
Что входит в состав вирусных частиц входят
Так как вирусы не относятся к клеточной форме жизни, в качестве обозначения дискретной вирусной частицы используется термин "вирион". Это понятие было введено в 1962 г. французом Андре Львовым.
Вирус существует в этом виде не постоянно, а лишь на определенном этапе своего жизненного цикла.
Что такое вирион
Вирион представляет собой конечную фазу развития вируса, включающую полный набор структурных и функциональных элементов, упакованных в единую частицу. Такая форма характерна для внеклеточной стадии жизненного цикла вируса, однако какое-то время после сборки вирион может существовать и внутри инфицированной клетки.
Так как вирион – это всего лишь обозначение морфологической единицы, его не стоит отождествлять с понятием "вирус". Последний включает всю совокупность биологических свойств, характеризующих этот таксон, а не только особенности строения.
Строение вириона
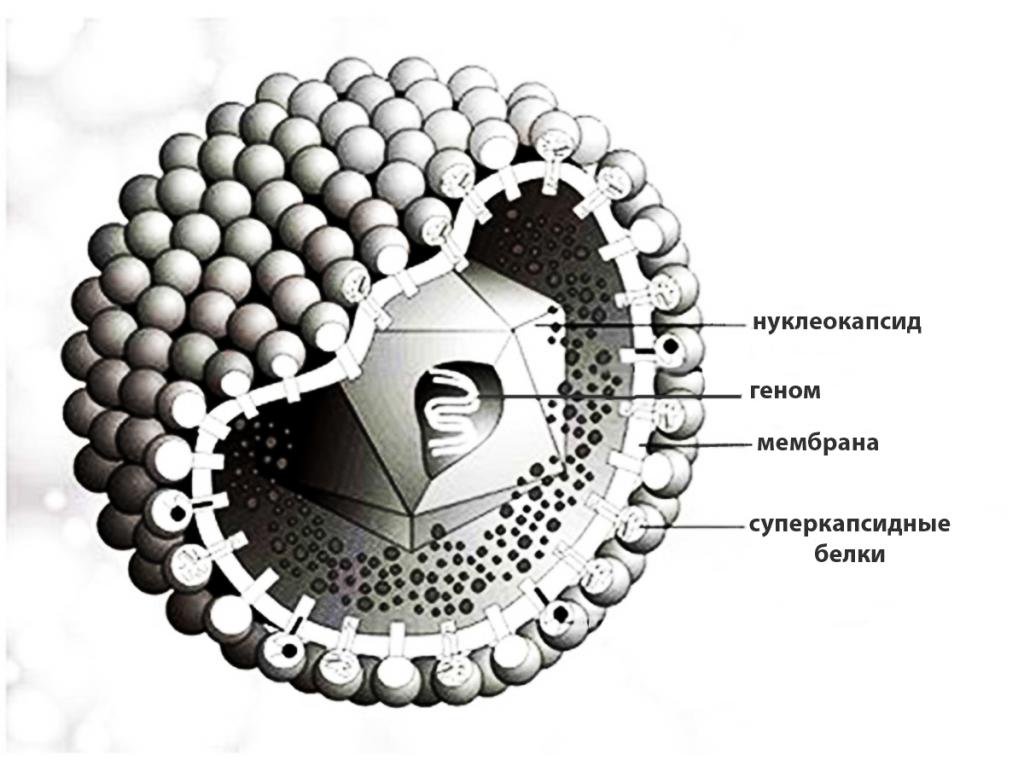
Вирусная частица состоит из нуклеиновой кислоты (РНК или ДНК), окруженной белковым слоем (капсидом), который выполняет защитные функции и обеспечивает взаимодействие с клеткой хозяина. Некоторые вирионы имеют дополнительную оболочку в виде билипидной мембраны, пронизанной торчащими наружу шипами белков вируса. Эта структура имеет клеточное происхождение и называется суперкапсидом. Размеры вирусных частиц варьируются в пределах от 20 до 200 нм.
