Закладка сосудистой системы. Бластогенез млекопитающих
Добавил пользователь Alex Обновлено: 08.01.2026
2) Бластогенез (греч. blastos - росток + генез) - стадия формирования первичных тканей или ростковая стадия развития организма. Эта стадия длится первые 9-16 суток после оплодотворения (приводятся разные данные о сроках бластогенеза), то есть слияния сперматозоида и яйцеклетки (гаплоидных геномов отца и матери), в результате чего образуется клетка с диплоидным набором хромосом - зигота (греч. zygote - соединенная в пару). Зигота делится, в результате возникают бластомеры или стволовые клетки (греч. blastos + meros - клетка, возникающая при делении яйца животных). За 16 суток накапливаются миллионы стволовых клеток - «строительный материал» будущего организма. Стволовые клетки образуют трехслойную структуру с полостью внутри - прообраз тела человека (животного).
В период бластогенеза формируются такие первичные ткани:
- эктодерма (греч. derma - кожа) - наружный слой;
- мезодерма - средний слой;
- эндодерма - внутренний слой бластомеров (стволовых клеток).
Из эктодермы в последующем развиваются нервная система и кожа с ее придатками, из мезодермы - опорно-двигательный аппарат и кроветворные органы, из эндодермы - внутренние органы. Повреждение одного и тем более двух-трех слоев стволовых клеток влечет развитие массивных пороков строения тела и его органов - бластопатию (греч. blastos + pathos - болезнь, страдание). Бластопатия является причиной гибели зародыша, внематочной беременности, пороков развития с нарушением оси зародыша (симметричные, асимметричные и не полностью разделившиеся близнецы, циклопия, сирингомиелия, аплазия почек, головного мозга и т. д.). Бластопатии возникают под влиянием различных тератогенных факторов.
Распространенность бластопатии неизвестна. Полагают, что 70% зигот гибнет вскоре после оплодотворения. Установлено, что частота многоплодных беременностей (двоен и т. д.) по данным УЗИ в 5-10 раз больше, чем регистрируемая при рождении. Считается доказанным, что первичные половые клетки (гоноциты) и возможные их повреждения появляются у женского зародыша на 14-15-й день развития. Эти повреждения приводят к развитию бластопатии в следующем поколении, т. е. у внуков женщины.
Лишь при отдельных бластопатиях организм доживает до рождения, еще реже, в искусственных условиях, некоторое время может существовать после рождения (так, младенец с анэнцефалией в искусственных условиях жил в течение 3 лет). На стадии бластогенеза возникают также мозаичные формы наследственной патологии, когда, к примеру, у части клеток организма недостает одной хромосомы, у другой их части может быть одна лишняя хромосома. В частности, мозаичная форма синдрома Дауна, при которой трисомия-21 наблюдается преимущественно в нейронах головного мозга.
Предполагается, что в период бластогенеза закладывается тип телесной конституции, что определяется различиями развития и функционирования первичных тканей организма. В соответствии с этим различают три типа строения:
- эктоморфное телосложение (преобладание вертикальных пропорций над горизонтальными и относительно большая голова);
- эндоморфное телосложение (округлость пропорций, склонность к полноте);
- мезоморфное телосложение (преимущественное развитие опорно-двигательного аппарата).
Существуют также смешанные, переходные формы телосложения.
Гипотетически, тип телесной конституции зависит от того, в каком из слоев первичной структуры организма накапливается большее число стволовых клеток. Существуют гипотезы Э. Кречмера и У. Шелдона, согласно которым тип телосложения и некоторые врожденные психические свойства (в основном темперамент) тесно связаны или положительным образом коррелируют друг с другом. Так, У. Шелдон различает три типа темперамента: висцеротонию (ее характеризуют любовь, спокойствие и общительность, она соответствует эндоморфному сложению), соматотонию (ей более свойственны предприимчивость, энергичность, спортивность, она соответствует мезоморфному сложению) и церебротонию (ее отличают сдержанность, застенчивость, тревожность, она соответствует эктоморфному сложению).
Закладка сосудистой системы. Бластогенез млекопитающих
Развитие сосудистого эндотелия в раннем периоде эмбриогенеза является одной из важнейших проблем исследования закономерных процессов гистогенеза и регенерации сосудистого эндотелия в различных тканях, органах и органных системах.
В своей работе мы, базируясь на фундаментальных исследованиях отечественных морфологов (3,4,5,7), исходили из концепции мезенхимного происхождения сосудистого эндотелия. Все существующие до сих пор разногласия касаются не мезенхимной теории происхождения эндотелиальных клеток, а возникают в вопросах трактовки происхождения самих мезенхимных клеток. По мнению одних исследователей (6, 9) основным источником мезенхимы служит мезодерма, дающая ангиобласт, детерминированный в сторону образования эндотелия; тогда как трудами Н.Г. Хлопина (5)и его учеников доказывалось, что клетки мезенхимы, дифференцирующиеся в ангиобласт, представляют собой внезародышевую мезодерму. По-мнению И.М. Борисова (1), R Reiss and D.Noden (8) в формировании эндотелиальной выстилки принимает участие пласт клеток, названный ими субваскулярным слоем, который выселяется из дорзального и вентрального краев боковых листков мезодермы.
Несмотря на большое число существующих теорий о происхождении самих мезенхимных клеток, мы солидарны с результатами исследований R.Wagner (9), установившим, что мезенхимные клетки, дифференцируясь в эндотелий, проходят стадию ангиобласта и в последующем формируют ангиобластические или кровяные островки. При этом происходят процессы агрегации и фиксации мезенхимных клеток, сопровождающиеся изменением их поверхности и появлением клеточных тяжей в местах локализации будущих сосудов.
Некоторые авторы (2) в качестве синонимов употребляют понятия «кровяной островок» и «сосудистый зачаток мезодермального происхождения».
Отсутствие в современной эмбриологической номенклатуре соответствующих терминов, обозначающих стадии развития эндотелия в эмбриогенезе и при его дифференциации в растущем организме, приводит к возникновению разнообразных названий, не всегда отражающих суть происходящего процесса или явления.
Учитывая, что вопрос об источниках развития сосудистого эндотелия в эмбриогенезе является одним из самых важных в учении об ангиогенезе, так как его решение приведет к определению тканевой принадлежности эндотелиальной выстилки, в настоящей статье более подробно хотелось бы остановиться на результатах изучения нами первичных сосудов мозга, полученных при изучении зародышей крыс.
Ранние этапы органогенеза характеризуются наличием путей дососудистой циркуляции, только с ростом массы тела эмбриона возрастает потребность организма в питательных веществах, что служит стимулом к развитию сосудистой циркуляции. Поэтому основное внимание мы уделили ранним этапам гисто- и органогенеза крысы. В связи с тем, что эктодерма обладает мощными ангиогенетическими свойствами, мы при введении в пупочные сосуды беременной крысы маркера сосудов- пероксидазы из хрена смогли наблюдать у 3-4 дневных зародышей на всем протяжении эмбриона первичное сосудистое русло, которое формировало мелкоячеистую сеть замкнутых сосудистых формаций, наиболее разветвленных в области головного конца . При электронномикроскопическом исследовании клеток, выстилающих первичные сосуды зародыша мы определили, что по своей организации на протяжении эмбриона могут быть выделены до 5 зон, характеризующихся отличиями в строении эндотелиальной выстилки сосудов плода. Это: 1) эндотелиальная выстилка сосудов хвостовой части; 2) эндотелиальная выстилка переходной зоны от хвостовой к туловищной; 3) внутренняя выстилка сосудов туловищной части; 4) эндотелиальная выстилка переходной зоны от туловищной области к головной; 5) эндотелиальная выстилка сосудов головы. Клетки внутренней выстилки во всех этих зонах характеризуются различным уровнем дифференцированности и зрелости.
Общие признаки изменения строения эндотелиальной выстилки от хвостовой части к голове представлены следующими изменениями: мезенхимные клетки веретеновидной формы соединяющиеся друг с другом в хвостовой части эмбриона, отграничивая первичные сосуды, по мере продвижения к головной части они несколько теряют отростчатость, однако у них так и не появляется более четками дифференциация на зоны в цитоплазме эндотелиоцита. То есть выстилка сосудов области головы по своему строению ближе к строению мезенхимных клеток, окружающих первичные сосуды; чем к эндотелиоцитам сосудовм с непрерывным типом эндотелия, которые формируются на 14-15 сутки эмбрионального развития плода крысы. При этом число межклеточных соединений с пятнами и зонами облитерации значительно продолжает оставаться достаточно высоким, не исчезают и десмосомы.
Изменения, наблюдаемые нами в цитоплазме клеток, вариабельны, но весьма незначительны: вначале появляются единичные микровезикулы, и на протяжении зародыша их число нарастает мало, при этом содержание органелл синтетического аппарата остается во всех зонах сосудистой выстилки плода достаточно высоким, а уменьшения содержания специфических эндотелиальных телец практически нет. В отношении телец Вейбеля - Паладе можно тем не менее проследить, иногда их смещение в артериальный отдел микрососудистого русла головы.
Итак, у 3-4 дневного крысиного эмбриона уже имеется сформированная первичная сосудистая сеть, окутывающая зародыш снаружи. Несмотря на некоторые различия в строении сосудистой выстилки разных отделов туловища, мы склонны отнести их по ультраструктурной организации к эндотелиобластам.
Такое заключение сделано нами на основании следующих критериев: сходной морфологической организации внутренней выстилки сосудов с окружающими их мезенхимными клетками; значительным развитием в их цитоплазме органелл синтетического аппарата, практическим отсутствием в клетках микропиноцитозных везикул, мощным развитием сократительного аппарата, отсутствия деления клеток на зоны по распределению в их цитоплазме органелл, что характерно для специализированных эндотелиоцитов, а также в связи с наличием чередующихся участков истончений клеток внутренней выстилки с утолщенными участками, особенно в зонах контактов соседних клеток с формированием пятен, зон облитерации или даже десмосом.
И тем не менее, до сих пор существует ряд вопросов по гистогенезу сосудистого эндотелия на ранних стадиях эмбрионального развития, ответы на которые возможно получить при комплексном подходе к изучаемой проблеме с использованием современных методов экспериментальной эмбриологии, а именно с проведением иммунного маркирования клеток - предшественников сосудистого эндотелия. Такой вид исследования с использованием в качестве маркеров моноклональных антител к TGFb 1 и TNF-α, PCNA- ядерного антигена пролиферирующих клеток,а также виментина - маркера клеток мезенхимального происхождения нами начат. В дальнейшем для исследования экспрессии цитокинов (TGFb 1 и TNF-α), локализующихся внутриклеточно, чтобы определить процентное соотношение положительно окрашенных клеток, и для определения цитокинов во внеклеточном пространстве мы проводим полуколичественный метод анализа с помощью световой микроскопии.
Филогенез кровеносной системы.
Кровеносная система позвоночных построена по тому же принципу, что и кровеносная система низших хордовых и даже кольчатых червей. Ее основу составляют брюшной и спинной сосуды, соединенные анастомозами в стенках кишки и в стенках тела. Особенно большое сходство существует между строением кровеносной системы бесчерепных и низших позвоночных (рыб). Это является доказательством единого происхождения всего типа хордовых.
У низших хордовых - ланцетник- кровеносная система замкнутая. Один круг кровообращения. Сердце отсутствует и его роль выполняет пульсирующий сосуд - брюшная аорта. В брюшную аорту поступает венозная кровь от органов и направляется в жаберные артерии. От туда уже окисленная (артериальная) кровь поступает в спинную аорту, которая многочисленные ветви всем органам. В органах эти ветви распадаются на капилляры, где кровь становится венозной. Из капилляров кровь собирается в брюшную аорту.
Класс рыбы. Кровеносная система рыб почти полностью повторяет схему кровеносной системы ланцетника. Отличиями прогрессивного характера являются:
1. Появление специального мышечного органа - сердца, стоящего из 2-ух камер: предсердия и желудочка. Сердце рыб содержит венозную кровь, которая поступает от органов по венозным сосудам в предсердие через венозный синус, затем в желудочек и далее по брюшной аорте в жаберные артерии, где окисляется.
2. Жаберные артерии рыб, в отличие от жаберных артерий ланцетника, распадаются на капилляры в жабрах, что увеличивает дыхательную поверхность.
3. Благодаря сокращениям сердца кровь рыб движется по сосудам быстрее, чем у ланцетника, что обеспечивает (вместе с жаберными капиллярами) более высокую скорость обменных процессов.
Класс Амфибии.У представителей этого класса появляется второй (легочный) круг кровообращения. Сердце амфибий состоит из трех камер: двух предсердий и одного желудочка. В сердце поступает кроме венозной и артериальная кровь. От сердца амфибий отходит только один сосуд - артериальный конус, гомологичный брюшной аорте рыб. От него начинается три пары сосудов:
1. Кожно-легочные артерии - несут венозную кровь в легкие и отдают крупную ветвь к коже. В легких венозная ветвь окисляется и возвращается по легочным венам в левое предсердие. Кожно-легочные вены и легочные артерии образуют малый или легочный круг кровообращения.
2. Пара сосудов (самого крупного диаметра), названных дугами аорты огибает сердце слева и справа, а затем соединяется в непарный сосуд - спинную аорту. Она идет по средней линии тела назад, отдавая многочисленные артерии внутренним органам.
3. Пара сосудов - сонные артерии, несущих кровь к головному мозгу.
В этих трех парах сосудов течет разная кровь: в кожно-легочных артериях - венозная, в дугах аорты - смешанная, в сонных артериях - артериальная.
Для дифференциального распределения крови по сосудам в сердце амфибий имеется специальный механизм. В единственный желудочек амфибий поступает артериальная кровь из левого предсердия и венозная - из правого предсердия. Предсердия сокращаются одновременно и оба вида крови попадают в желудочек. Однако полного смешения крови не происходит поскольку:
1. Стенки желудочка имеют многочисленные мышечные выросты — гребни (трабекулы), образующие мягкие пристеночные камеры и препятствую смешиванию крови в камере желудочка. В левой половине желудочка находится артериальная кровь, в правой - венозная, а по середине смешанная.
2. Сокращение желудочка происходит очень быстро, что так же уменьшает смешивание крови в камере желудочка
3. Наличие клапанов закрывающих лёгочные артерии.
Заканчивается большой круг кровообращения 2 полыми венами (задняя полая вена - от задней половины тела и внутренних органов, передняя полая вена - от головы). Полые вены впадают в правое предсердие.
Класс Рептилии.У рептилий отмечаются прогрессивные изменения, направленные на разделение артериальной и венозной крови, связанные как с изменением сердца, так и основных сосудов. Сердце рептилий 3-х камерное, но в желудочке появляется неполная перегородка, которая препятствует смешиванию крови. В момент сокращения желудочка перегородка практически полностью разделяет его на две камеры - правую и левую. У крокодила же 4-х камерное.
У рептилий происходит редукция артериального конуса и те сосуды, которые отходили от него (дуги кожно-легочных и сонных артерий) теперь отходят от желудочка самостоятельно. У рептилий в отличие от амфибий от сердца отходит не один сосуд, а три, каждый из них берет начало от определенного отдела желудочка и выносит неодинаковую по составу кровь. От левой половины желудочка отходит правая дугааорты, несущая артериальную кровь (она огибает сердце с правой стороны). От нее отходят сосуды к голове и передним конечностям. От середины желудочка (над перегородкой) начинается левая дуга аорты (огибает сердце слева) - несет смешанную кровь к органам задней части тела. От правой половины желудочка отходит легочная артерия, несущая венозную кровь к легким. Правая и левая дуги аорты соединяются позади сердца и образуют спинную аорту. Кровь в ней смешанная, но отличается более высоким содержанием кислорода по сравнению со смешанной кровью амфибий.
Класс птицы. Прогрессивные изменения у представителей класса заключаются в появлении полной перегородки в желудочке (сердце 4-х камерное), что приводит к полному разделению артериальной и венозной крови. Этому способствует также редукция одной из дуг аорты, а именно левой, несущей у рептилий смешанную кровь. У птиц остается только правая дуга аорты, которая начинается от левого желудочка, огибает сердце справа и переходит в спинную аорту, несущую артериальную кровь. Легочная артерия начинается от правого желудочка и несет венозную кровь.
Класс млекопитающие.Кровеносная система млекопитающих принципиально не отличается от кровеносной системы птиц. Из сердца выходит два кровеносных сосуда - дуга аорты и легочная артерия. Дуга аорты огибает сердце с левой стороны.
Приведенный материал показывает, что эволюция кровеносной системы идет по следующим путям:
1. Увеличение количества камер сердца.
2. Дифференцировка сосудов, отходящих от сердца.
3. Повышение содержания кислорода в крови.
У различных классов позвоночных в эмбриогенезе закладываются гомологичные или жаберные дуги. Схема закладки этих сосудов общая. От сердца отходит непарный сосуд - брюшная аорта и от нее к жаберным перегородкам направляются 6 пар крупных сосудов охватывающих глотку и соединяющихся на спинной ее стороне в спинной аорте.
У зародышей рыб- две пары из образовавшихся жаберных дуг быстро исчезают, т.к. две первые жаберные перегородки (висцеральные дуги), включаются в состав черепа. Оставшиеся 4 пары артериальных сосудов функционируют в качестве жаберных артерий. У двоякодышащих рыб от последней пары жаберных артерий обособляется легочная артерия.
У наземных позвоночных- первые две пары жаберных дуг также рано исчезают, а оставшиеся 4 пары, в связи с переходом к атмосферному дыханию подвергаются характерным преобразованиям.
Так, третья пара жаберных дуг у всех наземных теряет 1 связь со следующей парой артерий и несет кровь только вперед к головному мозгу, превращаясь в сонные артерии (а. согоtis).
Четвертая пара сосудов достигает наибольшего развития и во взрослом организме становится основным сосудом, несущим кровь к органам. В соответствии с этим они получили название «дуги аорты». У амфибий и рептилий оба сосуда четвертой пары развиты одинаково (это две дуги аорты), у птиц развивается правая дуга аорты, а левая редуцируется; у млекопитающих же наоборот.
Пятая пара жаберных дуг у наземных позвоночных редуцируется.
Шестая пара превращается в легочные артерии.
Таким образом, сонные артерии гомологичны III паре жаберных дуг; дуги аорты - IУ паре; легочные артерии - УI паре.
Проток, соединяющий в эмбриональном состоянии сонные артерии и дуги аорты, носит название сонного протока и сохраняется во взрослом состоянии у рептилий.
Онтогенез сердечно-сосудистой системы
Первичная закладка сердечно-сосудистой системы у млекопитающих происходит во вне- зародышевой мезенхиме из так называемой ангиобластической ткани (textus angioblasticus), ко- торая, группируясь, образует кровяные островки (insulae sanguineae). Сначала такие островки появляются в трофобласте, затем в мезодерме желточного мешка, а с переходом к плацентар- ному кровообращению и в мезодерме аллантоиса. Одновременно с образованием кровяных островков в желточном мешке аналогичные островки формируются и в теле эмбриона, что совпадает с закладкой у него первых 4 - 7 сомитов.
В кровяных островках периферически расположенные клетки представлены эндотелио- бластами, а центральные - гемоцитобластами. Первые из них, приобретая плоскую форму, дают начало развитию эндотелиальной выстилки будущих кровеносных сосудов, в то время как другие, постепенно округляясь, становятся первичными ядросодержащими клетками крови. В кровяных островках, которые постепенно приобретают удлиненную форму, между эндотелиобластами и гемоцитобластами образуются щелевидные пространства. Последние, постепенно увеличиваясь в размерах и объединяясь между собой, преобразуются в просветы первичных кровеносных сосудов. Объединение кровяных островков сопровождается соеди- нением и развивающихся сосудов, в результате чего формируются примитивные капиллярные сосудистые сети (rete vasculare). Из сосудистых сетей желточного мешка выходят желточные вены (vv. vitallinae), которые, проникая в тело эмбриона, впадают в венозный синус развиваю- щегося сердца зародыша. С образованием замкнутого круга кровообращения развитие новых кровеносных сосудов и их дифференциация происходят за счет боковых ответвлений от основ- ных магистральных сосудов и благодаря усилению сосудов капиллярных сетей развивающихся висцеральных и соматических органов. Последнее может происходить и у взрослых животных при развитии окольного кровообращения, если по той или иной причине нарушается крово- ток в основных сосудах.
раЗВИтИЕ СЕрдЦа. у млекопитающих сердце закладывается в конце 3-й недели эм- бриональной жизни. Первоначально под краниальным концом тела эмбриона в мезодерме спланхноплевры (mesoderma splanchnicum) происходит обособление ее кардиогенной части (mesoderma cardiogenicum) в виде пластинки, в которой развивается сосудистая трубка с ее пер- вичными эндо-, мио- и эпикардиальным слоями (primordium endo-, myo- et epicardiale). дор- сально от кардиогенной пластинки в спланхноплевре происходит образование щелевидных пространств, которые, объединяясь и увеличиваясь в размерах, дают начало формированию околосердечной полости (рис. 82).
С обособлением тела эмбриона от желточного мешка и образованием первичной кишки, правая и левая сосудистые трубки сближаются и объединяются в единую сердечную трубку, или первичное сердце (cor primordiale). Мезодермальные листки спланхноплевры при своем сближении образуют дорсальную и вентральную сердечные брыжейки (mesocardium dorsale et ventrale), которые вскоре рассасываются с образованием единой околосердечной полости.
таким образом, формирование первичного сердца к концу 4-й недели завершается. В нем различают три последовательно соединенные первичные отделы - венозный синус (primorduim sinus venosi), предсердие (primordium atriale) и внутрисердечный желудочек (primordium ventriculare endocardiale) с примитивной полостью (ventriculus saccularis primitivus). От прими- тивного желудочка отходит артериальный конус, который служит началом двух вентральных аорт (aortae ventrales) с отходящими от них аортальными дугами (arcus aortae IV-VI).
Все отделы первичного сердца, продолжая увеличиваться в длину, получают четко выра- женные границы, а артериальный конус преобразуется в примитивную сердечную луковицу (bulbus cordis primitivus), от которой берет начало артериальный ствол (truncus arteriosus). такое сердце получает название простого трубчатого сердца (cor tubulare simplex). В его стенке разли- чают три примитивные оболочки (endo-, myo- et epicardium primitivum).
Продолжая увеличиваться в длину, трубчатое сердце в околосердечной полости делает S-образный изгиб, что позволило его называть сигмовидным сердцем (cor sigmoideum). Его наибольший изгиб имеет место между примитивным желудочком и предсердием. В результате такого изгиба желудочек по отношению к предсердию стал занимать каудовентральное поло- жение, а сердечная луковица приблизилась к венозному синусу и заняла краниодорсальное
Рисунок 82 - Развитие сердца млекопитающего:
А, Б и В - три последовательные стадии развития трубчатого сердца: 1 - эктодерма, 2 - эндодерма, 3 - первичный сомит, 4 - париетальный и 5 - висцеральный листки мезодермы, 6 - вторичная полость тела, 7 - нервный желоб, 7' - нервная трубка, 8 - хорда, 9 - закладка головной кишки, 9' - головная кишка, 10 - нисходящая аорта (парная), 11 - эндотелиальная закладка сердца (парная), 12 - эпикард, 13 - миокард, 14 - эндокард, 15 - полость сердечной трубки, 16 - дорсальная сердечная брыжейка, 17 - вентральная сер дечная брыжейка (редуцирована), 18 - околосердечная (плевроперикардиальная) полость; Г, Д - стадии преобразования сигмовидного сердца в дефинитивное: 1 - венозный синус, 2 - предсердие, 3 - желудоч ковая петля, 3' - желудочек сердца, 4 - артериальный ствол, 5 - правое и 5' - левое сердечные ушки, 6 - венечная борозда; Е, Ж - стадии формирования предсердножелудочкового клапана: 1 - створка клапана, 2 - полость желудочка сердца, 3 - мышечные перекладины, 4 - сухожильные струны, 5 - миокард
должается в артериальный ствол как аортолегочная перегородка (septum aorticopulmonale). аортолегочная перегородка, проходя по спирали, делит артериальный ствол на два хода,
из которых один становится дугой аорты, а другой легочным стволом. По мере выхода из серд- ца они образуют характерный винтообразный изгиб. Если у своего начала аорта располагается каудально от легочного ствола, то затем, обогнув его с правой стороны, переходит на его кра- ниодорсальную поверхность.
Вентральный край аортолегочной перегородки, продолжаясь в полость желудочка и сра- стаясь с межжелудочковой перегородкой, полностью перекрывает межжелудочковое отверстие и одновременно делит предсердно-желудочковый канал на два (правое и левое) предсердно- желудочковые отверстия. различное происхождение межжелудочковой перегородки обусло- вило ее подразделение на мышечную и перепончатую части (partes muscularis et membranaceae). Благодаря проникновению аортолегочной перегородки в первичную полость желудочка и полному подразделению ее на две камеры, входное отверстие аорты стало располагаться в ле- вом желудочке, а легочного ствола - в правом. Между мышечной частью межжелудочковой перегородки и стенками желудочков сохраняются мышечные перемычки, или мясные пере- кладины (trabeculae carneae). Кроме мышечных перекладин на боковых стенках желудочков и межжелудочковой перегородке имеются еще и сосцевидные мышцы (mm. papillares), на ко- торых закрепляются сухожильные струны предсердно-желудочковых клапанов. Последние развиваются на месте предсердно-желудочковых бугорков, располагающихся в предсердно- желудочковых отверстиях. Одновременно во входных и выходных отверстиях сердца проис- ходит развитие полулунных, или кармашковых, клапанов, препятствующих обратному току крови при сокращениях как предсердий, так и желудочков.
раЗВИтИЕ артЕрИй. артериальные сосуды свое развитие получают несколько позже венозных. Начиная с 3-й недели эмбрионального развития, от артериального конуса отходит короткий артериальный ствол (truncus arteriosus), который делится на правую и левую вен- тральные аорты (aortae ventrales). От каждой из них отходят 6 аортальных дуг, из которых 1-я, 2-я и 5-я пары подвергаются ранней редукции, а остальные получают дальнейшее развитие.
артериальный ствол продольной аортолегочной перегородкой подразделяется на аорталь- ный и легочный стволы. аортальный ствол (truncus aortalis), соединяясь с 4-й левой аортальной дугой, образует дугу дефинитивной аорты, которая затем продолжается в грудную аорту. легоч- ный ствол (truncus pulmonalis), соединяясь с 6-й парой аортальных дуг, образует правую и левую легочные артерии.
3-я аортальная дуга вместе с соответствующей дорсальной аортой с правой и левой сто- роны дают начало внутренним сонным артериям. участки правой и левой вентральных аорт, находящиеся между 3-й и 4-й аортальными дугами, преобразуются в общие сонные артерии. Каудальный участок правой вентральной аорты вместе с 4-й правой аортальной дугой дают на- чало правой подключичной артерии.
В области шеи от правой и левой дорсальных аорт, а в области туловища - от их непар- ного продолжения, отходят межсегментные дорсальные, латеральные и вентральные артерии (aa. intersegmentales dorsales, laterales et ventrales).
дорсальные межсегментные артерии подразделяются на дорсальные и вентральные вет- ви. дорсальные ветви за счет дорсальных анастомозов участвуют в образовании позвоночной артерии, а за счет вентральных анастомозов - реберно-шейного ствола. Вентральные ветви участвуют в образовании подключичных, дорсальных межреберных, поясничных и частично наружных подвздошных артерий.
латеральные межсегментные, или латеральные внутренностные, артерии (aa. splanchnicae laterales) представлены парными каудальной диафрагмальной, надпочечниковой, почечной и половой артериями.
Вентральные межсегментные, или вентральные внутренностные, артерии (aa. splanchnicae ventrales) представлены желточными венами, непарным чревным стволом, краниальной и кау- дальной брыжеечными и парными аллантоисными (пупочными) артериями. Пупочные арте- рии участвуют в образовании внутренних подвздошных и частично наружных подвздошных артерий.
раЗВИтИЕ ВЕН. В первую очередь развиваются внезародышевые вены (vv. extraembryonicae), к которым относятся желточные, аллантоисные (пупочные) вены, до- ставляющие в тело эмбриона, а затем плода необходимые питательные вещества и кислород.
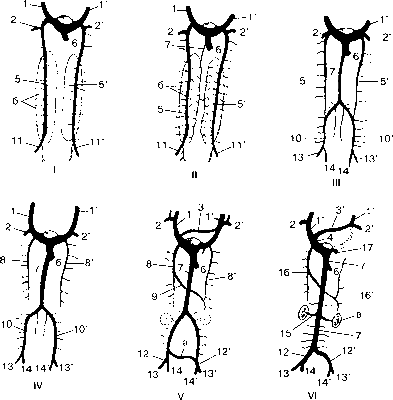
Рисунок 83 - Схема преобразований венозной системы в процессе эмбриогенеза у млекопитающих:
I-VI - последовательные стадии развития вен туловища (вид с вентральной поверхности). 1 - правая и 1' - левая краниальные кардинальные вены; 2, 2' - подключичные вены; 3 - анастомоз от левой краниальной кар динальной вены к одноименной вене с правой стороны; 3' - плечеголовная вена; 4 - краниальная полая вена; 5 - правая и 5' - левая каудальные кардинальные вены; 6 - печеночная вена; 7 - каудальная полая вена; 8, 8' - краниальные части каудальной кардинальной вены; 9 - анастомоз от левой каудальной кардинальной вены к одноименной вене правой стороны; 10, 10' - каудальные части каудальной кардинальной вены; 11, 11' - се далищные вены; 12, 12' - общие подвздошные вены; 13, 13' - наружные и 14, 14' - внутренние подвздошные вены; 15 - почечная вена; 16 - правая и 16' - левая непарные вены; 17 - венечный синус; а - анастомоз между общими подвздошными венами; б - туловищная (промежуточная) почка; в - дефинитивная почка
у эмбриона вместе с развитием сердца происходит и развитие внутризародышевых вен (vv. intraembryonicae). Вначале получают развитие парные (передние и задние) кардинальные вены (vv. cardinales craniales et caudales), отводящие венозную кровь из соответствующих участ- ков тела эмбриона. На уровне каудального конца сердца передняя и задняя кардинальные вены соответствующей стороны тела эмбриона, соединяясь друг с другом, образуют правую и левую общие кардинальные вены (vv. cardinales communes dextra et sinistra), которые вместе с желточными венами впадают в венозный синус сердца. Вскоре между левой и правой передни- ми кардинальными венами развивается крупный предкардинальный анастомоз, преобразую- щийся затем в левую плечеголовную вену (v. brachiocephalica sinistra). участок правой передней кардинальной вены, располагающийся перед анастомозом, становится правой плечеголовной веной. В результате таких преобразований вся венозная кровь, оттекающая от передних участ- ков тела, в сердце поступает по правой общей кардинальной вене, которая затем становится краниальной полой веной (v. cava cranialis). От левой общей кардинальной вены сохраняется лишь небольшая часть, преобразующаяся в венечный синус (sinus coronarius), в который впада- ют коронарные вены, отводящие венозную кровь от самого сердца. у жвачных и свиньи левая общая кардинальная вена частично сохраняется в виде левой непарной вены (v. azygos sinistra). C развитием промежуточной (туловищной) почки происходит образование парных над-
и подкардинальных вен (vv. supra- et subcardinales), которые имеют частые анастомозы как с задними кардинальными венами, так и между собой. С редукцией промежуточной почки зад- ние кардинальные вены подвергаются запустению и от них сохраняются лишь краниальные участки, которые вместе с надкардинальными венами участвуют в образовании непарных (vv. azygos) и большой сердечной вены (v. cordis magna).
Подкардинальные вены на уровне дефинитивной почки соединяются между собой круп- ным анастомозом (anastomosis subcardinalis), в который впадает венозный сосуд от левой дефи- нитивной почки (v. renalis sinistra). В подкардинальные вены впадают вены от надпочечников (vv. adrenales) и половых желез (w. gonodales), а их краниальные участки, соединяясь анастомо- зами с надкардинальными венами, принимают участие в образовании каудальной полой вены (v. cavacaudalis), отводящей венозную кровь от каудальных участков тела.
Надкардинальные вены, имея анастомозы с подкардинальными венами, подвергаются значительной редукции. Сохранившиеся ее передние участки преобразуются в правую и ле- вую непарные вены (vv. azygos dextra et sinistra), в образовании которых принимают участие и передние участки каудальных кардинальных вен.
Венозная кровь, оттекающая от первичной кишки, по брыжеечным (подкишечным) ве- нам направляется в зачаток печеночного синуса, откуда вместе с желточными венами впадает в венозный синус сердца. С развитием печени брыжеечные вены, составляющие группу при- носящих вен (afferentes hepatis), направляются в печень, где, распадаясь на мелкие сосуды, об- разуют ее синусоиды. Из последних выходят выносящие вены печени (vv. efferentes hepatis), или просто печеночные вены (hepaticae), впадающие в каудальную полую вену. Со сменой жел- точного кровообращения на плацентарное желточные вены редуцируются и вены кишечника преобразуются в систему воротной вены печени (v. portae hepatis), становясь частью каудальной полой вены.
С развитием венозных сосудов грудных и тазовых конечностей происходит усиление меж- сегментных вен (vv. intersegmentales).
Филогенез основных систем органов позвоночных животных филогенез кровеносной системы эволюция артериальной системыу позвоночных
Эволюцию артериальной системы у позвоночных можно проследить, наблюдая изменение сосудов в процессе развития зародышей. На ранних стадиях развития спереди от сердца закладывается крупный сосуд - ствол аорты (брюшная аорта), от него метамерно ответвляются парные сосуды - артериальные дуги, охватывающие глотку. Обычно у рыб их возникает 6-7 пар, а у наземных позвоночных - 6 пар. На спинной стороне они впадают в два корня спинной аорты, переходящих в спинную аорту.
По мере развития зародыша разных позвоночных происходит преобразование дуг аорты.

У рыб первые две пары артериальных дуг редуцируются, а четыре пары (3, 4, 5, 6) функционируют как приносящие и выносящие жаберные артерии. У наземных позвоночных редуцируются первая, вторая и пятая пары дуг. Третья пара жаберных дуг превращается в начальную часть сонных артерий.
За счет четвертой пары развиваются главные сосуды большого круга - дуги аорты. У земноводных и пресмыкающихся развиваются две дуги аорты, у птиц - только правая, у млекопитающих - только левая дуга. У хвостатых амфибий и некоторых рептилий сохраняется связь между сонными артериями и дугами аорты в виде сонного протока.
За счет шестой пары артериальных дуг у наземных позвоночных развивается главный сосуд малого круга - легочные артерии. До конца эмбриональной жизни они остаются связанными с аортой боталловым протоком. У хвостатых амфибий и некоторых рептилий боталлов проток сохраняется и во взрослом состоянии. У человека сонный и боталлов протоки редуцируются и могут встречаться только как аномалии развития.
КРОВЕНОСНАЯ СИСТЕМА ЛАНЦЕТНИКА
Кровеносная система ланцетника замкнутая, круг кровообращения один, кровь бесцветная, сердце отсутствует (рис. 2). Его функцию выполняет пульсирующий сосуд - брюшная аорта, расположенный под глоткой. В результате его пульсации венозная кровь из брюшной аорты поступает в многочисленные (100-150 пар) приносящие жаберные артерии.
Через стенки этих артерий, расположенных в перегородках между жаберными щелями, происходит газообмен и, образующаяся при этом, артериальная кровь по дистальным концам жаберных артерий собирается в парные корни аорты, которые, сливаясь, переходят в непарный сосуд - спинную аорту, тянущуюся под хордой назад. От корней аорты к переднему концу тела кровь поступает по сонным артериям.
После газообмена образуется венозная кровь, которая из капилляров тканей собирается в вены. Вены переднего и заднего отделов тела сливаются в парные передние и задние кардинальные вены, которые, соединяясь, образуют правый и левый кювьеровы протоки.
Непарная хвостовая вена переходит в подкишечную вену, которая подходит к печеночному выросту и образуют в нем воротную систему, которая на выходе образуют печеночную вену. Из печеночной вены и кювьеровых протоков кровь поступает в брюшную аорту.
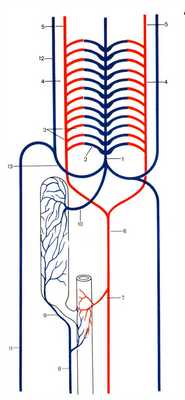
Рисунок 2. Строение кровеносной системы ланцетника. 1. Брюшная аорта 2. Жаберные приносящие артерии 3. Жаберные выносящие артерии 4. Корни спинной аорты 5. Сонные артерии 6. Спинная аорта 7. Кишечная артерия 8. Подкишечная вена 9. Воротная вена печени. 10. Печеночная вена 11. Правая задняя кардинальная вена 12. Правая передняя кардинальная вена 13. Общая кардинальная вена
КРОВЕНОСНАЯ СИСТЕМА РЫБ
Кровеносная система рыб замкнута, круг кровообращения один. Сердце двухкамерное (рис.3), состоит из желудочка и предсердия. К последнему примыкает венозный синус, в который собирается венозная кровь из органов.
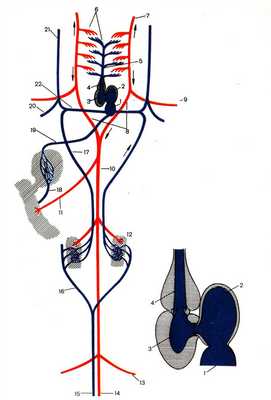
Рисунок 3. Строение кровеносной системы и сердца рыбы. 1. Венозный синус 2. Предсердие 3. Желудочек 4. Луковица аорты 5. Брюшная аорта 6. Жаберные сосуды 7. Левая сонная артерия 8. Корни спины аорты 9. Левая подключичная артерия 10. Спинная аорта 11. Кишечная артерия 12. Почки 13. Левая подвздошная артерия 14. Хвостовая артерия 15. Хвостовая вена 16. Правая воротная вена почек 17. Правая задняя кардинальная вена 18. Воротная вена печени 19. Печеночная вена 20 Правая подключичная вена 21. Правая передняя кардинальная вена 22. Общая кардинальная вена
КРОВЕНОСНАЯ СИСТЕМА ЗЕМНОВОДНЫХ
Кровеносная система земноводных имеет определенные черты прогрессивной организации, что связано с наземным образом жизни и появлением легочного дыхания.
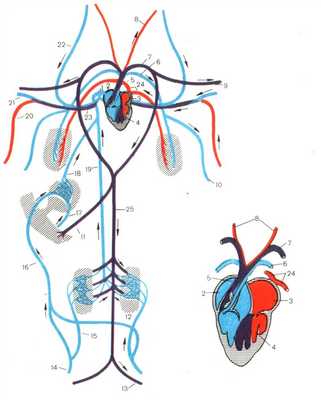
Рисунок 4. Строение кровеносной системы и сердца амфибии 1. Венозный синус 2. Правое предсердие 3. Левое предсердие 4. Желудочек 5. Артериальный конус 6. Левая легочная артерия 7. Левая дуга аорты 8. Сонные артерии 9. Левая подключичная артерия 10. Левая кожная артерия 11. Кишечная артерия 12. Почки 13. Левая подвздошная артерия 14. Правая подвздошная артерия 15. Воротная вена почек 16. Брюшная вена 17. Воротная вена печени 18. Печеночная вена 19. Задняя полая вена 20. Кожная вена 21. Правая подключичная вена 22. Правая яремная вена 23. Передняя полая вена 24. Легочные вены 25. Спинная аорта.
Сердце трехкамерное (рис. 4), состоит из двух предсердий, желудочка, венозного синуса и артериального конуса. Имеются два круга кровообращения, однако артериальная и венозная кровь частично смешиваются. Из желудочка кровь выходит одним потоком через артериальный конус, от которого берет начало брюшная аорта, разделяющаяся на три пары больших сосудов:
1) кожно-легочные артерии,
3) сонные артерии.
Но состав крови в этих сосудах различен, что обусловлено следующими особенностями сердца:
а) наличие в желудочке на задней стенке мышечных тяжей (трабекул), образующих многочисленные карманы;
б) отхождение артериального конуса от правой половины желудочка сзади;
в) наличие в артериальном конусе спирального лопастевидного клапана, перемещающегося в связи с сокращением стенок артериального конуса.
Во время систолы предсердий в желудочек поступает артериальная кровь из левого предсердия и венозная - из правого. В мышечных карманах часть крови задерживается, а смешивается лишь в середине желудочка. Поэтому во время диастолы (расслабления) желудочка в нем находится кровь разного состава: артериальная, смешанная и венозная.
При сокращении (систоле) желудочка в артериальный конус устремляется прежде всего венозная кровь из правых карманов желудочка. Она попадает в кожно-легочные артерии. При дальнейшем сокращении желудочка в артериальный конус поступает следующая, наибольшая по объему порция крови из средней части желудочка - смешанная. В связи с нарастанием давления в артериальном конусе спиральный клапан отклоняется влево и закрывает собой отверстие легочных артерий. Поэтому смешанная кровь поступает в следующую пару сосудов - дуги аорты. Наконец, на высоте систолы желудочка в артериальный конус поступает артериальная кровь из наиболее отдаленного от него участка - из левых карманов желудочка. Эта артериальная кровь направляется в незаполненную еще последнюю пару сосудов - в сонные артерии.
Кожно-легочное артерии недалеко от легких разветвляются на две ветви - легочную и кожную. После газообмена в капиллярах легких и в коже артериальная кровь поступает в вены, направляющиеся к сердцу. Это малый круг кровообращения. Легочные вены впадают в левое предсердие, кожные вены несут артериальную кровь в передние полые вены, впадающие в венозный синус. Следовательно, в правое предсердие поступает венозная кровь с примесью артериальной.
Дуги аорты, отдав сосуды к органам передней половины тела, соединяются и образуют спинную аорту, дающую сосуды к задней половине тела. Все внутренние органы снабжаются смешанной кровью, за исключением головы, куда поступает артериальная кровь из сонных артерий. Пройдя в капиллярах по органам тела, кровь становится венозной и поступает в сердце. Главнейшими венами большого круга являются: парные передние полые вены и непарная задняя полая вена, впадающие в венозный синус.
КРОВЕНОСНАЯ СИСТЕМА ПРЕСМЫКАЮЩИХСЯ
Кровеносная система рептилий (рис. 5) отличается более высокой организацией:
1. Сердце трехкамерное, но в желудочке имеется неполная перегородка, поэтому артериальная и венозная кровь смешиваются в значительно меньшей степени, чем у амфибий.
2. Артериальный конус отсутствует и артерии отходят от сердца не общим стволом, как у амфибий, а самостоятельно тремя сосудами.
От правой половины желудочка отходит легочная артерия, разделяющаяся на выходе из сердца на правую и левую, несущие венозную кровь. От левой половины желудочка отходит содержащая артериальную кровь правая дуга аорты, от которой ответвляются две сонные артерии, несущие кровь к голове, и две подключичные артерии.
На границе между правой и левой половиной желудочка берет начало левая дуга аорты, она несет смешанную кровь.
Каждая дуга аорты огибает сердце: одна справа, другая слева и соединяются в непарную спинную аорту, которая тянется назад, отсылая ряд крупных артерий к внутренним органам.
Венозная кровь из передней части тела собирается по двум передним полым венам, а из задней части тела по непарной задней полой вене. Полые вены впадают в венозную пазуху, которая сливается с правым предсердием.
В левое предсердие впадают легочные вены, несущие артериальную кровь.
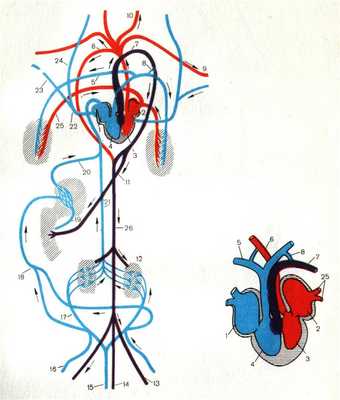
Рисунок 5. Строение кровеносной системы и сердца рептилии. 1.Правое предсердие. 2. Левое предсердие 3. Левая половина желудочка 4. Правая половина желудочка 5. Правая легочная артерия 6. Правая дуга аорты 7. Левая дуга аорты 8. Левый артериальный проток 9. Левая подключичная артерия 10. Левая сонная артерия 11. Кишечная артерия 12. Почки 13. Левая подвздошная артерия 14. Хвостовая артерия 15. Хвостовая вена 16. Правая бедренная вена 17. Правая воротная вена почек 18. Брюшная вена 19. Воротная вена печени 20. Печеночная вена 21. Задняя полая вена 22. Правая передняя полая вена 23. Правая подключичная вена 24. Правая яремная вена 25. Правая легочная вена 26 Спинная аорта
КРОВЕНОСНАЯ СИСТЕМА ПТИЦ
Кровеносная система птиц по сравнению с пресмыкающимися обнаруживает черты прогрессивной организации.
Сердце четырехкамерное, малый круг кровообращения полностью отделен от большого. От желудочков сердца отходят два сосуда. От правого желудочка по легочной артерии венозная кровь поступает в легкие, откуда по легочной вене окислившаяся кровь поступает в левое предсердие.
Сосуды большого круга начинаются от левого желудочка одной правой дугой аорты. Вблизи сердца от дуги аорты отходят правая и левая безымянные артерии. Каждая из них разделяется на сонную, подключичную и грудную артерии соответствующей стороны. Аорта, обогнув сердце, проходит под позвоночником назад. От нее отходят артерии к внутренним органам, задним конечностям и хвосту.
Венозная кровь из передней части тела собирается в парные передние полые вены, а из задней - в непарную заднюю полую вену, эти вены впадают в правое предсердие.
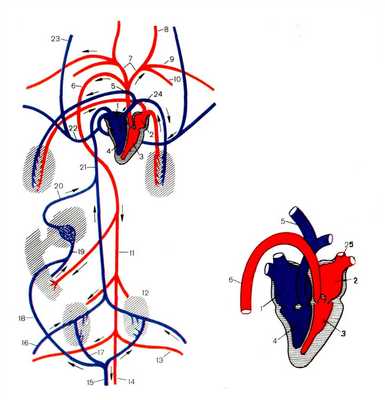
Рисунок 6. Строение кровеносной системы и сердца птицы. 1.Правое предсердие 2. Левое предсердие 3. Левый желудочек 4. Правый желудочек 5. Правая легочная артерия 6. Дуга аорты 7. Безымянная артерия 8. Левая сонная артерия 9. Левая подключичная артерия 10. Левая грудная артерия 11. Спинная аорта 12. Почки 13. Левая подвздошная артерия 14. Хвостовая артерия 15. Хвостовая вена 16. Правая бедренная вена 17. Правая воротная вена почек 18. Ключично-брыжеечная вена 19. Воротная вена печени 20. Печеночная вена 21. Задняя полая вена 22. Правая передняя полая вена 23. Правая яремная вена 24. Правая легочная вена
КРОВЕНОСНАЯ СИСТЕМА МЛЕКОПИТАЮЩИХ
Сердце, как и у птиц, четырехкамерное. Правая половина сердца, содержащая венозную кровь полностью отделена от левой - артериальной.
Малый круг кровообращения начинается от правого желудочка легочной артерией, которая несет венозную кровь к легким. Из легких артериальная кровь собирается в легочные вены, которые впадают в левое предсердие.
Большой круг кровообращения начинается аортой, выходящей из левого желудочка (рис.).
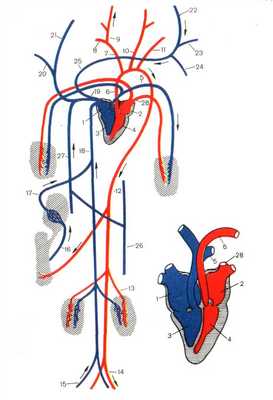
Рисунок 7. Строение кровеносной системы и сердца млекопитающих. 1.Правое предсердие 2. Левое предсердие 3. Правый желудочек 4. Левый желудочек 5. Левая легочная артерия 6. Дуга аорты 7. Безымянная артерия 8. Правая подключичная артерия 9. Правая сонная артерия 10. Левая сонная артерия 11.Левая подключичная артерия 12. Спинная аорте 13. Почечная артерия 14. Левая подвздошная артерия 15. Правая подвздошная вена 16. Воротная вена печени 17. Печеночная вена 18. Задняя полая вена 19. Передняя полая вена 20. Правая подключичная вена 21. Правая яремная вена 22. Левая яремная вена 23. Левая подключичная вена 24. Верхняя межреберная вена 25. Безымянная вена 26. Полунепарная вена 27. Непарная вена 28. Легочные вены
В отличие от птиц аорта млекопитающих огибает сердце слева. От левой дуги аорты отходят три сосуда: короткая безымянная артерия, левая сонная артерия и подключичная. Обогнув сердце, аорта тянется назад вдоль позвоночника, от нее отходят сосуды к внутренним органам.
Венозная кровь собирается в заднюю и переднюю полые вены, которые впадают в правое предсердие.
В эмбриогенезе человека наблюдается ряд филогенетических преобразований сердца (рис. 8), что имеет важное значение для понимания механизмов развития врожденных пороков сердца.
У низших позвоночных (рыбы, амфибии) сердце закладывается под глоткой в виде полой трубки. У высших позвоночных и у человека сердце закладывается в виде двух далеко отстоящих друг от друга трубок. Позже они сближаются, перемещаясь под кишку, а затем смыкаются, образуя единую трубку, расположенную посередине.
У всех позвоночных передняя и задняя части трубки дают начало крупным сосудам. Средняя же часть начинает быстро и неравномерно расти, образую S-образную форму. После этого задняя часть трубки перемещается на спинную сторону и вперед, образуя предсердие. Передняя часть трубки при этом не смещается, стенки ее утолщаются, и она преобразуется в желудочек.
У рыб одно предсердие, а у амфибий оно разделяется растущей перегородкой на два. Желудочек у рыб и амфибий один, но в желудочке последних имеются мышечные выросты (трабекулы), образующие мелкие пристеночные камеры. У рептилий образуется неполная перегородка, растущая снизу вверх, каждое предсердие уже имеет свой выход в желудочек.
У птиц и млекопитающих желудочек разделен на две половины - правый и левый.
В процессе эмбриогенеза у млекопитающих и человека вначале имеется одно предсердие и один желудочек, отделяющиеся друг от друга перехватом с атриовентрикулярным каналом, сообщающим предсердие с желудочком. Затем в предсердии спереди назад начинает расти перегородка, разделяющая предсердие на два. Одновременно с дорсальной и вентральной стороны начинают расти утолщения (атриовентрикулярные подушки). Соединяясь, они разделяют общее атриовентрикулярное отверстие на два отверстия: правое и левое. Позднее в этих отверстиях формируются клапаны.
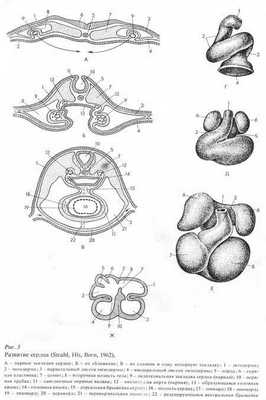
Рисунок 8. Развитие сердца. А - парные закладки сердца, Б - их сближение, В - их слияние в одну непарную закладку: 1- эктодерма; 2 - энтодерма; 3 - париетальный листок мезодермы; 4 - висцеральный листок мезодермы; 5 - хорда; 6 - нервная пластинка; 7 - сомит; 8 - вторичная полость тела; 9 - эндотелиальная закладка сердца; 10 - нервная трубка; 11 - ганглиозные нервные валики; 12 - образующаяся головная кишка; 14 - головная кишка; 15 - дорсальная брыжейка сердца; 16 - полость сердца; 17 - эпикард; 18 - миокард; 19 - эндокард; 20 - перикард; 21 - вертикальная полость; 22 - редуцирующаяся вертикальная брыжейка.
Межжелудочковая перегородка образуется из разных источников: верхняя ее часть возникает за счет клеток атриовентрикулярных подушек, нижняя - за счет гребнеобразного выпячивания дна желудочка, средняя - за счет перегородки общего артериального ствола, который подразделяется на сосуды - аорту и легочный ствол. В месте соединения трех закладок перегородки образуется перепончатая часть, на месте которой формируется межжелудочковая перегородка. Отклонения в развитии межжелудочковой перегородки являются причиной такой врожденной патологии, как отсутствие или недоразвитие ее. Кроме этого, нарушение эмбриогенеза сердца может выражаться в незаращении межпредсердной перегородки, чаще в области овальной ямки (у эмбрионов - отверстие) или внизу, если не произошло ее срастание с атриовентрикулярным кольцом.
Из аномалий развития сосудов наиболее часто встречается незаращение боталлова протока (от 6 до 22%), который функционирует во время внутриутробной жизни, направляя кровь из легких (спавшихся) в аорту. После рождения в норме он зарастает в пределах 10 недель. Если проток сохраняется во взрослом состоянии, у больного повышается давление в малом кругу, развивается застой крови в легких, что приводит к сердечной недостаточности. Реже встречается более тяжелая патология - незаращение сонного протока. Кроме того, может вместо одной дуги аорты развиться две - левая и правая, которые образуют аортальное кольцо вокруг трахеи и пищевода. С возрастом это кольцо может сужаться и нарушается глотание.
На определенной стадии эмбрионального развития от желудочков отходит общий артериальный ствол, который в дальнейшем разделяется спиральной перегородкой на аорту и легочный ствол. Если такая перегородка не формируется, то образуется общий артериальный ствол, в котором смешивается артериальная и венозная кровь. Это приводит к смерти.
Иногда встречается транспозиция аорты, когда она начинается не от левого желудочка, а от правого, а легочная артерия - от левого желудочка, если перегородка общего артериального ствола приобретает не спиральную, а прямую форму.
Серьезной аномалией является развитие в качестве главного сосуда правой артерии четвертой жаберной дуги и правого корня спинной аорты вместо левых. В этом случае дуга аорты берет начало из левого желудочка, но поворачивает вправо. При этом возможно нарушение функции соседних органов.
Читайте также: