная часть церебелло-таламических волокон отходит от зубчатых ядер. Эти волокна проецируются также к красному ядру, полосатому телу. Таким образом, влияния мозжечка на спинальные мото нейрон ы осуществляются через вестибулоспинальные и ретикулоспинальные пути, а на прецентральную область коры - через вентро латерал ьное ядро таламуса.
Строение коры мозжечка. Кора мозжечка различных представителей позвоночных, включая человека, построена по единому плану и состоит из трех слоев (см. приложение 6). Поверхностный, или молекулярный, слой содержит разветвления дендрит ов клеток Пуркинье и параллельные волокна. Клетки Пуркинье имеют уплощенный дендрит , ориентированный параллельно сагиттальным зонам долек (folia) мозжечка. Дендрит ы и аксон ы звездчатых клеток в молекулярном слое расположены таким же образом, тогда как параллельные волокна ориентированы строго трансверзально (перпендикулярно) по отношению к фоллиуму и сагиттальному направлению мшистых волокон. У кошки, например, параллельные волокна, имеют среднюю длину 2 мм (диапазон до 5-7 мм). Одно параллельное волокно пересекает около 700 дендрит ов клеток Пуркинье.
Проекции мшистых и лазающих волокон в коре мозжечка организованы в виде сагиттальных полосок. В нижней части молекулярного слоя находятся также тела корзинчатых клеток, аксон ы которых идут перпендикулярно направлению листка коры мозжечка и оплетают тела и начальные сегменты аксон ов клеток Пуркинье. В молекулярном слое имеется также небольшое число звездчатых клеток.
Самый нижний, гранулярный слой содержит клетки-зерна, или гранулярные клетки. От тела клетки-зерна отходит 4-7 коротких дендрит ных отростков, с которыми мшистые волокна образуют синапс ы. Аксон клетки-зерна поднимается вертикально вверх и в молекулярном слое Т-образно ветвится, образуя параллельные волокна. Плоскость его ветвления перпендикулярна плоскости ветвления дендрит ов клеток Пуркинье. В гранулярном слое находятся клетки Гольджи, аксон ы которых восходят в молекулярный слой. Ганглиозный слой находится между описанными выше молекулярным и гранулярным слоями и содержит тела клеток Пуркинье.
Афферентный вход в кору мозжечка осуществляется в основном через две системы волокон: лазающие и моховидные, или мшистые. Каждая клетка Пуркинье получает вход только от одного лазающего волокна (медиатор - аспартат), которое заканчивается на начальной (проксимальной) части дендрит а. Лазающие волокна представляют собой аксон ы нейрон ов, тела которых находятся в нижних оливах. По этому входу оказывается мощное возбуждающее действие: клетка Пуркинье деполяризуется на 10-15 мс, и в ней развиваются кальцийзависимые потенциал ы действия. За потенциал ом действия наступает следовая деполяризация, которая возникает вследствие активации кальцийзависимой калиевой проводимости соматической мембраны. После следовой деполяризации наступает следовая гиперполяризация. По этой причине клетка Пуркинье может возбуждаться по этому входу не более 1-2 раз в 1 с.
Моховидные волокна характеризуются обширной дивергенцией (одно волокно образует синапс ы примерно на 20 клетках-зернах) и оказывают как тормозное, так и возбуждающее действие на клетки Пуркинье. Возбудительные влияния на клетки Пуркинье от моховидных волокон переключаются через клетки-зерна, аксон ы которых поднимаются к поверхности коры мозжечка и, разветвляясь в молекулярном слое, образуют параллельные волокна. Последние оканчиваются возбудительными синапс ами (медиатор - глутамат) на дистальных участках дендрит ов клеток Пуркинье. По этому входу клетка Пуркинье может активироваться в среднем 30-40 раз в 1 с. Важным свойством этого пути является то, что активные клетки-зерна преимущественно активируют клетки Пуркинье, лежащие непосредственно над ними, т.е. эта активность незначительно распространяется по системе параллельных волокон. Отсюда можно сделать вывод о двойственной функции аксон ов клеток-зерен. С одной стороны, их восходящая часть образует радиальную организацию, благодаря которой может осуществляться локальное возбудительное влияние на клетки Пуркинье. Другая функция - модулирующая - связана собственно с параллельными волокнами. Стимуляция мшистых волокон вызывает через аксон ы клеток-зерен кортикальные ВПСП в клетках Пуркинье; они характеризуются градуальностью и последующими ТПСП. Показано, что эти ТПСП генерируются за счет активации тормозных интер нейрон ов в молекулярном слое. Синаптическая активация через параллельные волокна вызывает генерацию натриевых ПД в соме, а при большой амплитуде - генерацию дендрит ных кальциевых ПД.
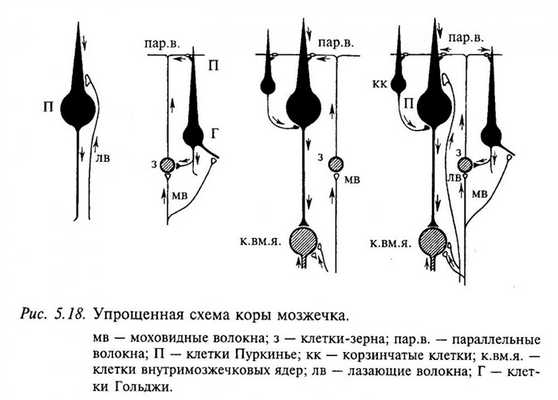
На рис. 5.18 представлена упрощенная морфофункциональная схема коры мозжечка ( нейрон ы, оказывающие тормозное действие, закрашены черным). Видно, что все входы в кору превращаются в тормозные в большинстве случаев через два переключения. Возбуждающее действие оказывают только два типа нейрон ов: клетки-зерна и нейрон ы внутримозжечковых ядер. Оба типа входных волокон (моховидные и лазающие) также являются возбуждающими. Предполагают, что такое большое количество тормозных нейрон ов в коре мозжечка необходимо для предотвращения длительной циркуляции импульсов по нервным цепям. Благодаря этому свойству мозжечок может участвовать в оперативном управлении движениями.
Функция клеток Гольджи состоит в подавлении разряда всех гранулярных клеток, которые слабо возбуждены. За счет этого осуществляется своего рода «фокусирование» ответа на те гранулярные клетки, которые сильно возбуждены через моховидные волокна. Контроль ответа гранулярной клетки осуществляется как за счет отрицательной обратной связи через клетки Гольджи, так и через синаптический вход на когтевидные дендрит ы гранулярной клетки. Как правило, каждый «коготок» дендрит а активируется отдельным моховидным волокном. Чтобы гранулярная клетка возбудилась, необходима суммация по крайней мере двух «коготков». Следовательно, только при «концентрированной» активности в моховидных волокнах можно вызвать разряд в гранулярной клетке. Аксон ы клеток Пуркинье образуют единственный выход из коры мозжечка.
.
Афферентные связи мозжечка делят на следующие группы: восходящие от спинного мозга, вестибулярные - от нижней оливы, ретикулярной формации и ядер моста. В кору мозжечка проецируются также зрительные, слуховые и вегетативные афференты. Все эти пути заканчиваются мшистыми и лазающими волокнами в коре мозжечка. Основная часть этих путей передает информацию о состоянии интер нейрон ного аппарата спинного мозга.
Кора больших полушарий, особенно кора вокруг центральной борозды, образует многочисленные прямые проекции на нейрон ы, залегающие в толще моста (так называемые мостовые ядра). Аксон ы этих нейрон ов проецируются на кору мозжечка. Эти проекции у человека очень обширны, что выражается в образовании валика на вентральной стороне моста (см. приложение 6). Эта система является основным каналом, по которому импульсация от коры больших полушарий достигает коры мозжечка, образуя проекцию как к червю, так и к полушариям мозжечка.
Влияние мозжечка на сегментарный аппарат спинного мозга.
Охлаждение передней доли мозжечка вызывает увеличение как разгибательных, так и сгибательных моносинаптических рефлексов вследствие высвобождения альфа-мото нейрон ов спинного мозга из-под тонического тормозного контроля со стороны мозжечка. Альфа-ригидность, появляющаяся после удаления мозжечка, исчезает после перерезки вестибулоспинального тракта. Это говорит о том, что клетки Пуркинье оказывают на сегментарный аппарат спинного мозга тормозное влияние. Действительно, нейрон ы ядра Дейтерса увеличивают частоту спонтанного разряда примерно в 2 раза после удаления мозжечка. Поскольку вестибулоспинальный тракт организован соматотопически, таким же образом организована регуляция мозжечком антигравитационного тонуса. Напротив, активность гамма-мото нейрон ов при охлаждении передней доли снижается. Это, в свою очередь, снижает частоту фонового разряда первичных и вторичных окончаний мышечных афферентов, а также подавляет их реакции на растяжение мышцы.
На нейрон ах вестибулярных ядер заканчиваются как аксон ы клеток Пуркинье коры червя, так и волокна, берущие начало от ядра шатра. Аксон ы клеток Пуркинье заканчиваются преимущественно на нейрон ах крупноклеточной части латерал ьного вестибулярного ядра (ядро Дейтерса), тогда как аксон ы ядра шатра - на других ядрах вестибулярного комплекса. Через вестибулоспинальные пути осуществляется фазный и тонический контроль экстензорной (антигравитационной) мускулатуры.
Удаление задней части мозжечка (флоккулюса или нодулюса) вызывает у животных дефицит вестибулярного контроля позы и движения. При этом сохраняется ощущение тяжести или движений в пространстве, отсутствуют галлюцинации движения или головокружения. Нарушаются движения глаз: появляется нистагм (обычно горизонтальный с быстрой фазой, направленной к стороне повреждения).
Установлено, что мозжечок модулирует активность нейрон ов пирамидного тракта. Основные пути к коре больших полушарий от мозжечка образованы аксон ами зубчатых и частично промежуточных ядер (через верхние ножки мозжечка). Они переключаются через вентро латерал ьное ядро таламуса, нейрон ы которого проецируются на моторную кору (см. выше). Этот путь организован соматотопически. Другой восходящий путь проходит через ретикулярную формацию ствола, куда поступают волокна от верхних и нижних ножек мозга.
Нарушения позы и движений, вызванные повреждением мозжечка.
Систематические исследования мозжечковых расстройств у животных были начаты итальянским ученым Л. Лючиани в конце XIX века. Его наблюдения были в дальнейшем многократно подтверждены, расширены и уточнены, в том числе клиницистами при изучении симптомов поражения мозжечка у людей.
Удаление мозжечка растормаживает ряд вестибулярных рефлексов и рефлексов, регулируемых сегментарными механизмами спинного мозга. У хронических собак и кошек, у которых удален мозжечок, сразу после операции усиливаются сухожильные рефлексы, гипертонус разгибателей туловища и конечностей (так называемый опистотонус). Примерно через 2 нед после операции собака без мозжечка приобретает способность стоять, и в этот период гипертонус начинает сменяться гипотонией. У обезьян удаление мозжечка сразу вызывает падение мышечного тонуса. Аналогичная картина наблюдается у больных с поврежденным мозжечком. В связи с этим больные с пораженным мозжечком не способны поддерживать определенную позу. Например, если руки больного вытянуты вперед и опираются на подставку, которую внезапно убирают, то руки пассивно падают вниз, в то время как у здорового человека они или сохраняют прежнее положение, или только слегка опускаются. По-видимому, с гипотонусом мускулатуры связано и возникновение мышечной слабости, которая проявляется в быстром наступлении усталости. Примерно в 2 раза уменьшается вес, который животное без мозжечка может нести на спине. Симптомы гипотонии объясняют подавлением активности гамма-мото нейрон ов после удаления мозжечка.
Одно из характерных проявлений мозжечковой недостаточности - это появление тремора, который проявляется в виде колебательных движений конечностей и головы в покое (статический тремор) и во время движения (кинетический, или интенционный, тремор). Обычно тремор более выражен во время движений или при эмоциональном возбуждении. При расслаблении мускулатуры, особенно во время сна, он исчезает. Тремор, который проявляется только во время мышечной активности, связывают с деятельностью гамма-системы. Кинетический тремор наиболее ярко выражен в начале движения и при его завершении, а также при перемене направления движения.
Под атаксией понимают нарушение координации, порядка (лат. taxia - порядок) движений. Наиболее сильно атаксия проявляется у приматов и особенно у человека. Принято различать статическую атаксию - нарушение равновесия при стоянии и динамическую атаксию -нарушение координации двигательных навыков. Больной с пораженным мозжечком не способен быстро сменить одно движение другим, например, барабанить пальцами.
Дисметрия (нарушение размерности движения) проявляется при совершении целенаправленных движений, когда конечность либо не достигает цели (гипометрия), либо проносится мимо нее (гиперметрия). Человек, страдающий дисметрией, не способен выполнить пальценосовую пробу (с закрытыми глазами дотронуться указательным пальцем до собственного носа). У обезьяны, обученной передвигать рычаг между двумя ограничителями, после холодового выключения зубчатого ядра резко ухудшается выполнение этого навыка: рычаг в конце каждого движения упирается в ограничитель, и проходит некоторое время, прежде чем животное начинает передвигать его в противоположном направлении (дисметрия).
Установлено, что поражение полушарных структур (кора полушарий и зубчатое ядро) приводит к атаксии конечностей, тогда как поражение червя, включая ядра шатра, вызывает атаксию туловища. Мозжечковая атаксия хорошо изучена у животных - кошек, собак, обезьян. Так, у «безмозжечковых» собак и кошек лапы широко расставлены, иногда перекрещиваются, при этом животное теряет равновесие и падает. При ходьбе или беге на тредбане амплитуда движений у них сильно варьирует от шага к шагу.
Под асинергией понимают отсутствие дополнительных содружественных движений при выполнении данного двигательного акта. Например, при попытке ходить больной заносит ногу вперед, не переместив центра тяжести, и это приводит к падению назад; при попытке сесть без помощи рук из положения, лежа изолированно сокращаются сгибатели бедра, ноги поднимаются вверх, и больной не может подняться. Больной не способен подняться со стула без помощи рук. Здоровый человек это делает в следующей последовательности: отклоняет туловище вперед, перенося тем самым центр тяжести в площадь будущей опоры, и затем встает. В результате проявления асинергии движение как бы распадается на ряд выполняемых последовательно простых движений. Так, обезьяна, у которой холодовой блокадой выключено зубчатое ядро, выполняет движение рукой, чтобы нажать кнопку, в виде последовательной серии коротких движений. Больной с поражением мозжечка, если рука поднята вверх, а его просят дотронуться до кончика носа, сначала опускает руку, затем сгибает в локте и только после этого подносит палец к носу.
Удаление флоккулонодулярной доли мозжечка у приматов, включая человека, вызывает комплекс расстройств движений глаз: гиперметрию саккад, ухудшение плавных прослеживающих движений глаз, неспособность удерживать взор в эксцентричном положении, осцилляции (тремор) глазных яблок. Из этого следует, что флоккулонодулярная доля участвует в контроле мускулатуры глаз, конечностей и туловища в ситуациях, в которых используется вестибулярный аппарат. Передняя доля мозжечка участвует в контроле локомоции, а полушария - в произвольн ых движениях. Передняя доля получает сигналы от тех областей переднего мозга, в которых формируются двигательные программы. Сюда поступает также информация от головы, шеи, туловища и конечностей. Несмотря на то что мозжечок не имеет своей собственной двигательной системы, он участвует в коррекции движений всех частей тела.
Источник: Шульговский В. В. Основы нейрофизиологии
Дата создания: 13.03.2008
Последнее редактирование: 27.03.2017
Мозжечок
Мозжечок расположен позади больших полушарий мозга над продолговатым мозгом и мостом мозга и представляет собой очень древнюю структуру, выполняющую функцию координации и регуляции произвольных и непроизвольных движений, их коррекции и программирования. Мозжечок принимает участие в двигательной адаптации и двигательном научении, в осуществлении вегетативных функций и некоторых поведенческих актов.
Анатомически мозжечок состоит из средней части - червя, расположенных по обе стороны от него полушарий и боковых флоккулонодулярных долей. Кроме того, мозжечок делят на архицеребеллум (древний мозжечок), к нему относится флоккуломедулярная доля, связанная с вестибулярной системой, и палеоцеребеллум (старый мозжечок), в него входят участки червя мозжечка, пирамида, язык, парафлоккулярный отдел. Он получает информацию в основном от проприоцептивных систем мышц, сухожилий, надкостницы, оболочек суставов. Неоцеребеллум (новый мозжечок) состоит из коры полушарий, мозжечка и участков червя, получает информацию от коры, зрительных и слуховых сенсорных систем.
Кора мозжечка состоит из трех слоев. Верхний слой - молекулярный, в него входят дендритные разветвления грушевидных клеток (клеток Пуркинье), параллельные волокна - аксоны вставочных нейронов. В нижней части молекулярного слоя находятся тела корзинчатых клеток, аксоны которых контактируют с помощью синапсов с телами клеток Пуркинье. Там же находятся и звездчатые клетки. Второй слой - ганглиозный - содержит тела клеток Пуркинье, имеющих самую мощную дендритную систему с большим количеством синапсов (до 60 тысяч). Третий, гранулярный слой состоит из тел вставочных нейронов (гранулярных клеток или клеток-зерен). Аксоны этих клеток образуют в молекулярном слое Т-образные разветвления. В гранулярном слое находятся также клетки Гольджи, аксоны которых направляются в молекулярный слой.
Аксоны клеток Пуркинье передают информацию в подкорковые структуры мозга и ядра мозжечка: ядра шатра, пробковидное, шаровидное и зубчатое ядра. В кору мозжечка поступают два типа афферентных волокон - лазающие и мшистые (ИЛИ моховидные). Лазающие волокна - это аксоны нейронов нижних олив продолговатого мозга, через которые проходят спиномозжечковые тракты (дорсальный и вентральный), передающие информацию от рецепторов кожи, мышц, суставов. Лазающие волокна контактируют с дендритами клеток Пуркинье.
Мшистые волокна - это афферентные пути от ядер моста в мозжечок, они заканчиваются на клетках-зернах коры мозжечка. С помощью афферентных адренергических волокон мозжечок связан с голубым пятном среднего мозга. В окончаниях этих волокон выделяется норадреналин, который, попадая в кору мозжечка, изменяет возбудимость его клеток.
Аксоны гранулярных клеток тормозят клетки Пуркинье и клетки-зерна своего же слоя. Клетки Пуркинье являются только тормозными. Они тормозят клетки ядер мозжечка, регулирующих тонус двигательных центров промежуточного, среднего, продолговатого и спинного мозга.
Ядра мозжечка имеют морфофункциональные связи с различными структурами мозга. Медиальные части коры мозжечка проецируются к ядрам шатра и являются вестибулярными регуляторами. Последние имеют связи с ядром Дейтерса и ретикулярной формацией продолговатого и среднего мозга и через ретикулоспинальный тракт - с мотонейронами спинного мозга.
Промежуточные части коры мозжечка отвечают за координацию позы, целенаправленное движение и коррекцию выполнения медленных движений на основе механизма обратной связи с пробковидным и шаровидным ядрами. Последние влияют на деятельность красного ядра и ретикулярной формации, проецируясь на таламус и двигательную кору.
От нейронов латеральной части коры мозжечка информация поступает в зубчатое ядро, затем через таламус в двигательную кору, а оттуда - к мотонейронам спинного мозга.
Латеральные части коры мозжечка отвечают за программирование сложных движений без использования механизмов обратной связи.
Из мозжечка информация поступает по трем парам ножек. Через верхние - к таламусу, варолиеву мосту, к красному ядру, ядру ствола мозга, в ретикулярную формацию среднего мозга. Через нижние ножки мозжечка сигналы идут к вестибулярным ядрам продолговатого мозга, оливам. Средние ножки связывают неоцеребеллум с лобными долями коры больших полушарий.
ЛЕКЦИЯ № 6. Мозжечок. Строение, функции. Расстройства координации движений
ЛЕКЦИЯ № 6. Мозжечок. Строение, функции. Расстройства координации движений Мозжечок является центром координации движения. Он расположен в задней черепной ямке вместе со стволом мозга. Крышей задней черепной ямки служит намет мозжечка. Мозжечок имеет три пары ножек.Эти
Мозжечок: что мы можем потерять вместе с ним?
Мозжечок: что мы можем потерять вместе с ним? Состоит мозг из белого вещества и серого — это знают все. И одно и второе является нервной тканью. Только белое вещество образуется в основном нейронами, проводящими сигнал в одном направлении, а серое вещество состоит из
1.4.4. Мозжечок
1.4.4. Мозжечок Мозжечок (cerebellum), или малый мозг, расположен в задней черепной ямке, кзади от продолговатого мозга и моста, под затылочными долями полушарий большого мозга. От него мозжечок отделяется твердой оболочкой головного мозга - наметом мозжечка (tentorium cerebelli).
Читайте также:
- Синдром Олбрайта-Баттлера-Блумберг (Albright-Buttler-Bloomberg)
- Артериальная гипотензия: причины, симптомы и лечение
- Сдавление головного мозга. Острые травматические гематомы.
- Разрыв прямой кишки, влагалища при половом акте. Травмы прямой кишки, влагалища при половых сношениях.
- Пример послеоперационной пневмонии. Отек легких