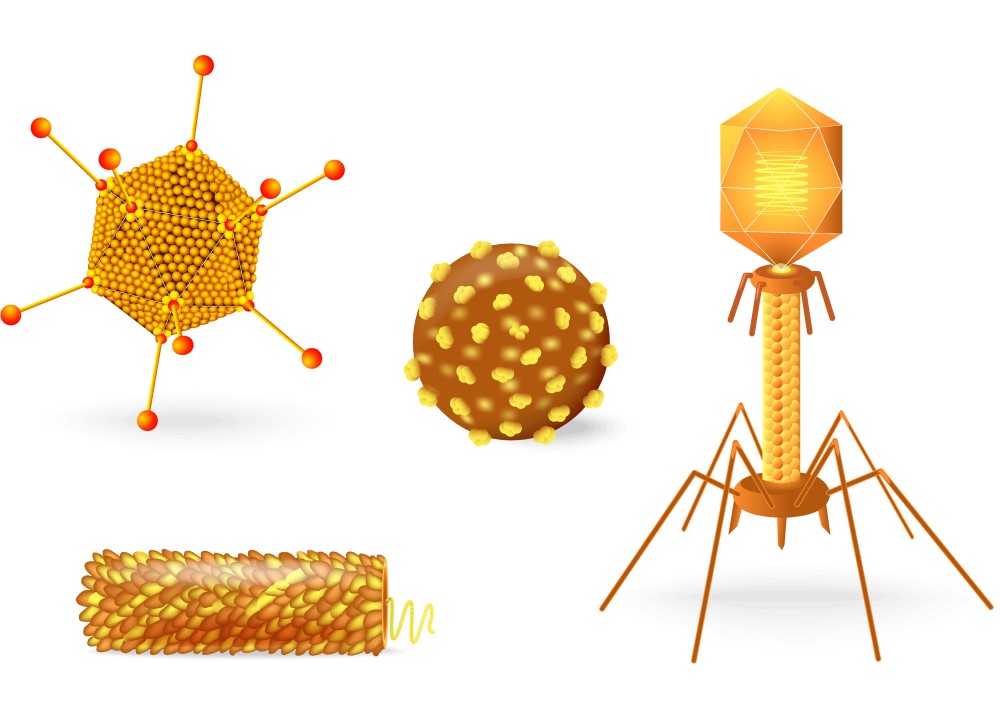
Белковые субъединицы оболочки вириона могут складываться в различные пространственные конфигурации, на основании которых строится морфологическая классификация вирусов. По типу структурной организации выделяют вирионы:
- со спиральной симметрией – белковые единицы располагаются по спирали, в центре которой пролегает аналогично структурированная нуклеиновая кислота;
- с кубической симметрией – сформированные из белковых молекул равносторонние треугольники (капсомеры) образуют различные формы многогранников (тетраэдры, октаэдры, икосаэдры и др.);
- с бинарной (смешанной) симметрией – совмещение обоих типов организации в одной вирусной частице (характерно для бактериофагов);
- сложно организованные, покрытые суперкапсидом.
Помимо структурных субъединиц оболочки некоторые вирионы содержат ферменты, необходимые для транскрипции генетического материала.
Пространственная структура, белковый состав и тип нуклеиновой кислоты вириона – это основные таксономические признаки биологической дифференциации вирусов. Дополнительными критериями являются особенности жизненного цикла и спектр хозяев.
Генетический материал вирусных частиц
В отличие от генетического материала других организмов, вирионы вирусов содержат только один тип нуклеиновой кислоты: ДНК либо РНК. Эти молекулы могут быть кольцевыми или линейными, фрагментированными или цельными, с замкнутыми (полностью или частично) или свободными концами, содержать как две цепочки, так и одну. Такое разнообразие организации нуклеиновых кислот характерно только для вирусов.
Вирусный геном также имеет и функциональную характеристику. Так, РНК вириона может быть позитивной, то есть транслироваться в клетке хозяина с образованием вирусных белков, и негативной, не обладающей матричной активностью (в данном случае трансляции предшествует синтез позитивной РНК входящим в состав вируса ферментом – транскриптазой).
В зависимости от комбинации этих характеристик в составе вирионов различают 6 типов РНК:
- одноцепочечная нефрагментированная позитивная;
- одноцепочечная нефрагментированная негативная;
- одноцепочечная фрагментированная негативная;
- двуцепочечная фрагментированная негативная;
- одноцепочечная двойная позитивная;
- одноцепочечная кольцевая дефектная.
В ДНК-геноме различают "+" и "-" цепи и выделяют следующие типы молекулярной организации:
- частично одноцепочечная кольцевая;
- сверхспиральная замкнутая кольцевая;
- одноцепочечная линейная;
- линейный дуплекс;
- линейный дуплекс с ковалентно сшитыми концами;
- одноцепочечная линейная;
Среди всех типов геномов выделяют группы, для каждой из которых характерен определенный механизм репликации в инфицированной клетке.
Сборка вириона внутри клетки-хозяина
Формирование вирусных частиц осуществляется за счет ферментов и механизмов биосинтеза зараженной клетки, которые вирус заставляет работать на себя. Этот процесс включает несколько этапов.
Вначале генетический материал вириона попадает в клетку хозяина. При этом у простых вирусов белковая оболочка остается за пределами, а у сложных проникает внутрь благодаря слиянию суперкапсида с плазматической мембраной (рецепторный эндоцитоз). В последнем случае оказавшийся в цитоплазме капсид разрушается под действием литических ферментов фагосомы.
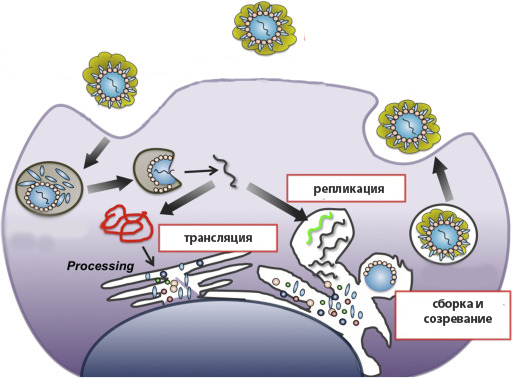
На основе нуклеиновой кислоты параллельно протекают 2 процесса: репликация генома (создание множества копий генетических молекул ДНК или РНК) и трансляция белков вириона в рибосомном аппарате клетки-хозяина.
Синтезированные белковые и генетические элементы соединяются в нуклеокапсид – полноценный вирион простых вирусов. У сложных сборка завершается в момент выхода частицы из клетки, в процессе которой происходит обволакивание капсида плазматической мембраной, содержащей заранее встроенные в нее рецепторные белки.
Научно-производственное объединение "Вирион"
Научно-исследовательское предприятие "Вирион" – это крупнейший фармацевтический комплекс по созданию и производству иммунобиологических препаратов в России. В 1906 году он был основан как Томский бактериологический институт имени Ивана и Зинаиды Чуриных, а в 1953 году получил статус НИИ вакцин и сывороток. В 1988 году институт переименовали в научно-производственное объединение (НПО) "Вирион", которое впоследствии стало филиалом Московского ФГУП НПО "Микроген".

К основным направлениям работы компании относятся создание и производство иммуноглобулинов, противовирусных вакцин, пробиотиков, психотропных лекарств и различных препаратов для диагностики. Предприятие расположено по адресу г. Томск, улица Ивановского 8.
В настоящее время производственный комплекс "Вирион" – это известная крупная компания с высокотехнологичной базой производства и профессиональным штатом сотрудников из 600 человек.
В вирусологии внутриклеточную форму существования вирусной частицы принято называть вирусом, внеклеточную – вирионом.
Размеры вирионов различных вирусов варьируют в широких пределах: от 15– 18 до 300– 400 нм. Они могут иметь разнообразную форму: палочковидную и цилиндрическую (вирус табачной мозаики и другие вирусы растений), нитевидную (вирусы растений и некоторые бактериальные вирусы), сферическую, напоминающую многогранники (аденовирусы, пикорнавирусы и др.), кубовидную (поксвирусы), сперматозоидную (большинство вирусов бактерий).
Вирионы в отличие от всех клеточных форм живых существ содержат только один тип нуклеиновой кислоты – ДНК или РНК. Из нескольких сотен (около 500) известных в настоящее время вирусов человека и животных, РНК – геном содержит около 80% вирусов. Способность РНК хранить наследственную информацию является уникальной особенностью вируса.
Генетический материал вирусов крайне разнообразен. Различают ДНК- и РНК-содержащие вирусы. Они обычно гаплоидны, т.е. имеют один набор генов. Исключением являются ретровирусы, имеющие диплоидный геном (РНК-двунитевые вирусы). Геном вирусов содержит от шести до нескольких сотен генов и представлен различными видами НК: двунитевыми, однонитевыми, линейными, кольцевыми, фрагментированными.
Среди РНК-содержащих вирусов различают вирусы с положительным (плюс-нить РНК) геномом. Плюс-нить РНК этих вирусов выполняет наследственную (геномную) функцию и функцию информационной РНК (иРНК). Имеются также РНК-содержащие вирусы с отрицательным (минус-нить РНК) геномом. Минус-нить РНК этих вирусов выполняет только наследственную функцию.
Геном вирусов способен включаться в геном клетки в виде провируса, проявляя се-бя генетическим паразитом клетки. Нуклеиновые кислоты некоторых вирусов, например, вирусов герпеса, могут находиться в цитоплазме инфицированных клеток, напоминая плазмиды.
Вирион состоит из центрально расположенной нуклеиновой кислоты (ДНК или РНК), плотно упакованной в одну или две оболочки.
Первая оболочка, в которую заключена нуклеиновая кислота - белковая. Она получила название – капсид (от греч.капса - ящик). Капсид состоит из повторяющихся белковых субъединиц - капсомеров, хорошо видных в электронном микроскопе. Просто устроенные вирусы состоят из НК и капсида. По морфологической структуре капсид представляет собой нуклеокап–сид, а по химическим свойствам может быть охарактеризован как нуклео–протеид. Нуклеиновая кислота вирусов может находить–ся внутри капсида как в свободном виде, так и в соединении с вну–тренним белком.
В отличие от просто устроенных вирусов многие сложноорганизованные вирионы имеют внешнюю оболочку (суперкапсид). Эта наружная оболочка кроме белков может содержать липиды и углеводы, в том числе "захваченные" и из клетки хозяина, в которой воспроизводились вирусные частицы. В суперкапсиде могут присутствовать вирусоспецифические углеводсодержащие белки - гликопротеиды, образующие на оболочке выступы-шипы. Как видите, вирус очень надёжно защищает свою наследственную информацию от неблагоприятных внешних условий. Т.о., сложно устроенные вирусы состоят из НК, капсида и липопротеиновой оболочки – супепкапсида.
Вирусные капсиды имеют удивительно упорядоченную организацию, в основе которой лежат принципы спиральной, кубической или сложной (комбинированной) симметрии. Капсиды состоят из структурных белковых субъединиц. Структурные асимметричные субъединицы объединены в симметричные группы, названные морфологическими субъединицами, или капсомерами, которые видны в электронном микроскопе.
Капсид и суперкапсид выполняют следующие функции:
защищают вирионы от воздействия окружающей среды,
обусловливают избирательное взаимодействие (адсорбцию) с определенными клетками хозяина,
обусловливают антигенные и иммуногенные свойства вирионов.
Наряду с белками капсида имеются еще так называемые внутренние белки, представляющие собой основные гистонодоподобные белки. Внут-ренние структуры вирусов называют сердцевиной. У аденовирусов сердцевина состоит из гистоноподобных белков, связанных с ДНК, у реовирусов – из белков внутреннегокапсида. Они находятся внутри капсида и связаны с вирусной нуклеиновой кислотой. По-видимому, внутренние белки могут образовывать рибо- или дезоксирибонуклеопротеид, покрывающий вирусную нулеиновую кислоту.
Вирусные внутренние белки и нуклеопротеид обладают видовой спе-цифичностью, наделяя вирус антигенными свойствами.
Основным структурным компонентом вирионов (полных вирусных частиц) является нуклеокапсид, т.е. белковый чехол (капсид) в котором заключен вирусный геном (ДНК или РНК). Нуклеокапсид большинства семейств вирусов окружен липопротеиновой оболочкой. Между оболочкой и нуклеокапсидом у некоторых вирусов (орто-, парамиксо-, рабдо-, фило- и ретровирусов) находится негликозилированный матриксный белок, придающий дополнительную жесткость вирионам. Вирусы большинства семейств имеют оболочку, которая играет важную роль в инфекционности. Наружный слой оболочки вирионы приобретают, когда нуклеокапсид проникает через клеточную мембрану почкованием.
Белки оболочки кодируются вирусом, а липиды заимствуются из мембраны клетки. Гликопротеины обычно в виде димеров и тримеров образуют пепломеры (выступы) на поверхности вирионов (орто-, парамиксовирусы, рабдо-, фило-, корона-, бунья-, арена-, ретровирусы). Гликозилированные белки слияния связаны с пепломерами и выполняют ключевую роль в проникновении вируса в клетку. Капсиды и оболочки вирионов образуются множеством копий одного или нескольких видов белковых субъединиц в результате процесса самосборки. Взаимодействие в системе белок-белок, благодаря слабым химическим связям, ведет к объединению симметричных капсидов.
Различия вирусов по форме и размеру вирионов зависят от формы, размера и количества структурных белковых субъединиц и природы взаимодействия между ними.
Капсид состоит из множества морфологически выраженных субъединиц (капсомеров), собранных из вирусных полипептидов строго определенным образом, в соответствии с относительно простыми геометрическими принципами. Белковые субъединицы, соединяясь друг с другом, образуют капсиды двух видов симметрии: изометрические и спиральные. Структура нуклеокапсида оболочечных вирусов сходна со структурой нуклеокапсида безоболочечных вирусов. На поверхности оболочки вирусов различают морфологически выраженные гликопротеиновые структуры — пепломеры.
В состав суперкапсидной оболочки входят липиды (до 20—35%) и углеводы (до 7—8%), имеющие клеточное происхождение. Она состоит из двойного слоя клеточных липидов и вирусспецифических белков, расположенных снаружи и изнутри липидного биослоя. Наружный слой суперкапсидной оболочки представляют пепломеры (выступы) одного или более типов, состоящие из одной или нескольких молекул гликопротеинов. Нуклеокапсид у оболочечных вирусов часто называют сердцевиной (core), а центральную часть вирионов, содержащую нуклеиновую кислоту, называют нуклеоидом.
Капсомеры (пепломеры) состоят из структурных единиц, построенных из одной либо из нескольких гомологичных или гетерологичных полипептидных цепей (белковых субъединиц).
Изометрические капсиды представляют собой не сферы, а правильные многогранники (икосаэдры). Их линейные размеры идентичны по осям симметрии. Согласно Каспару и Клугу (1962), капсомеры в капсидах расположены в соответствии с икосаэдрической симметрией.
Такие капсиды состоят из идентичных субъединиц, образующих икосаэдр. Они имеют 12 вершин (углов), 30 граней и 20 поверхностей в виде равнобедренных треугольников. В соответствии с этим правилом капсид полиовируса и вируса ящура образован 60 белковыми структурными единицами, каждая из которых состоит из четырех полипептидных цепей.
Икосаэдр оптимально решает проблему укладки повторяющихся субъединиц в строгую компактную структуру при минимальном объеме. Только некоторые конфигурации структурных субъединиц могут сформировать поверхности, образовать вершины и грани вирусного икосаэдра. Например, структурные субъединицы аденовируса на поверхностях и гранях формируют шестигранные капсомеры (гексоны), а на вершинах - пятигранные капсомеры (пептоны). У одних вирусов оба вида капсомеров образуются одними и теми же полипептидами, у других — разными полипептидами. Так как структурные субъединицы разных вирусов различаются между собой, то одни вирусы кажутся более гексагональными, другие — более сферическими.
Все известные ДНК-содержащие вирусы позвоночных, за исключением вирусов оспы, а также многие РНК-содержащие вирусы (7 семейств) имеют кубический тип симметрии капсида.
Реовирусы, в отличие от других вирусов позвоночных, имеют двойной кап-сид (наружный и внутренний), причем каждый состоит из морфологических единиц.
Вирусы, обладающие спиральным типом симметрии, имеют вид цилиндрической нитевидной структуры, их геномная РНК имеет вид спирали и находится внутри капсида. Все вирусы животных спиральной симметрии окружены липопротеиновой оболочкой.
Спиральные нуклеокапсиды характеризуются длиной, диаметром, шагом спирали и числом капсомеров, приходящихся на один оборот спирали. Так, у вируса Сендай (парамиксовирус) нуклеокапсид представляет собой спираль длиной около 1 мкм, диаметром 20 нм и шагом спирали 5 нм. Капсид состоит примерно из 2400 структурных единиц, каждая из которых является белком с молекулярной массой 60 кД. На каждый виток спирали приходится 11—13 субъединиц.
У вирусов со спиральным типом симметрии нуклеокапсида укладка белковых молекул в спираль обеспечивает максимальное взаимодействие между нуклеиновой кислотой и белковыми субъединицами. У икосаэдрических вирусов нуклеиновая кислота находится внутри вирионов в скрученном состоянии и взаимодействует с одним или несколькими полипептидами, расположенными внутри капсида.
Простые (безоболочечные) вирусы состоят из нуклеиновой кислоты и белка и представляют собой нуклеопротеиды или нуклеокапсиды. Сложные (оболочечные) вирусы кроме нуклеиновой кислоты и белка содержат также липиды и углеводы.
Нуклеиновые кислоты. Клетки всех живых организмов содержат два типа нуклеиновых кислот — ДНК и РНК. Их структура относительно однообразна: ДНК представляет собой двуспиральную молекулу, РНК — односпиральную молекулу. Функция ДНК заключается в хранении и репликации наследственной информации, таким образом, она является клеточным геномом. РНК представлена в клетке в трех формах: информационной (иРНК), рибосомальной (рРНК) и транспортной (тРНК). Каждая из них выполняет определенную функцию: иPHК образуется в результате транскрипции генома и передает информацию с генома на белоксинтезирующий аппарат клетки, рРНК является структурным компонентом рибосом, тРНК доставляет аминокислоты белоксинтезирующему аппарату клетки.
В отличие от клетки вирусы содержат один тип нуклеиновой кислоты — или ДНК, или РНК. Каждая из них выполняет функцию вирусного генома. Структура нуклеиновых кислот у разных вирусов весьма разнообразная. По количеству цепей они бывают одно — и двуспиральными, по форме — линейными и кольцевыми (циркулярными), а также непрерывными и фрагментированными.
Содержание нуклеиновой кислоты в вирионе различных вирусов составляет от 1 % у ортомиксо- и парамиксовирусов до 32 % у парвовирусов и не коррелирует ни с систематическим положением вируса, ни со степенью сложности его организации.
Вирусные ДНК. Молекулярная масса ДНК различных вирусов варьирует в широких пределах: от 2 МД у цирко — и парвовирусов до 375 МД у поксвирусов. Самые большие геномы содержат до нескольких сотен генов, самые маленькие — несколько генов. По структуре молекулы ДНК бывают одно — и двуспиральными, линейными и кольцевыми. У вирусов с двуспиральными ДНК информация обычно закодирована на обеих спиралях, что говорит о максимальной экономии генетического материала. Большинство нуклеотидных последовательностей в молекуле ДНК встречается однократно. Однако в концевом фрагменте линейных ДНК возможно наличие ее начального участка в виде повтора, который бывает прямым или инвертируемым. Благодаря таким повторам молекулы ДНК могут приобретать циркулярную форму, которая обеспечивает их устойчивость к эндонуклеазам. Кроме того, стадия образования циркулярной формы обязательна для интеграции вирусной ДНК с геномом клетки.
Вирусные РНК. Молекулярная масса вирусных РНК варьирует в пределах от 4—5 МД у нодавирусов до 32 МД у реовирусов.
По предложению Балтимора (1971) вирусы с односпиральными РНК из-за различий в функциях генома было принято разделять на две подгруппы. У вирусов первой подгруппы вирусный геном обладает функциями иРНК и их условно обозначают как плюс-нитевые вирусы, или вирусы с позитивным геномом. У вирусов второй группы РНК не обладает функцией иРНК. На ней, как на матрице, синтезируется комплементарная молекула. Это происходит только в присутствии вирусного белка — фермента транскриптаза, который обязательно находится в структуре минус-нитевых вирусов (в клетках — ее аналога нет).
Существуют вирусы, которые содержат как плюс-нитевые, так и минус-нитеные РНК-гены, так называемые амбигюлирные РНК.
Белки. Белки всех известных в настоящее время вирусов позвоночных являются основными компонентами вирионов и составляют от 57 до 90 % массы вириона. По аминокислотному составу вирусные белки принципиально не отличаются от состава белков животных.
В геноме вирусов кодируются две группы белков: структурные, которые входят в состав вирионов потомства, и неструктурные, участвующие в репродукции вируса на разных этапах, но не входящие в состав вирионов.
Структурные белки в составе вириона варьируют в широких пределах, что зависит от сложности организации вириона. Простые вирусы животных содержат 3—4 белка, сложные, например вирусы оспы, — более 30. Среди структурных белков различают две группы — капсидные и пепломеры. Первая группа включает белки, которые формируют капсид (от греч. capsa — вместилище), окружающий нуклеиновую кислоту, а также геномные белки и ферменты; вторая группа — белки суперкапсидной оболочки, называемой пеплос (от греч. peplos — покров, мантия). Простые вирусы содержат только капсидные белки, сложные — и капсидные, и пепломеры.
Белки в составе вирусного калсида называются капсомерами. Основной их функцией является защита вирусного генома от неблагоприятных воздействий внешней среды. Они представляют собой идентичные полипептидные цепи (белковые субъединицы), которые обладают способностью к самосборке. Сборка капсида из субъединиц запрограммирована в первичной структуре белка и происходит самопроизвольно или при взаимодействии с нуклеиновой кислотой. Принцип субъединичности в строении капсида — уникальное свойство капсидных белков, благодаря которому достигается огромная экономия генетического материала. Кроме того, в механизме самосборки заложена возможность контроля за белковыми субъединицами: дефектные и, чужеродные белковые цепи не включаются в капсид. Принцип самосборки характерен только для простых вирусов. Сложные вирусы сборку осуществляют по более сложному многоступенчатому механизму. Однако отдельные ее этапы (формирование капсидов и нуклеокапсидов) основаны на самосборке.
Суперкапсидные белки (пепломеры) находятся в липопротеидной оболочке сложных вирусов. Они либо пронизывают липидный бислой вириона, либо не доходят до его внутренней поверхности. Являясь типичными внутримембранными белками, они, как правило, гликозилированы (гликопротеиды), т. е. к молекуле белка в определенных местах прикреплены углеводные цепи. Гликозилирование осуществляют клеточные ферменты, поэтому один и тот же вирус, но реплицирующийся в разных клетках, может иметь разные углеводные остатки по составу углеводов, длине углеводной цепи, месту прикрепления к белку.
Неструктурные белки менее изучены. Существуют определенные трудности в их выделении из зараженных клеток и очистки от клеточных белков. К ним относятся: предшественники вирусных белков, которые существуют в зараженной клетке очень непродолжительное время, а затем нарезаются; ферменты синтеза РНК. и ДНК — полимеразы; регуляторы стадий репродукции вирусов; ферменты, модифицирующие вирусные белки — протеиназы и протеинкиназы.
Липиды и углеводы. В состав вирионов всех сложных (оболочечных) вирусов позвоночных кроме нуклеиновой кислоты и белков входят липиды и углеводы.
Состав липидов вирионов сходен с липидным составом клетки хозяина: примерно 50—60 % составляют фосфолипиды и 20—30 % — холестерин. У отдельных представителей липидов содержится до 20—35 % от массы вириона (ортомиксо-, ретро-, буньявирусы). Липиды обнаружены только в суперкапсидной оболочке вирионов и имеют клеточное происхождение. Это связано с тем, что оболочечные вирусы формируются путем почкования на плазматической мембране клеток. Поэтому суперкапсидная оболочка вирионов представляет собой мембрану клетки-хозяина, модифицированную за счет встроенных в нее вирусных белков — пепломеров. Липидный компонент стабилизирует структуру вирусных частиц, поэтому их обработка детергентами или липазами приводит к потере инфекционности.
Исключение составляют вирусы оспы. У них липиды не образуют дифференцированной оболочки. Обработка вирусов осповакцины жирорастворителями не приводит к потере инфекционной активности или каким-либо другим структурным изменениям вириона.
Углеводы находятся в вирионах в виде гликопротеинов, встроенных в суперкапсидный слой, а также гликолипидов и имеют клеточное происхождение. У отдельных представителей вирусов позвоночных содержание углеводов доходит до 7—8 % от массы вириона (ортомиксо-, тогавирусы). Химический состав их полностью определяется клеточными ферментами, которые обеспечивают перенос и присоединение сахарных остатков. В вирионах в основном обнаруживают фруктозу, сахарозу, маннозу, галактозу, нейраминовую кислоту, глюкозамин. Углеводы являются каркасом для локальных участков гликопротеидов, обеспечивают сохранение конформации белковых молекул и защищают от действия протеаз.
Компоненты клетки-хозяина. В составе вирионов могут обнаруживаться некоторые компоненты клетки-хозяина. Это могут быть белки или цельные клеточные структуры. Например, в составе некоторых оболочечных вирусов находится белок цитоскелета актин, в составе папиллома — и полиомавирусов содержатся клеточные гистоны, у ареновирусов обнаружены рибосомы. Клеточные компоненты могут включаться в вирион случайно или закономерно. Во втором случае они могут играть существенную роль в репродукции вирусов, как, например, гистоны для папиллома — и полиомавирусов.
Структурная организация вирионов. Вирионы (вирусные частицы) по архитектуре подразделяются на два типа: имеющие внешнюю липопротеидную оболочку (сложные, или оболочечные, вирусы) и не имеющие такой оболочки (простые, или безоболочечные, вирусы).
Капсомеры соединяются друг с другом строго определенным образом и образуют только два типа капсидов: спиральный и изометрический (квазиметрически й). Только такие способы укладки капсомеров вокруг нуклеиновой кислоты (типы симметрии) обеспечивают формирование энергетически экономичных структур с минимумом свободной энергии.
Изометрические капсиды представляют структуры, в которых капсомеры соединяются между собой в правильные многогранники, в центре которых расположен геном. Такой способ укладки капсомеров называется кубическим типом симметрии, а многогранник — икосаэдром. Это означает, что он симметричен в трех взаимно перпендикулярных направлениях и его линейные размеры вдоль прямоугольных осей идентичны. Обычно изометрические капсиды состоят из 60 (или кратных 60) геометрически идентичных элементов, которые имеют 12 вершин, 20 граней и 20 ребер. Так, капсид вируса полиомиелита образован 60 белковыми структурными единицами, каждая из которых состоит из четырех полипептидных цепей (химических единиц) — vp1, vp2, vp3, vp4 (virion protein).
Наиболее сложно структурно организованы вирусы оспы. Их сердцевина (нуклеоид), содержащая вирусную ДНК в составе нуклеотида, имеет форму двояковогнутого кольца и окружена двумя линзообразными латеральными тельцами. Капсид окружен дополнительными внутренними белковыми структурами — вирусным матриксом. По архитектуре вирусы оспы отличаются от всех других вирусов позвоночных и их принято называть вирусами со сложным типом симметрии.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Вирусы – неклеточная форма жизни, обладает собственным геномом, способностью к самовоиспроизведению (репродукции) в клетках живых организмов или клеточных культурах, адаптационными свойствами и изменчивостью.
Выделены в отдельное царство – Vira.
- нет клеточной организации: не имеют цитоплазмы и ядра, митохондрий, рибосом и других органелл
- содержат только одну из двух нуклеиновых кислот – ДНК или РНК, выполняющих функции генома.
- не имеют собственных белоксинтезирующих и генерирующих энергию систем и являются абсолютными внутриклеточными паразитами на генетическом уровне, полностью зависят от клетки-хозяина
- размножаются не обычным бинарным делением, а репродуцируются в чувствительной клетке, согласно генетической программе в нуклеиновой кислоте вируса, при этом используют биосинтетические системы и ресурсы
Различают две формы существования вирусов – внеклеточную и внутриклточную.
Внеклеточный вирус = вирион. Это покоящаяся (зрелая) форма вируса. Не проявляет жизедеятельности. Функции: сохранение вируса во внешней среде и перенос его из организма в другой организм или из клетки в другую клетку.
Внутриклеточный вирус - вегетативный вирус - репродуцируется в инфицированной клетке, вызывая репродуктивную инфекцию, заканчивающуюся образованием дочернего поколения вирионов и, как правило, гибелью клетки. Процесс репродукции может быть незавершенным, без образования вирионов – возникает абортивная инфекция.
Некоторые вирусы способны встраивать свой генетический материал в хромосомы клетки-хозяина в виде провируса, которые реплицируется вместе с этой хромосомой в процессе деления и переходит в дочерние клетки. Это – интегративная инфекция, она модет существовать длительное время или переходить обратно в продуктивную.
строение вирусов (вирионов). Размеры вирусов находятся в диапазоне 20-350 нм.
Могут иметь палочковидную, многогранную, пулевидную, сферическую, нитевидную, булавовидную формы.
Различают: простые (безоболочечные) и сложные (оболочечные) вирусы. У них в центре – молекула нуклеиновой кислоты (ДНК/РНК), окруженная белковой оболочкой – капсидом. Вся структура носит название – нуклокапсид.
Простые вирусы – нуклеиновая кислота, ассоциированная с внутренними белками и капсидом (т.е. представляют собой нуклеокапсид).
Защитная белковая оболочка – капсид – состоит из множества однородных белковых субъединиц. Т.к. на такое строение капсида расходуется мало генетической информации, оно важно для вирусов, обладающих небольшим геномом. Капсиды построены по спиральному или кубическому типу симметрии, в зависимости от расположения белковых субъединиц.
Химический состав вируса.Основные компоненты вируса – нуклеиновая кислота и белки. Простые вирусы состоят только из них. В состав сложных вирусов входят углеводы и липиды клеточного происхождения.
В зависимости от типа нуклеиновой кислоты вирусы делят на ДНК- и РНК-геномные.
Вирусные ДНК – обычно двунитевые, редко – однонитевые.
Двунитевые ДНК: линейные с незамкнутыми концами, линейные с замкнутыми концами, кольцевидные, кольцевидные с одной неполной цепью ДНК.
Вирусные РНК – однонитевые, бывают двунитевые с фрагментированным геномом.
Однонитевые РНК: цельные линейные, фрагментированные (сегментированные) линейные, кольцевые сегментированные.
Различают РНК с положительным геномом – +РНК (одновременно геном и информационная РНК (и-РНК), служит матрицей для дочерних геномов);
и РНК с отрицательным геномом – –РНК (только геномная функция, т.е. матрица для синтеза генома и и-РНК).
Важнейшая особенность вирусных нуклеиновых кислот – инфекционность (способность инициировать в клетке –хозяине продуктивную инфекцию без участия других компонентов вируса). Ей обладает большинство вирусных ДНК и +РНК.
Вирусные белки.
# структурные – входят в состав вириона:
- капсидные белки- формируют капсид
- внутренние белки – геномные белки и ферменты (полимеразы), участвующие в процессе репродукции и ассоциации генома с капсидом.
- матриксные белки сложных вирусов, образуют М-слой под суперкапсидом. Участвуют в заключительных этапах самосборки вирионов и их стабилизации.
- суперкапсидные поверхностные белки – гликопротеины, протективные Аг, участвуют в прикреплении вирионов к клеточным рецепторам и их проникновении в клетку.
# Неструктурные белки – синтезируются в инфицированной клетке для обеспечения процессов репродукции, в состав вирусов не входят.
- вирусиндуцированные ферменты, обслуживают транскрипцию и трансляцию вирусного генома.
- регуляторные белки
- нестабильные белки – предшественники, из которых формируются структурные белки вириона
- ферменты, модифицирующие вирусные белки (протеазы, протеинкиназы)
Липиды. Переходят в состав вирионов из клеточных, ядерных, других внутренних мембран инфицированной клетки при почковании. Являются основным компонентом суперкапсид, способствуют стабильности вириона. При обработке эфиром суперкапсид разрушается из-за потери липидов.
Углеводы. Клеточное происхождение. Входят в состав поверхностных белков – гликопротеинов. Их гликолизирование осуществляется клеточными ферментами во время транспортировки белков на наружную поверхность суперкапсида, при этом клеточные белки вытесняются из мембран.
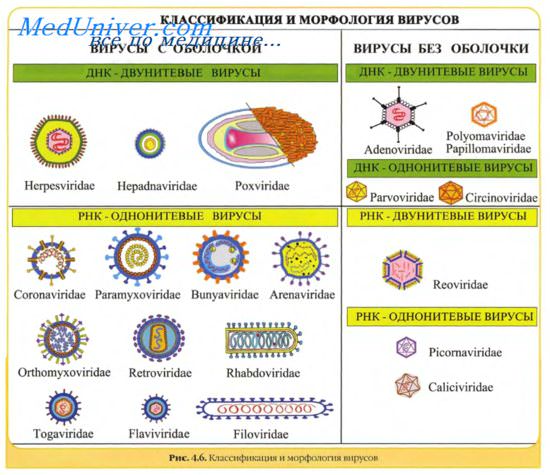
В основу классификации вирусов положены следующие категории:
• тип нуклеиновой кислоты (ДНК или РНК), ее структура, количество нитей (одна или две), особенности воспроизводства вирусного генома;
• размер и морфология вирионов, количество капсомеров и тип симметрии;
• чувствительность к эфиру и дезоксихолату;
• место размножения в клетке;
• антигенные свойства и пр.
Вирусы имеют уникальный геном, так как содержат либо ДНК, либо РНК. Поэтому различают ДНК-содержащие и РНК-содержащие вирусы. Они обычно гаплоидны, т.е. имеют один набор генов. Геном вирусов представлен различными видами нуклеиновых кислот: двунитчатыми, однонитчатыми, линейными, кольцевыми, фрагментированными. Среди РНК- содержащих вирусов различают вирусы с положительным (плюс-нить РНК) геномом. Плюс-нить РНК этих вирусов выполняет наследственную функцию и функцию информационной РНК (иРНК). Имеются также РНК-содержащие вирусы с отрицательным (минус-нить РНК) геномом. Минус-нить РНК этих вирусов выполняет только наследственную функцию.
Форма вирионов может быть различной: палочковидной (вирус табачной мозаики), пулевидной (вирус бешенства), сферической (вирусы полиомиелита, ВИЧ), нитевидной (филовирусы), в виде сперматозоида (многие бактериофаги). Различают просто устроенные и сложно устроенные вирусы.
Простые, или безоболочечные, вирусысостоят из нуклеиновой кислоты и белковой оболочки, называемой капсидом. Капсид состоит из повторяющихся морфологических субъединиц — капсомеров. Нуклеиновая кислота и капсид взаимодействуют друг с другом, образуя нуклеокапсид.
Сложные, или оболочечные, вирусыснаружи капсида окружены ли-попротеиновой оболочкой (суперкапсидом, или пеплосом). Эта оболочка является производной структурой от мембран вирус-инфицированной клетки. На оболочке вируса расположены гликопротеиновые шипы, или шипики (пепломеры). Под оболочкой некоторых вирусов находится матриксный М-белок.
Последнее изменение этой страницы: 2017-01-19; Нарушение авторского права страницы
Читайте также: