Вегетативные рефлексы вестибулярного аппарата. Сенсорные реакции вестибулярной системы
Добавил пользователь Дмитрий К. Обновлено: 21.01.2026
Раздражение вестибулярного аппарата
Одним из наиболее характерных эффектов раздражения вестибулярного аппарата в результате вращения тела является глазной нистагм. Он состоит в том, что глаза совершают ритмические движения; при каждом из них глаза сначала медленно движутся в сторону, противоположную направлению вращения, а затем очень быстро — в сторону, совпадающую с этим направлением. Иногда вращение тела вызывает также качательные толчкообразные движения головы — нистагм головы .
В возникновении вращательного нистагма основное значение имеет раздражение полукружных каналов, в которых происходит смещение эндолимфы при вращении; некоторое значение имеет при этом и отолитовый аппарат, раздражаемый отклонением головы во время вращения.
При раздражении вестибулярного аппарата при движениях тела или головы происходит перераспределение тонуса скелетной мускулатуры и возникают тонические рефлексы. Большинство этих реакций объясняется функцией отолитового аппарата. Раздражение рецепторных клеток отолитового аппарата вызывает тонические рефлекторные движения шеи, туловища и конечностей.
Кроме всех перечисленных рефлекторных реакций скелетной мускулатуры (вестибуло-моторные рефлексы), раздражение вестибулярных рецепторов вызывает, в особенности при повышенной возбудимости этих рецепторов, характерное ощущение головокружения (вестибуло-сенсорная реакция). Это ощущение представляет собой своеобразное нарушение ориентировки во внешнем мире, сопровождающееся ощущением вращения окружающих предметов.
У животных и у человека при раздражении вестибулярного аппарата возникают также рефлекторные реакции органов, иннервируемых вегетативной нервной системой. Это происходит потому, что импульсы, возникающие в рецепторах вестибулярного аппарата, распространяются на вегетативные нервные центры, вызывая при этом комплекс вестибуло-ативных рефлексов. Последние проявляются в изменениях сердечного ритма, в сужении или расширении сосудов, падении артериального давления, усилении движений желудка и кишечника, рвоте и т. п.
У здоровых людей вестибуло-вегетативные рефлексы возникают при большей силе раздражения, чем вестибуло-моторные, в частности, глазо-двигательные рефлексы. Это связано с тем, что возбудимость центров ативных рефлексов ниже, чем центров моторных рефлексов. При некоторых заболеваниях вестибуло-вегетативные рефлексы проявляются при малой силе раздражения вестибулярного аппарата. Подобно другим реакциям, возникающим при раздражении вестибулярного аппарата, вестибуло-вегетативные рефлексы при вращательном прямолинейном движении возникают лишь при его ускорении или замедлении; при равномерном движении они отсутствуют. Доказательством связи этих рефлексов с раздражением вестибулярного аппарата является исчезновение их после разрушения лабиринтов.
Наличие у некоторых людей резко выраженных вестибуло-вегетативных рефлексов может стать для них препятствием для службы в морском флоте, авиации, на транспорте. Однако возможно несколько повысить выносливость к раздражениям вестибулярного аппарата постепенной тренировкой людей путем многократного их вращения на вращающемся кресле или раскачивания на специальных качелях. Такая тренировка дает возможность подготовить летчиков из людей, вначале непригодных для летного дела вследствие высокой возбудимости вестибулярного аппарата.
Морская болезнь . При повышенной возбудимости вестибулярного аппаратам и связанных с ним нервных центров легко возникает во время пребывании на пароходе в условиях морской качки патологическое состояние, носящее название морской болезни. Оно характеризуется следующими симптомами: бледнеет лицо, выступает хаолодный пот на лбу, возникает ощущение головокружения и тошноты, затем развиваются расстройства равновесия, появляется слюнотечение, дыхание учащается, артериальное давление падает, сердечные сокращения сначала учащаются, затем урежаются возникает рвота. В тяжелых случаях развивается состояние общего угнетения центральной нервной системы. Основное значение в симптомокомплексе морской болезни имеют вестибуло-вегетативные и вестибуло-сенсорные реакции. Главная роль в их возникновении принадлежит рецепторам вестибулярного аппарата; некоторое значение имеют также проириорецеиция и зрительные ощущения (в частности, те нарушения зрения, которые возникают в результате нистагма).
Патологические состояния, аналогичные морской болезни, наблюдаются при полетах (летная болезнь), при поездках в автомобиле и поезде.
ВЕСТИБУЛЯРНЫЕ РЕАКЦИИ
ВЕСТИБУЛЯРНЫЕ РЕАКЦИИ — реакции организма, возникающие в ответ на адекватные (инерционно-гравитационные силы) и не адекватные (температура, электрический ток) раздражения вестибулярного анализатора.
Благодаря наличию двух систем рецепторных образований вестибулярного анализатора — отолитовой и купуло-эндолимфатической (полукружные каналы лабиринта внутреннего уха) — происходит дифференцированное восприятие сил, порождающих прямолинейные, включая силу тяжести, и криволинейные ускорения, осуществляются так наз. лабиринтные реакции. Пороги чувствительности рецепторов отолитовой системы исчисляются в 0,001 — 0,03 g, рецепторов полукружных каналов— в пределах 0,12—2 град/сек 2 при длительности воздействия 0,15 сек. Исключение представляют ускорения Кориолиса (см. Кориолиса ускорение), которые, несмотря на классификацию их как прямолинейных ускорений, имеют низкие пороги раздражения вестибулорецепторов: 0,006—0,06 м/сек 2 . Есть основание полагать, что по мере возрастания силы раздражения дифференциация между двумя рецепторными системами лабиринта становится менее закономерной. Обширные нервные связи вестибулярного анализатора (см.) с корковыми и подкорковыми представительствами различных видов чувствительности, а также с центрами вегетативной нервной системы обеспечивают многообразие проявлений вестибулярных реакций. Выделяют 3 типа вестибулярных реакций: соматические, сенсорные и вегетативные. Первые являются основными, эволюционно закрепленными и определяющими главную функцию вестибулярного анализатора по сохранению пространственного положения. К ним относятся спинальные рефлексы поперечнополосатой мускулатуры, рефлексы более высоких уровней ц. н. с., направленные на сохранение тонуса мышц и равновесия, нормализующие позу и обеспечивающие координацию движений. К соматическим рефлексам относят и вестибулярный нистагм (см.).
Сенсорные (корковые) реакции возникают при определенной интенсивности раздражений и выражаются чувством головокружения, разнообразными иллюзорными ощущениями собственного пространственного положения или окружающих предметов относительно друг друга. Они корригируются представлениями об условнорефлекторной зрительной вертикали.
Вестибуло-вегетативные реакции представляют комплекс разнообразных по своим функциональным проявлениям рефлексов, в осуществлении которых принимает участие вегетативная нервная система.
В нормальных условиях вестибуло-вегетативные реакции обеспечивают необходимый в данный момент уровень обменных процессов в мышечной системе. Доказаны регулирующие воздействия вестибулярного анализатора на состояние внутренней среды организма (в частности, на систему свертываемости крови).
Однако при длительном воздействии вестибулярных раздражений, особенно знакопеременных ускорений, возможен срыв адаптационных механизмов вегетативной нервной системы с возникновением патологических реакций в виде симптомокомплекса болезни передвижения (кинетозы, укачивание, морская, воздушная, космическая, транспортная болезни). Проявления: состояние дискомфорта, нарушение гемоциркуляции, преимущественно в сосудах головного мозга (ранние проявления), нарушение сердечной деятельности (уменьшение ударного объема сердца), дыхания, появление потливости, побледнения, тошноты, рвоты, вплоть до коллапса (см. Вестибулярный симптомокомплекс, Укачивание).
Патогенетическую основу болезни передвижения при знакопеременных ускорениях составляют нарушения функциональных взаимоотношений структур, образующих вестибулярный анализатор, с другими образованиями ц. н. с., в том числе и теми, которые ответственны за регуляцию вегетативных функций.
Библиография: Григорьев Ю. Г., Фарбeр Ю. В. и ВолоховаН. А. Вестибулярные реакции (Методы исследования и влияния различных факторов внешней среды), М., 1970, библиогр.; М о г e н д о-в и ч М. Р. Рефлекторное взаимодействие локомоторной и висцеральной систем, Л., 1957, библиогр.; Разумев в А. Н. и Шипов А. А. Нервные механизмы вестибулярных реакций, М., 1969, библиогр.; X и л о в К. Л. Функция органа равновесия и болезнь передвижения, Л., 1969, библиогр.
ВЕСТИБУЛЯРНЫЙ АНАЛИЗАТОР
Вестибулярный анализатор (статокинетический) — нейродинамическая система, осуществляющая восприятие и анализ информации о положении и движении тела в пространстве. Вестибулярный анализатор состоит из рецепторов, проводящих путей (афферентных и эфферентных), промежуточных центров и коркового отдела. Поступающие в ц. н. с. импульсы от вестибуло- и проприорецепторов обусловливают возникновение вестибуло-моторных (тонических), вестибуло-сенсорных и вестибуло-вегетативных рефлексов.
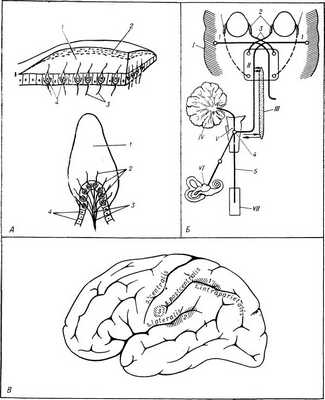
Рис. Структурно-функциональная организация вестибулярного анализатора. А (верхний рисунок) — схематическое строение чувствительного пятна мешочка (utriculus или sacculus): 1 — желатинозное вещество, в к-рое погружены волосковые клетки (4); 2 — слой отолитов; 3 — нервные волокна; А (нижний рисунок) — схематическое строение гребешка (crista ampullaris): 1 — желатинозное вещество (cupula); 2 — волосковые клетки; 3— нервные волокна; 4 — слой опорных клеток. Б — схема внутрицентральных взаимоотношений вестибулярного анализатора: I — кора больших полушарий; II — уровень среднего мозга; III — ретикулярная формация ствола; IV — мозжечок; V — уровень продолговатого мозга; VI — рецептор вестибулярного анализатора; VII — уровень спинного мозга. 1 — m. rectus lat.; 2 — m. rectus med.; 3 — n. abducens; 4 — nuci, vestibulares; 5 — tractus vestibulospinalis. B — проекция вестибулярного анализатора в коре больших полушарий мозга человека (заштриховано): 1 — по Ферстеру; 2 — по Пенфилду; 3 — по Корнхуберу.
Рецепторы Вестибулярного анализатора расположены в перепончатом лабиринте внутреннего уха (см.). Лабиринт представлен тремя полукружными каналами и двумя мешочками — utriculus и sacculus. Каждый канал выходит из utriculus и, описав две трети круга, вновь в него впадает. На месте впадения имеется расширение — ампула, а внутри последней — гребешок (crista ampullris), поверхность к-рого образована слоем волосковых (сенсорноэпителиальных) клеток; одни из них имеют форму кувшина и локализуются на верхушках крист, другие — цилиндрической формы, расположены в склонах; они более организованны и реагируют, видимо, на более локальные раздражения. Те и другие клетки несут на свободном конце сенсорные волокна, каждое из которых представляет собой пучок, состоящий из 70— 80 неподвижных стереоцилий и одной длинной подвижной киноцилии на периферии. Гребешок (рис., А, нижний) покрыт студенистой массой (cupula), примыкающей вплотную к стенке ампулы, в силу чего под влиянием сдвигов эндолимфы происходит ее смещение. Рецепторы мешочков (macula) также состоят из волосковых клеток двух типов и слоя опорных клеток. Над сенсорным эпителием лежит желатиноподобная мембрана, в к-рую вкраплены известковые образования — отолиты (рис., А, верхний). Между отолитами и macula имеется узкое субмембранное пространство, к-рое дает возможность отолитовой мембране скользить по macula и деформировать волоски сенсорных клеток. Macula utriculi имеет форму вогнутого овального пятна размером 3x1 —5 X 2 мм и располагается в основном по дну, по передней и медиальной стенке utriculus. Macula sacculi лежит на медиальной его стенке, имеет овальную форму (0,48 мм 2 ) и образует с macula utriculi угол ок. 90° (см. Отолитовый аппарат).
Естественными раздражителями В. а. являются инерционные сдвиги эндолимфы, возникающие в полукружных каналах при воздействии поворотных и кориолисовых ускорений (см. Ускорение), и перемещение отолитов (преимущественно, видимо, скольжение) относительно волосковых клеток при прямолинейных ускорениях и измененной силе тяжести. Расположение полукружных каналов соответственно плоскостям трехмерного пространства позволяет рецепторам реагировать на инерционные силы, возникающие практически при любых перемещениях тела (активных или пассивных), изменениях величины и вектора гравитации.
От рецепторов лабиринтов начинаются первичные нервные волокна. Первый нейрон проводящей системы находится в вестибулярном ганглии, лежащем в глубине внутреннего слухового прохода и представляющем скопление клеток с двумя отростками. Длинные отростки нейрона составляют вестибулярный нерв (см. Преддверно-улитковый нерв), несущий импульсы ко вторым нейронам — комплексу ядер в мосту, проецирующихся на латеральные углы ромбовидной ямки (area vestibularis).
Вестибулярные волокна оканчиваются в основном на нейронах в центральной части верхнего ядра и латерального ядра, в задне-латеральной части медиального ядра и в задне-медиальной части нижнего преддверного ядра. Часть первичных волокон, не прерываясь в ядрах, направляется в мозжечок. В ядрах берут начало вторичные вестибулярные волокна и направляются в составе медиального продольного пучка к моторным нейронам глазодвигательной мускулатуры и через вестибуло-спинальный тракт до нижних сегментов спинного мозга.
В ядрах ретикулярной формации, по-видимому, проецируются верхние, латеральные и нижние вестибулярные ядра. Обнаружено, что в составе вестибулярного нерва проходят миелиновые волокна, связанные с мозговым стволом и нек-рыми ядрами мозжечка; их основной источник — промежуточное ядро среднего мозга и ядро заднего продольного пучка. От подкорковых образований начинаются волокна нейронов третьего порядка, заканчивающиеся в коре головного мозга (рис., Б).
Локализация коркового отдела В. а. изучена в основном на кошках путем регистрации вызванных потенциалов. Изучение В. а. человека показывает, что корковый отдел В. а. включает поле 21 по К. Бродманну, верхнюю теменную дольку по О. Ферстеру (1936), кору верхней височной извилины по У. Пенфилду (1957) и постцентральную извилину по Корнхуберу (рис., В).
Афферентные пути от вестибулярных рецепторов к коре изучены недостаточно.
Важно, что на нейронах вестибулярных ядер промежуточных центров и в корковой вестибулярной зоне осуществляется конвергенция импульсов, приходящих по различным афферентным волокнам.
Многочисленные афферентные и эфферентные связи Вестибулярного анализатора с различными образованиями ц. н. с. способствуют генерализации эффекта от вестибулярных раздражений. У высокоорганизованных животных и человека пространственный анализ дополняется зрительным, проприоцептивным, тактильным, слуховым анализаторами при главенствующей роли коры головного мозга как интегратора афферентной импульсации. При дефиците сенсорной информации или при чрезмерных вестибулярных раздражениях возможна вестибуло-сенсорная реакция, выражающаяся в потере пространственной ориентации (возникновение головокружения, иллюзорных ощущений).
При перевозбуждении В. а. возникает комплекс вестибуло-вегетативных расстройств, выражающихся повышением тонуса, гл. обр. парасимпатической нервной системы: урежение пульса, снижение кровяного давления, изменения ЭКГ, сужение зрачка, усиление потоотделения, перистальтики жел.-киш. тракта и др. Существенную роль в происхождении вегетативных расстройств играет ретикулярная формация мозгового ствола, с к-рой вестибулярные ядра имеют тесные структурно-функциональные взаимоотношения. Кроме того, необходимо учитывать тесные связи различных ядер В. а. с внутренними органами (В. С. Райцес, А. М. Дутов, 1971). Вестибуло-вегетативные рефлексы могут составлять основу симптомокомплекса болезни передвижения.
Методы оценки функционального состояния В. а. основаны на анализе реакций, вызванных адекватными дозированными раздражениями. Для этого существуют специальные вращающиеся кресла, центрифуги, установки, имитирующие пониженную опору, лифты. Практикуется также использование неадекватных раздражителей (электрический ток, калорический метод — см. Вестибулометрия). Путем повторных воздействий на В. а. достигается привыкание к раздражителям, или тренировка (см. Вестибулярная тренировка), физиологический смысл к-рой сводится к снижению порогов вестибуло-спинальных рефлексов и торможению сенсорных и вегетативных проявлений. Возможно, что патогенетическую основу привыкания составляет иной характер распространения возбуждения в коре головного мозга и изменение корково-нодкорковых взаимоотношений. В. а. функционирует на основе тесного взаимодействия с другими системами мозга и анализаторами: мозжечком, стрио-паллидарной системой, зрительным анализатором и т. д.
Изучение В. а. приобретает особое значение в практике освоения космоса, когда в результате воздействия невесомости и нарушения определенных видов чувствительности создаются условия, временно меняющие физиологические свойства всех анализаторов, определяющих положение тела в пространстве.
Биофизические механизмы ориентационной информации
Для ориентации по отношению к гравитационному полю Земли животные и человек имеют построенный по одному и тому же принципу рецептор гравитации — датчик положения тела. Он характеризуется наличием отолитового аппарата и cupula в вестибулярном аппарате у позвоночных и человека и статоцистов у беспозвоночных. Отолиты и cupula биофизически являются «пробной массой». Рецепторные клетки непосредственно не воспринимают гравитацию. Но любое отклонение в положении тела (линейное и угловое ускорения) сопровождается смещением пробной массы и появлением сил, которые вызывают возбуждение соответствующих групп рецепторных клеток.
Рецептор гравитации животных и человека всегда функционирует по одной и той же схеме, к-рая включает в себя три основных элемента. Первый — упоминавшаяся пробная масса; второй — отходящие от вершины рецепторных клеток способные к отклонению чувствительные образования (киноцилии и стереоцилии); третий элемент — рецепторная клетка, оценивающая изменения чувствительных образований и передающая в закодированном виде информацию ц. н. с. Благодаря особому расположению чувствительных образований на вершине рецепторных клеток при движении пробной массы одни клетки возбуждаются, другие тормозятся. Структурная и функциональная организация рецептора гравитации позволяет ц. и. с. определять ориентацию и осуществлять установку тела животных и человека.
Нарушение функции вестибулярного анализатора при лучевом воздействии. Чувствительность и реактивность В. а. в условиях острого облучения организма в различных дозах исследованы при использовании адекватного раздражителя (углового ускорения и ускорения Кориолиса). Установлено, что изменения в функции лабиринта отмечаются уже при дозах 50—100 рад. При этих дозах, как правило, в первые часы по-еле облучения повышается возбудимость анализатора. Облучение в больших дозах (500 и более рад) приводит к значительному первоначальному угнетению функции лабиринта, что выражается в снижении возбудимости и реактивности В. а. При действии раздражителя большей силы развиваются парадоксальные и ультрапарадоксальные реакции. Через 5 и более часов после облучения происходит нормализация деятельности анализатора. В условиях хронического облучения организма животных в течение 3—6 лет при суммарной дозе 50 рад/год функция вестибулярного анализатора существенно не нарушается, ответные реакции адекватны величине примененного раздражителя; аналогичные результаты были получены и в клинике.
В экспериментах с локальным облучением живота, лицевой части головы или только головного мозга было показано, что в возникновении функциональных нарушений со стороны В. а. большую роль играют так наз. отраженные эффекты. Облучение периферических частей тела также приводит к опосредованному действию ионизирующей радиации на В. а., что выражается в угнетении его деятельности.
Используя рекомендуемые средства профилактики лучевой болезни (см. Радиопротекторы), можно ослабить функциональные нарушения со стороны Вестибулярного анализатора.
На основании проведенных клинико-экспериментальных исследований сделан вывод, что при особых условиях комбинированного действия излучения и различных физ. факторов окружающей среды (вибрация, шумы и др.) возможно сопряженное повышение реактивности В. а.
Нарушение функции В. а. при его облучении может наблюдаться при облучении среднего уха в терапевтических целях, у работающих в определенных профессиональных условиях (напр., рентгенологи), а также у космонавтов при длительных космических полетах.
Нарушение функции вестибулярного анализатора при других патологических состояниях — см. Вестибулярный симптомокомплекс, Лабиринтит, Лабиринтопатия, Меньера болезнь, Укачивание.
Библиография: Винников Я. А. Цитологические и молекулярные основы рецепции, Л., 1971, библиогр.; Горгиладзе Г. И. Корковые механизмы вестибулярной функции, Усп. совр, биол., т. 70, в. 1, с. 65, 1970, библиогр.; Григорьев Ю. Г., Фарбер Ю. В. и Волохова Н. А. Вестибулярные реакции (Методы исследования и влияния различных факторов внешней среды), М., 1970, библиогр.; Курашвили А. Е. и Бабияк В. И. Физиологические функции вестибулярной системы, Л., 19 75, библиогр.; Парин В. В. и др. Очерки по космической физиологии, М., 1967, библиогр.; Разумеев А. Н. и Шипов А. А. Нервные механизмы вестибулярных реакций, М., 1969, библиогр.; Рецептор гравитации, эволюция структурной, цитохимической и функциональной организации, под ред. В. Н. Черниговского, Л., 1971, библиогр.; Хилов К. Л. Кора головного мозга в функции вестибулярного анализатора, М.— Л., 1952, библиогр.; он же, Функция органа равновесия и болезнь передвижения, Л., 1969, библиогр.; Циммерман Г. С. Ухо и мозг, М., 19 74; Handbook of sensory physiology, ed. by H. Autrum a. o., v. 1, В. a. o., 1971; Penfield W. Vestibular sensation and the cerebral cortex, Ann. Otol. (St. Louis), v. 66, p. 691, 1957.
Ф. П. Ведяев, М. Д. Емельянов. Я. А. Винников, К. А. Койчев (биофиз.), Ю. Г. Григорьев (рад.)
Физиология вестибулярного анализатора. Механизмы адекватных раздражителей. Исследования Эвальда, его выводы.
Различают функцию полукружных каналов и функцию мешочков преддверия, где находятся рецепторы вестибулярного анализатора.
Вестибулярный аппарат — орган равновесия — регулирует тонус мышц, поддерживает заданное положение тела, доставляет в кору мозга информацию о положении и перемещении тела в пространстве. При раздражении рецепторов вестибулярного аппарата возникают рефлекторные реакции, способствующие сохранению равновесия тела.
Вестибулярный анализатор в комплексе с рядом других анализаторов регистрирует и координирует положение и движение тела в пространстве. Основной функцией вестибулярного аппарата является сигнализация о возникающих изменениях положения тела в пространстве под влиянием угловых или прямолинейных ускорений.
Рецепторы, сигнализирующие о положении туловища в пространстве, участвуют в механизме регуляции тонуса мышц, способствуя правильному соотношению отдельных частей тела. Рефлекс может исходить из отолитового аппарата (раздражителем в этом случае будет сила тяжести) или из ампулярного — при воздействии углового ускорения (при изменении скорости и направления движения).
Было установлено, что адекватным раздражителем ампулярных аппаратов полукружных каналов является угловое ускорение, для отолитового аппарата преддверия — ускорение или замедление прямолинейных движений тела, а также гравитационное поле Земли.
Угловое ускорение, вызывая смещение эндолимфы, влечет за собой смещение подвижных волосков (киноцилий) чувствительного эпителия на ампулярном гребешке. Фазы движения — ускорение и замедление — постоянно возникают при всех поворотах головы человека в обычной жизни. Вследствие того, что масса эндолимфы в полукружном канале обладает текучестью и определенной инерцией покоя, при ускорении движения эндолимфа отстает и смещается по отношению к стенкам канала, которые повторяют ускорение. Трение о стенки канала постепенно разгоняет эндолимфу и, если движение стало равномерным, жидкость приобретает ту же скорость, что и костные стенки канала, следовательно, смещение ее по отношению к стенкам прекращается, киноцилий чувствительных клеток на ампулярном гребешке приходят в состояние покоя, реакция раздражения из этого канала полностью исчезает. Когда происходит остановки (ускорение с обратным знаком), эндолимфа в силу инерции движения и текучести опережает движение стенок канала, поэтому вновь
отклоняет волоски нейроэпителия только в обратную сторону уже по ходу движения, пока не исчерпается трением энергия движения жидкости.
Отклонение киноцилий чувствительных клеток на ампулярном гребешке и есть момент трансформации механической энергии в энергию электрического импульса. Движение эндолимфы к ампуле называется ампулопетальным током, движение ее в обратном направлении к гладкому концу — ампулофугальным. Закономерности возникающих при этом реакций изучены рядом ученых; наиболее полно они сформулированы в положениях Эвальда и под его именем вошли к практику.
Раздражение полукружных каналов вызывает сенсорные, соматические (анимальные) и вегетативные реакции. Сенсорные реакции возникают сразу в начале углового ускорения и выражаются в появлении ощущения головокружения в сторону направления движения эндолимфы, т.е. в сторону, обратную вращению; после остановки движения эндолимфа смещается уже в сторону вращения, так как она продолжает двигаться по инерции; следовательно, после вращения ощущение головокружения будет в сторону бывшего вращения. Эти сенсорные реакции являются первичными; на основе их появления в процессе филогенеза выработались вторичные соматические (анимальные) реакции.
При раздражении полукружных каналов возникают рефлексы на поперечнополосатую мускулатуру (соматические реакции). Различают рефлексы на мышцы глаз, конечностей, туловища, шеи. Рефлекс на мышцы глаз выражается в появлении нистагма — ритмичных колебаний глазных яблок, которые состоят из медленного и быстрого компонентов. Медленный компонент нистагма обусловлен воздействием со стороны ампулярных рецепторов, а быстрый — воздействием коры мозга. Наркоз снимает быстрый компонент; нистагм при этом не возникает, а глазные яблоки на время раздражения лабиринта остаются в отведенном в соответствующую сторону положении. Раздражение горизонтальных каналов вызывает нистагм в горизонтальной плоскости, раздражение передних каналов — во фронтальной плоскости и задних — в сагиттальной плоскости. Одновременное раздражение двух или трех каналов может вызвать наряду с основной реакцией, например нистагм в горизонтальной плоскости, появление нистагма в вертикальной или фронтальной плоскости (горизонтально-ротаторный или горизонтально-вертикальный нистагм).
В соответствии с силой экспериментального или патологического раздражения различают три степени нистагма. Нистагм первой степени регистрируется лишь в одном положении глазных яблок — при отведении их в сторону быстрого компонента нистагма. Исследующий устанавливает палец на расстоянии 70 см от лица обследуемого и просит его фиксировать взгляд на пальце, затем отводит его в сторону, ожидаемого быстрого компонента, но не очень резко, чтобы не вызвать перенапряжения глазных мышц.
После выявления нистагма в этом положении палец (а следовательно, и глазные яблоки) устанавливают прямо; прекращение нистагма в этом положении свидетельствует о том, что имеется первая степень нистагма. Наличие нистагма и в положении взгляда прямо указывает на то, что он второй степени, но может быть |и третьей. Если при отведении глаз в сторону медленного компонента нистагм исчезает, то, следовательно, его относят ко второй степени. Если нистагм сохраняется и при отведении взгляда в сторону медленного компонента — это нистагм третьей степени.
Чтобы определить продолжительность нистагма, фиксируют взгляд исследуемого в сторону быстрого компонента. Время от начала нистагма (сразу после остановки вращения) и до его прекращения характеризует длительность нистагма. В условиях патологии нистагм может появиться без всякого дополнительного раздражения и продолжаться длительно — такой нистагм называют спонтанным.
Таким образом, нистагм характеризуют по плоскости (горизонтальный, фронтальный, вертикальный), по направлению (вправо, влево, вверх, вниз), по силе (три степени), по амплитуде (крупноразмашистый, среднеразмашистый, мелкоразмашистый), по быстроте (быстрый, медленный или живой, вялый), по продолжительности (в секундах).
Следует отметить, что наибольшую информацию о функциональном состоянии лабиринта дает исследование нистагма после экспериментального раздражения рецепторов полукружных каналов.
Анимальные рефлексы при раздражении ампулярных рецепторов проявляются в виде отклонения рук, ног, туловища и головы в сторону направления медленного компонента нистагма (в сторону движения эндолимфы) для горизонтального полукружного канала, для остальных каналов — в обратную сторону. Эта же закономерность отклонения проявится и при ходьбе в момент действия рефлекса. Следовательно, при раздражении ампулярного рецептора возникает афферентная импульсация к двигательным, вегетативным, мозжечковому и корковому центрам, вызывающая безусловные рефлексы, которые возникают с большим постоянством и потому названы «железными законами».
Эвальд в эксперименте наглухо запломбировал гладкий конец полукружного канала у голубя, затем пневматическим приспособлением (поршнем) надавливал на стенку канала и вызывал тем самым движение эндолимфы к ампуле; при снятии этого давления возникало смещение эндолимфы к гладкому концу канала. Такое же управляемое движение эндолимфы можно вызвать, если рядом с пломбой ввести в канал полую иглу и с помощью поршня шприца направлять движение эндолимфы в одну или другую сторону и регистрировать при этом характер возникающих реакций
ТРИ ЗАКОНА ЭВАЛЬДА:
1. Нистагм возникает в плоскости раздражаемого канала.
2. Ампулопетальный ток эдолимфы в сторону ампулы в горизонтальном полукружном канале - более сильный раздражитель, чем ампулофугальный. В вертикальных каналах этот закон обратный.
3. Нистагм направлен в сторону более активного лабиринта.
Методы исследования функции полукружных каналов вестибулярного анализатора— вращательная проба (кресло Барани - 5 оборотов за 10 секунд, в обе стороны), калорическая проба, прессорная (фистульная) проба (надавливание пальцем на козелок - повышение давления в барабанной полости, появление нистагма и головокружения), исследование отклонений конечностей. Эти методы называются экспериментальными; по силе вызываемого раздражения они стоят в следующем .порядке: более грубым является раздражение вращением, несколько мягче раздражает калорическая проба и прессорная проба.
Вестибуло-вегетативные реакции выражаются в изменении частоты и глубины дыхания, частоты и ритма пульса, артериального давления, изменении дермографизма, температуры кожи, потоотделения.
Адекватными раздражителями отолитового аппарата являются земная гравитация и прямолинейное ускорение. Давление отолитов на волоски нейроэпителия в нормальном положении в покое вызывает рефлексы, поддерживающие тонус мышц для удержания заданного положения. При изменении положения головы в пространстве соответственно меняется положение волосков чувствительных клеток и отолитов, однако остается постоянным направление давления (к центру земли) отолитов. При этом волоски нейроэпителия будут испытывать давление в другом, чем прежде, направлении; вместо давления может возникнуть их натяжение, если они окажутся над отолитами. Таким образом, четко регистрируется изменение положения. Прямолинейные ускорения вызывают смещение (давление) отолитов в силу инерции в сторону, обратную направлению ускорения, так как отолиты не закреплены жестко, а находятся в желатиноподобной мембране. Такое давление является адекватным раздражителем, который вызывает анимальные рефлексы, направленные на поддержание тела в равновесии; возникают также сенсорные и вегетативные рефлексы. При прыжке вверх или прямолинейном ускорении вверх (на лифте) в начале движения отолиты будут сильнее давить на волоски в силу своей инерции; при этом возникает гипертонус сгибателей. Сразу после остановки отолиты по инерции продолжат путь и натянут волоски, что вызовет рефлекс противоположного направления — возникает гипертонус разгибателей.
Физиология равновесия.1.1.Функции вестибулярного анализатора .Организация вестибулярного аппарата.
Вестибулярная сенсорная система состоит из следующих отделов:
периферический отдел включает два образования, содержащие механорецепторы вестибулярной системы — преддверие (мешочек и маточка) и полукружные каналы;
корковый отдел представляют четвертые нейроны, часть которых представлена в проекционном (первичном) поле вестибулярной системы в височной области коры, а другая часть — находится в непосредственной близости к пирамидным нейронам моторной области коры и в постцентральной извилине. Точная локализация коркового отдела вестибулярной сенсорной системы у человека в настоящее время не установлена.
Строение и функции вестибулярного анализатора
Вестибулярный анализатор имеет важное значение в регуляции положения тела в пространстве и его движений. Периферический отдел вестибулярного анализатора является частью внутреннего уха и состоит из полукружных каналов, размещенных в трех взаимно перпендикулярных плоскостях, и из статоцистных органов - двух мешочков - овального (маточки) и круглого, который расположен ближе к улитке.
Вестибулярный орган (орган равновесия, орган гравитации) состоит из трех полукружных каналов и преддверия.Полукружные каналы расположены в трех взаимно перпендикулярных плоскостях: верхний—во фронтальной,задний—в сагиттальной инаружный—в горизонтальной. Преддверие состоит из двух мешочков—круглого (саккулюс), расположенного ближе к улитке, иовального (утрикулюс)расположенного ближе к полукружным каналам.
Полукружные каналы своими устьями открываются в преддверие и сообщаются с ним пятью отверстиями (колена двух каналов: верхнего и заднего — соединены вместе). Один конец каждого канала имеет расширение, которое называется ампулой. Все эти структуры состоят из тонких перепонок и образуют перепончатый лабиринт, внутри которого находится эндолимфа.Вокруг перепончатого лабиринта и между ним и костным его футляром имеетсяперилимфа,которая переходит в перилимфу органа слуха. В каждом мешочке преддверия имеются небольшие возвышения, называемые пятнами, а в ампулах полукружных каналов — гребешками. Они состоят из нейроэпителиальных клеток, имеющих на свободной поверхности волоски (реснички), которые разделяются на две группы: тонкие (их много) — стереоцилии и один более толстый и длинный на периферии пучка—киноцилия
Волосковые клеткипредставляют собой рецепторы вестибулярного анализатора иявляются вторичными. Рецепторные клетки преддверия покрыты желеобразной массой, состоящей в основном из мукополисахаридов; благодаря содержанию в ней значительного количества кристаллов карбоната кальция она получила названиеотолитовой мембраны. В ампулах полукружных каналов желеобразная масса не содержит солей кальция и называетсялистовидной мембраной (купула). Волоски рецепторных клеток пронизывают эти мембраны.
Возбуждение волосковых клеток происходит вследствие скольжения мембраны по волоскам, изгибания волосков (стереоцилии) в сторону киноцилий. Это связано с тем, что механическое управление ионными каналами мембраны волоска с помощью микрофиламентов, зависит от направления сгиба волоска:отклонение в одну сторону приводит к открыванию каналов и деполяризации волосковой клетки, а отклонение в противоположном направлении вызывает закрытие каналов и гиперполяризацию рецептора.
В волосковых клетках преддверия и ампулы при их сгибании генерируется рецепторный потенциал, который усиливает выделение ацетилхолина и через синапсы активирует окончания волокон вестибулярного нерва.
проводниковый отдел. К рецепторам подходят периферическиеволокна биполярных нейронов вестибулярного ганглия, расположенного во вутреннем слуховом проходе (первый нейрон). Аксоны этих нейронов в составе вестибулярного нерва направляются квестибулярым ядрам продолговатогомозга (второй нейрон). Вестибулярные ядра продолговатого мозга (верхнее—ядро Бехтерева,медиальное—ядро Швальбе,латеральное—ядро Дейтерсаинижнее—ядро Роллера) получают дополнительную информацию по аф- ферентным нейронам от проприорецепторов; мышц или от суставных сочленений шейного отдела позвоночника. Эти ядра, где расположен второй нейрон вестибулярного анализатора, тесносвязаны с различными отделами центральной нервной системы.Благодаря этому обеспечиваются контроль и управление эффекторными реакциями соматического, вегетативного и сенсорного характера.
третий нейронрасположен вядрах зрительного бугра, откуда возбуждение направляется в кору большого полушария.
Центральный отделвестибулярного анализатора локализуется ввисочной области коры большого мозга, несколько кпереди от слуховой проекционной зоны (21—22-е поля по Бродману, четвертый нейрон).
Нервные волокна, выходящие из вестибулярных ядер, образуют связи с другими отделами центральной нервной важнейшими из них являются следующие: вестибулоспинальные, вестибуловегетативныеивестибулоглазодвигательные. Эти связи обеспечивают контроль и управление различными двигательными реакциями, а также являются основой для рефлексов обеспечивающих равновесие.
а. Вестибулоспинальный тракт, волокна которого в конечном итоге оказывают влияние в основном наγ-мотонейроны мышц-разгибателей, хотя часть волокон оканчивается и наα-мотонейронах.
б. Связи с мотонейронамишейного отдела спинного мозга, входящие главным образом в вестибулоспинальный тракт.
в.Связи с ядрамиглазодвигательного нерва, которые опосредуют движения глаз, вызываемые вестибулярной активностью; эти волокна проходят в составе медиального продольного пучка. Функцию статолитового аппарата можно тестировать, наблюдая глазодвигательные реакции при наклонах головы.
г. Тракты, направляющиеся ввестибулярные ядра противоположной стороны мозга, благодаря которым афферентация с обеих сторон тела может обрабатываться совместно.
д. Связи сретикулярной формацией, посредством которых обеспечивается воздействие на ретикулоспинальный тракт, являющийся еще одним (полисинаптическим) путем кα- иβ-мотонейронам.
е. Тракты, проходящие через таламус впостцентральную извилину коры головного мозга, обеспечивающие сознательную обработку вестибулярной информации и таким образом сознательную ориентацию в пространстве.
ж.Волокна, направляющиеся вгипоталамус, которые в основном участвуют в возникновении кинетозов (укачивания).
з. Связи смозжечком, в особенности сarchicerebellum( вторичные вестибулярные афференты).мозжечок, может получать некоторые первичные вестибулярные афференты (так называемый прямой сенсорный мозжечковый путь). И первичные, и вторичные вестибулярные афференты у млекопитающих оканчиваются в мозжечке мшистыми волокнами на клетках-зернах флокку-лонодулярной доли (относящейся кarchicerebellum) и частично клеткахuvulaиparaflocculus(paleocerebellum). Клетки-зерна оказывают возбуждающее действие на клетки Пуркине в этих областях, а аксоны последних проецируются опять-таки в вестибулярные ядра. Такая цепь осуществляет тонкую регулировку вестибулярных рефлексов. При нарушениях функции мозжечка вследствие различных заболеваний эти рефлексы утрачивают тормозной компонент, что проявляется в возникновении таких симптомов, как усиленный или спонтанно возникающий нистагм, утрата равновесия, проявляющаяся в неустойчивой походке, избыточной амплитуде движений, особенно при ходьбе («петушиный шаг»). Эти симптомы являются частью синдромамозжечковой атаксии.
Читайте также: