
Этапы синтеза белка вируса
Размножение вирусов не осуществляется бинарным делением. Еще в 50-х годах прошлого века было установлено, что размножение осуществляется методом репродукции (в переводе с англ. reproduce – делать копию, воспроизводить), то есть путем воспроизведения нуклеиновых кислот, а также синтеза белка с последующим сбором вирионов. Данные процессы происходят в различных частях клетки так называемого хозяина (к примеру, в ядре или цитоплазме). Данный разобщенный метод репродукции вирусов называется дизъюнктивным. Именно на этом мы и остановимся подробнее в нашей статье.

Процесс репродукции
Данный процесс имеет свои особенности репродукции вирусов и отличается последовательной сменой некоторых стадий. Рассмотрим их по отдельности.
Вирусы не могут размножаться в питательной среде, так как они представляют собой строгие внутриклеточные паразиты. Кроме того, в отличие от хламидий или риккетсий, во время репродукции вирусы в клетке хозяина не способны расти и не размножаются методом деления. Все составные части данного вируса включают в себя нуклеиновые кислоты, а также белковые молекулы, которые синтезируются в "хозяйской" клетке раздельно, в различных частях клетки: в цитоплазме и в ядре. Помимо этого, белоксинтезирующие клеточные системы подчиняются одному вирусному геному, а также его НК.

Вирусная репродукция в клетке осуществляется в несколько фаз, которые описаны ниже:
Адсорбция
Под этой стадией репродукции вирусов подразумевается проникновение вирусной частицы в клетки. Адсорбция начинается на клеточной поверхности при помощи взаимодействия клеточных, а также вирусных рецепторов. В переводе с латинского слово "рецепторы" означает "принимающий". Они представляют собой специальные чувствительные образования, которые воспринимают раздражения. Рецепторы - это молекулы либо молекулярные комплексы, расположенные на поверхности клеток, а также способны распознавать химические специфические группировки, молекулы либо другие клетки, связывать их. У наиболее сложных вирионов такие рецепторы располагаются с внешней оболочки в виде шиповидного выроста или ворсинки, у простых вирионов они находятся, как правило, на поверхности капсида.

Механизм адсорбции на поверхности восприимчивой клетки основывается на взаимодействии рецепторов с так называемыми комплементарными рецепторами "хозяйской" клетки. Рецепторы вириона и клетки являются некими специфическими структурами, которые расположены на поверхности.
Аденовирусы и миксовирусы адсорбируются непосредственно на мукопротеиновых рецепторах, а арбовирусы и пикорнавирусы ― на липопротеиновых рецепторах.
У вириона миксовирусов нейраминидаза разрушает мукогфотеиновый рецептор и отщепляет N-ацетилнейраминовые кислоты от олигосахарида, который содержит в себе галактозу и галактозамин. Их взаимодействия на данном этапе обратимы, ведь на них значительно влияет температура, реакция среды и солевые компоненты. Адсорбции вириона препятствуют гепарин и сульфатированные полисахариды, несущие при этом отрицательный заряд, однако их ингибирующее воздействие снимается некоторыми поликарионами (экмолин, ДЭАЭ-декстран, протаминсулъфат), нейтрализующие отрицательный заряд от сульфатированных полисахаридов.
Попадание вириона в "хозяйскую" клетку
Путь внедрения вируса в чувствительную к нему клетку не всегда будет одним и тем же. Многие вирионы способны проникать в клетки методом пиноцитоза, что в переводе с греческого означает "пить", "выпивать". При данном методе пиноцитозная вакуоль будто бы втягивает вирион непосредственно внутрь клетки. Остальные вирионы могут проникать в клетку напрямую сквозь ее оболочку.

Контакт фермента нейраминидаза с клеточными мукопротеидами способствует попаданию вирионов в клетку среди миксовирусов. Результаты исследований последних лет доказывают, что ДНК и РНК вирионов от внешней оболочки не отделяются, т. е. вирионы проникают целиком в чувствительные клетки путем пиноцитоза или виропексиса. На настоящий момент это подтверждено в отношении вируса оспы, осповакцины, а также других вирусов, выбирающих средой обитания организм животных. Если говорить о фагах, они заражают нуклеиновой кислотой клетки. Механизм заражения основывается на том, что те вирионы, которые содержатся в вакуолях клеток, гидролизуются ферментами (липаз, протеаз), в процессе чего от оболочки фага освобождается ДНК и попадает в клетку.
Для проведения эксперимента выполнялось заражение клетки с помощью нуклеиновой кислоты, которая была выделена от некоторых вирусов, и вызывается один полный цикл репродукции вирионов. Однако в естественных условиях инфицирования при помощи такой кислоты не происходит.
Дезинтеграция
Следующий этап репродукции вирусов – дезинтеграция, которая представляет собой освобождение НК от капсида и внешней оболочки. После попадания вириона в клетки, капсид переживает некоторые изменения, приобретая чувствительность к клеточному протеазу, затем он разрушается, параллельно освобождая НК. У отдельных бактериофагов в клетки попадает свободная НК. Фитопатогенный вирус проникает через повреждение в клеточной стенке, а затем он адсорбируется на внутреннем клеточном рецепторе с одновременным высвобождением НК.
Репликация РНК и синтез вирусного белка
Следующим этапом репродукции вирусов является синтез вирусоспецифичного белка, который происходит с участием так называемых информационных РНК (у отдельных вирусов они находятся в составе вирионов, а у некоторых синтезируются только в зараженных клетках непосредственно на матрице вирионной ДНК или РНК). Происходит репликация вирусной НК.

Существует 2 вида РНК-полимераз. К таковым относятся: РНК-полимераза I, которая катализирует формирование репликативной формы непосредственно на матрице плюс-цепи, а также РНК-полимераза II, которая принимает участие в синтезе однонитчатой вирусной РНК на матрице репликативного типа. Синтез нуклеиновых кислот у мелких вирусов происходит в цитоплазме. Что касается вируса гриппа, то в ядре синтезируется внутренний белок и РНК. РНК выделяется затем из ядра и проникает в цитоплазму, в которой совместно с рибосомами начинает синтезировать вирусный белок.
После попадания вирионов в клетки, в них подавляется синтез нуклеиновой кислоты, а также клеточных белков. При репродукции вирусов, ДНК содержащих, на матрице в ядре синтезируется еще и-РНК, которая несет в себе информацию для синтеза белка. Механизм синтеза вирусного белка осуществляется на уровне клеточной рибосомы, а источником построения будет аминокислотный фонд. Активизация аминокислот осуществляется ферментами, при помощи и-РНК переносятся непосредственно в рибосомы (полисомы), в которых они располагаются уже в синтезированной молекуле белков.
Таким образом, в зараженных клетках синтез нуклеиновых кислот и белков вириона осуществляется в составе репликативно-транскриптивного сложного комплекса, который регулируется некой системой механизма.

Морфогенез вириона
Образование вирионов может произойти только в случае строго упорядоченного соединения структурных вирусных полипептидов, а также их НК. А это обеспечивается так называемой самосборкой молекул белка около НК.
Формирование вириона
Формирование вириона происходит с участием некоторых структурных компонентов, входящих в состав клетки. Вирусы герпеса, полиомиелита и осповакцины образуются в цитоплазме, а аденовирусы ― в ядре. Синтез вирусной РНК, а также формирование нуклеокапсида происходит непосредственно в ядре, а гемагглютинин формируется в цитоплазме. После этого нуклеокапсид перебирается из ядра в цитоплазму, в которой осуществляется образование оболочки вириона. Нуклеокапсид покрывается снаружи вирусными белками, а в состав вириона при этом включаются гемагглютинины и нейраминидазы. Именно таким образом происходит образование потомства, например, вируса гриппа.
Высвобождение вириона из "хозяйской" клетки
Из "хозяйской" клетки частицы вируса выделяются одновременно (во время разрушения клеток) либо постепенно (без каких-либо разрушений клеток).
Именно в таком виде и происходит репродукция вирусов. Вирионы высвобождаются из клеток, как правило, двумя способами.
Первый метод
Первый способ подразумевает следующее: после абсолютного созревания вирионов непосредственно внутри клетки они округляются, там образуются вакуоли, а затем разрушается и клеточная оболочка. По завершению этих процессов вирионы выходят все одновременно и полностью из клеток (пикорнавирусы). Данный способ принято называть литическим.

Второй метод
Второй способ подразумевает процесс освобождения вирионов по мере их созревания в течение 2―6 часов на цитоплазматической мембране (миксовирусы и арбовирусы). Выделению из клетки миксовирусов способствует нейраминидазы, разрушающие клеточную оболочку. Во время этого способа 75-90 % вирионов выходят спонтанно в культуральную среду, а клетки постепенно погибают.
Продуктивный тип взаимодействия вируса с клеткой, т. е. репродукция вируса (лат. re — повторение, productio — производство), проходит в 6 стадий:
1) адсорбция вирионов на клетке;
2) проникновение вируса в клетку;
4) синтез вирусных компонентов;
5) формированиевирионов;
6) выход вирионов из клетки.
У различных вирусов эти стадии отличаются
Специфические рецепторы клеток имеют различную природу, являясь белками, липидами, углеводными компонентами белков, липидов и др. Так, рецепторами для вируса гриппа является сиаловая кислота в составе гли-копротеинов и гликолипидов (ганглиозидов) клеток дыхательных путей. Вирусы бешенства адсорбируются на ацетилхолиновых рецепторах нервной ткани, а вирусы иммунодефицита человека — на СО4-рецепторах Т-хелперов, моноцитов и дендритных клеток. На одной клетке находится от десяти до ста тысяч специфических рецепторов, поэтому на ней могут адсорбироваться десятки и сотни вирионов.
Наличие специфических рецепторов лежит в основе избирательности поражения вирусами определенных клеток, тканей и органов. Это так называемый тропизм(греч. tropos — поворот, направление). Например, вирусы, репродуцирующиеся преимущественно в клетках печени, называются гепатотропными, в нервных клетках — нейротропными, в иммунокомпетентных клетках — иммунотропными и т. д.
Проникновение вирусов в клетку.Вирусы проникают в клетку путем рецептор-зависимого эндоцитоза (виропексиса), или слияния оболочки вируса с клеточной мембраной, или же в результате сочетания этих механизмов.
2. Слияние обточки вириона с клеточной мембраной характерно только для некоторых оболочечных вирусов (парамиксовирусов, ретровиру-сов, герпесвирусов), в составе которых имеются белки слияния. Происходит точечное взаимодействие вирусного белка слияния с липидами клеточной мембраны, в результате чего вирусная липопротеиновая оболочка интегрирует с клеточной мембраной, а внутренний компонент вируса попадает в цитозоль.
В) Синтез вирусных компонентов. Синтез белков и нуклеиновых кислот вируса, который разобщен во времени и пространстве. Синтез осуществляется в разных частях клетки, поэтому такой способ размножения вирусов называется дизъюнктивным (от лат. disjunctus — разобщенный).
С)Синтез вирусных белков. В зараженной клетке вирусный геном кодирует синтез двух групп белков:
1. неструктурных белков, обслуживающих внутриклеточную репродукцию вируса на разных его этапах;
2. структурных белков, которые входят в состав вириона (геномные, связанные с геномом вируса, капсидные и су-перкапсидные белки).
К неструктурным белкамотносятся: 1) ферменты синтеза РНК или ДНК (РНК- или ДНК-полимеразы), обеспечивающие транскрипцию и репликацию вирусного генома; 2) белки-регуляторы; 3) предшественники вирусных белков, отличающиеся своей нестабильностью в результате быстрого нарезания на структурные белки; 4) ферменты, модифицирующие вирусные белки, например, протеиназы и протеинкиназы.
I. ДНК-содержашие вирусы реализуют генетическую информацию так же, как и клеточный геном, по схеме:
Причем ДНК-содержашие вирусы используют для этого процесса клеточную полимеразу (вирусы, геномы которых транскрибируются в ядре клетки — аденовирусы, па-повавирусы, герпесвирусы) или собственную РНК-полимеразу (вирусы, геномы которых транскрибируются в цитоплазме, например поксвирусы).
II. Плюс-нитевые РНК-содержашие вирусы(например, пикорнавирусы, флавивирусы, тогавирусы) имеют геном, выполняющий функцию иРНК; он распознается и транслируется рибосомами. Синтез белков у этих вирусов осуществляется без акта транскрипции по схеме:
геномная РНК вируса-> трансляция белка вируса.
III. Геном минус-однонитевых РНК-содержаших вирусов (ортомиксовирусов, парамиксовирусов, рабдовирусов) и двунитевых (реовирусов) служит матрицей, с которой транскрибируется иРНК, при участии РНК-полимеразы, связанной с нуклеиновой кислотой вируса. Синтез белка у них происходит по схеме:
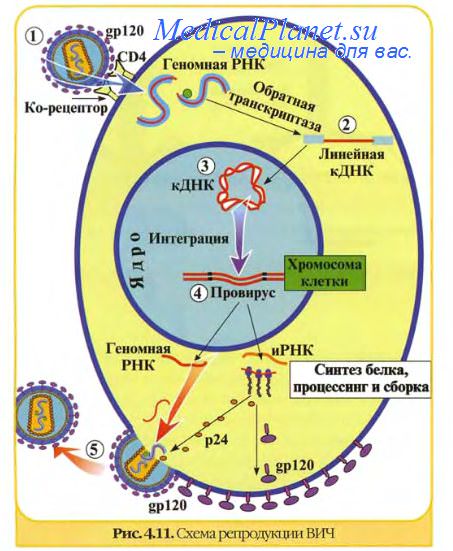
IV. Ретровирусы(вирусы иммунодефицита человека, онкогенные ретровирусы) имеют уникальный путь передачи генетической информации. Геном ретровирусов состоит из двух идентичных молекул РНК, т. е. является диплоидным. В составе ретровирусов есть особый вирусоспецифический фермент — обратная транскриптаза, или ревертаза, с помощью которой осуществляется процесс обратной транскрипции, т. е. на матрице геномной РНК синтезируется комплементарная однонитевая ДНК (кДНК). Комплементарная нить ДНК копируется с образованием двунитевой комплементарной ДНК, которая интегрирует в клеточный геном и в его составе транскрибируется в иРНК с помощью клеточной ДНК-зависимой РНК-полимеразы. Синтез белков для этих вирусов осуществляется по схеме:
Репликация вирусных геномов,т. е. синтез вирусных нуклеиновых кислот, приводит к накоплению в клетке копий исходных вирусных геномов, которые используются при сборке вирионов. Способ репликации генома зависит от типа нуклеиновой кислоты вируса, наличия вирусоспецифических или клеточных полимераз, а также от способности вирусов индуцировать образование полимераз в клетке.
Механизм репликации отличается у вирусов, имеющих:
1) двунитевую ДНК;
2) однонитевую ДНК;
3) плюс-однонитевую РНК;
4) минус-одноните-вую РНК;
5) двунитевую РНК;
6) идентичные плюс-нитевые РНК (ретровирусы).
1. Двунитевые ЛНК-вирусы. Репликация двунитевых вирусных ДНК происходит обычным полуконсервативным механизмом: после рас-
плетения нитей ДНК к ним комплементарно достраиваются новые нити. Каждая вновь синтезированная молекула ДНК состоит из одной родительской и одной вновь синтезированной нити. К этим вирусам относится большая
группа вирусов, которые содержат двунитевую ДНК в линейной (например, герпесвирусы, аденовирусы и поксвирусы) или в кольцевой форме, как папилломавирусы. У всех вирусов, кроме поксвирусов, транскрипция вирусного генома происходит в ядре.
Уникальный механизм репликации характерен для гепаднавирусов (вируса гепатита В). Геном гепаднавирусов представлен дву-нитевой кольцевой ДНК, одна нить которой короче (неполная плюс-нить) другой нити. Первоначально достраивается (рис. 3.7). Затем полная двунитевая ДНК с помощью клеточной ДНК-зависимой РНК-полимеразы транскрибируется с образованием небольших молекул иРНК и полной однонитевой плюс-РНК. Последняя называется прегеномной РНК; она является матрицей для репликации генома вируса. Синтезированные иРНК участвуют в процессе трансляции белков, в том числе вирусной РНК-зависимой ДНК-полимеразы (обратной транскриптазы). С помощью этого фермента мигрирующая в цитоплазму прегеномная РНК обратно транскрибируется в минус-нить ДНК, которая, в свою очередь, служит матрицей для синтеза плюс-нити ДНК. Этот процесс заканчивается образованием двунитевой ДНК, содержащей неполную плюс-нить ДНК.
2.Однонитевые ДНК-вирусы. Единственными представителями однонитевых ДНК-вирусов являются парвовирусы. Парвовирусы
используют клеточные ДНК-полимеразы для создания двунитевого вирусного генома, так называемой репликативной формы послед
него. При этом на исходной вирусной ДНК (плюс-нить) комплементарно синтезируется минус-нить ДНК, служащая матрицей для синтеза плюс-нити ДНК нового вириона. Параллельно синтезируется иРНК, происходит трансляция вирусных пептидов.
3.Плюс-однонитевые РНК-вирусы. Эти вирусы включают большую группу вирусов — пикорнавирусы, флавивирусы, тогавирусы (рис.3.8), у которых геномная плюс-нить РНК выполняет функцию иРНК. Например, РНК
полиовирусов после проникновения в клетку связывается с рибосомами, работая как иРНК, и на ее основе синтезируется большой полипептид, который расщепляется на фрагменты:
РНК-зависимую РНК-полимеразу, вирусные протеазы и капсидные белки. Полимераза на основе геномной плюс-нити РНК синтезирует минус-нить РНК; формируется временно двойная РНК, названная промежуточным
репликативным звеном. Это промежуточное репликативное звено состоит из полной плюс-нити РНК и многочисленных частично завершенных минус-нитей. Когда образованы все минус-нити, они используются как шаблоны
для синтеза новых плюс-нитей РНК. Этот механизм используется как для размножения геномной РНК вируса, так и для синтеза большого количества вирусных белков.
4.Минус-однонитевые РНК-вирусы. Минус -однонитевые РНК-вирусы (рабдовирусы, парамиксовирусы, ортомиксовирусы) имеют в своем составе РНК-зависимую РНК-полимеразу. Проникшая в клетку геномная минус-
нить РНК трансформируется вирусной РНК-зависимой РНК-полимеразой в неполные и полные плюс-нити РНК. Неполные копии выполняют роль иРНК для синтеза вирусных белков. Полные копии являются матрицей (промежуточная стадия) для синтеза минус-нитей геномной РНК потомства
5.Двунитевые РНК-вирусы. Механизм репликации этих вирусов (реовирусов и ротави-русов) сходен с репликацией минус-однонитевых РНК-вирусов. Отличие состоит в том, что образовавшиеся в процессе транскрипции плюс-нити функционируют не только как иРНК, но и участвуют в репликации: они являются матрицами для синтеза минус-нитей
РНК. Последние в комплексе с плюс-нитями РНК образуют геномные двунитевые РНК вирионов. Репликация вирусных нуклеиновых кислот этих вирусов происходит в цитоплазме клеток.
6. Ретровирусы (плюс-нитевые диплоидные РНК-содержащие вирусы). Обратная транс-криптаза ретровирусов синтезирует (на матрице РНК-вируса) минус-нить ДНК, с которой копируется плюс-нить ДНК с образованием двойной нити ДНК, замкнутой в кольцо (рис. 3.10). Далее двойная нить ДНК интегрирует с хромосомой клетки, образуя провирус. Многочисленные вирионные РНК образуются в результате транскрипции одной из нитей интегрированной ДНК при участии клеточной ДНК-зависимой РНК-полимеразы.
Формирование вирусов.Вирионы формируются путем самосборки: составные части вириона транспортируются в места сборки вируса — участки ядра или цитоплазмы клетки. Соединение компонентов вириона обусловлено наличием гидрофобных, ионных, водородных связей и стерического соответствия.
Существуют следующие общие принципы сборки вирусов:
Формирование вирусов— многоступенчатый процесс с образованием промежуточных форм, отличающихся от зрелых вирионов по
составу полипептидов.
□ Сборка просто устроенных вирусов заключается во взаимодействии вирусных нуклеиновых кислот с капсидными белками и в образовании нуклеокапсидов.
□ У сложноустроенных вирусов сначала формируются нуклеокапсиды, которые взаимодействуют с модифицированными мембранами клеток (будущей липопротеиновой оболочкой вируса).
Причем сборка вирусов, реплицирующихся в ядре клетки, происходит с участием мембраны ядра, а сборка вирусов, репликация которых идет в цитоплазме, осуществляется с
участием мембран эндоплазматической сети или плазматической мембраны, куда встраиваются гликопротеины и другие белки оболочки вируса.
□ У ряда сложноустроенных вирусов минус-нитевых РНК-вирусов (ортомиксовирусов, парамиксовирусов) в сборку вовлекается так называемый матриксный белок (М-белок), который расположен под модифицированной клеточной ембраной. Обладая гидрофобными свойствами, он выполняет роль посредника между нуклеокапсидом и вирусной липопротеиновой оболочкой.
□ Сложноустроенные вирусы в процессе формирования включают в свой состав некоторые компоненты клетки хозяина, например липиды и углеводы.
Выход вирусов из клетки.Полный цикл репродукции вирусов завершается через 5—6 ч (вирус гриппа и др.) или через несколько суток (гепатовирусы, вирус кори и др.). Процесс репродукции вирусов заканчивается выходом их из клетки, который происходит взрывным путем или почкованием, экзоцитозом.
□ Взрывной путь: из погибающей клетки одновременно выходит большое количество вирионов. По взрывному пути выходят из клетки просто устроенные вирусы, не имеющие липопротеиновой оболочки.
□ Почкование, экзоцшпт присущи вирусам, имеющим липопротеиновую оболочку, которая является производной от клеточных мембран. Сначала образовавшийся нуклеокапсид или сердцевина вириона транспортируется к клеточным мембранам, в которые уже встроены вирусоспецифические белки. Затем в области контакта нуклеокапсида или сердцевины вириона с клеточной мембраной начинается выпячивание этих участков. Сформировавшаяся почка отделяется от клетки в виде сложно устроенного вируса. При этом клетка способна длительно сохранять жизнеспособность и продуцировать вирусное потомство.
Почкование вирусов, формирующихся в цитоплазме, может происходить либо через плазматическую мембрану (например, парамиксовирусы, тогавирусы), либо через мембраны эндоплазматической сети с последующим их выходом на поверхность клетки (например, буньявирусы).
Вирусы, формирующиеся в ядре клетки (например, герпесвирусы), почкуются в перинуклеарное пространство через модифицированную ядерную мембрану, приобретая таким образом липопротеиновую оболочку. Затем они транспортируются в составе цитоплазма-тических везикул на поверхность клетки.
Вскоре после обнажения вирусного генома происходит уменьшение или исчезновение инфекционности, поскольку вирионов как таковых уже не существует, а инфекционность нуклеиновой кислоты намного меньше инфекционности полных вирионов. Данное явление было названо эклипсом (затмением), а фаза вирусной инфекции с момента адсорбции и до появления в клетке новых инфекционных вирионов — эклипс-фазой, или латентным периодом. При высокой множественности заражения клеток некоторая часть вирионов сохраняет инфекционные свойства в течение всей эклипс-фазы, что зачастую затрудняет определение ее продолжительности.
В течение этого периода происходят все существенные процессы вирусной репродукции, приводящие к образованию вирусных компонентов и формированию нового потомства вирионов. С появлением в клетке первых вновь синтезированных вирионов завершается эклипс-период размножения вируса.
Процессы, происходящие в эклипс-фазе, схематично показаны на рисунке, где в качестве примера представлены основные этапы репликации аденовирусов.
Из представленной схемы видно, что после прикрепления вирион входит в клетку и частично или полностью лишается оболочки, обнажая вирусный геном. Некоторые ранние вирусные гены транскрибируются с образованием мРНК, которые могут затем изменяться различными путями, включая сплайсинг. Ранние мРНК транслируются с образованием трех главных продуктов: белков, которые подавляют или останавливают синтез белков и нуклеиновых кислот; белков, которые регулируют экспрессию вирусного генома; ферментов, необходимых для репликации вирусных нуклеиновых кислот. Вслед за репликацией вирусной нуклеиновой кислоты транскрибируются поздние вирусные гены, продуктами трансляции которых являются в основном структурные вирусные белки, используемые при сборке новых вирионов. Некоторые из них претерпевают посттрансляционные изменения перед включением в структуру вирионов. В каждой инфицированной клетке образуются сотни и тысячи вирионов, которые, освобождаясь, заражают другие клетки.
У большинства ДНК-вирусов транскрипция и репликация ДНК происходит в ядре клетки, с использованием клеточной РНК-полимеразы II и других клеточных ферментов. Большинство РНК-вирусов размножается в цитоплазме, где отсутствуют ферменты, копирующие РНК на РНК-матрице, поэтому вирусный геном должен сам по себе функционировать как мРНК или вирус должен нести свою РНК-полимеразу, чтобы транскрибировать РНК на РНК-геноме.
Синтез вирусных белков происходит только в цитоплазме клетки. В инфицированных клетках вирусные нуклеиновые кислоты и вирусспецифические белки синтезируются в значительно большем количестве, чем включаются в вирионы. Избыточный синтез структурных компонентов при репродукции вирусов является своеобразной платой за паразитизм на генетическом уровне.
Потомство вирионов проявляется спустя определенный период после того, как начал осуществляться синтез составляющих их компонентов. Продолжительность периода от момента инфицирования до появления дочерних вирионов довольно вариабельна и отражает глубокое различие между разными вирусами.
Латентный период мелких вирусов, не имеющих оболочки и содержащих односпиральную РНК, длится примерно 2 часа и является наиболее коротким среди всех известных вирусов животных. ДНК-содержащие вирусы имеют более продолжительный латентный период. Длительный латентный период отмечен также у реовирусов и вируса саркомы Рауса. У простых мелких РНК-содержащих вирусов быстрее протекает не только латентный период, но и весь цикл размножения вируса. Вирусы, геном которых выполняет функции мРНК, имеют более короткий репликативный цикл, чем вирусы, у которых функции мРНК выполняет не вирионная РНК, а комплементарная ей минус-нить.
Размножение ДНК-содержащих вирусов может продолжаться даже через 24 часа после заражения. Причины, определяющие длительность цикла размножения разных вирусов, не выяснены. Возможно, что это связано с более сложной последовательностью событий при репродукции крупных, сложно устроенных вирусов.
Укорочение латентного периода при высокой множественности заражения было обнаружено у различных РНК и ДНК вирусов и, возможно, связано с ускорением процесса образования ферментов, участвующих в синтезе нуклеиновой кислоты. При высокой множественности заражения не исключается также роль капсидных белков в ускорении начала цикла репродукции.
Число инфекционных вирусных частиц, образуемых в одной клетке, зависит от типа вируса, и количество их варьирует очень широко. Интересно, что на долю вирусспецифических продуктов приходится от 0,1 до 5% массы клетки животного.
Биосинтез белка – важная часть пластического обмена всех клеток. Рассматривает данный процесс наука биология. В результате образуются специфичные вещества, характерные для данного организма. Происходит воспроизведение наследственной информации.
Последовательность процессов биосинтеза белка
Образование белка является многоступенчатым процессом.
Чтобы запустить реакции образования вещества, осуществляется целый ряд последовательных событий:
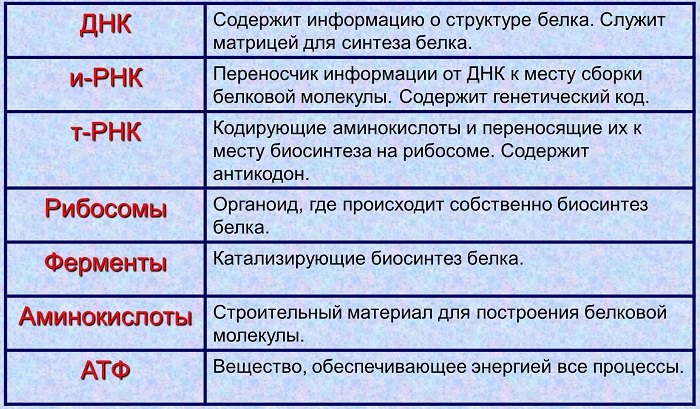
Транскрипция - это реакции переписывания наследственной информации с макромолекулы ДНК на матричную РНК. Ее называют также информационной. Краткое обозначение: м-РНК, и-РНК. Процесс протекает в ядре клетки.
Перемещение и-РНК к месту синтеза белка.
Трансляция - это перенос информации о чередовании нуклеотидов м-РНК на макромолекулу белка. Процесс идёт вне ядра.
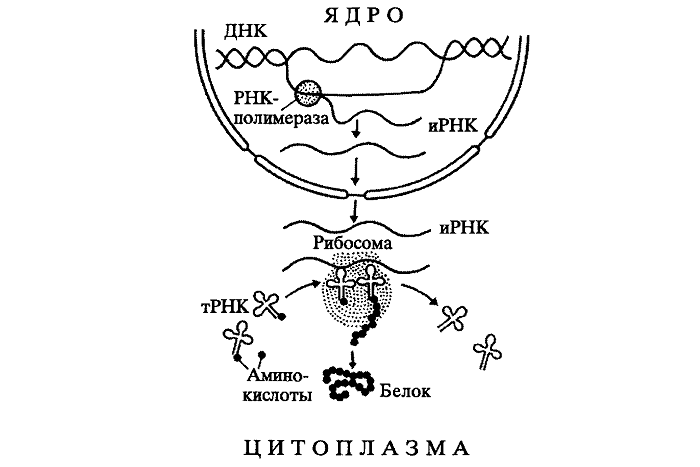
Где происходит синтез белка
Образование высокомолекулярного соединения протекает в цитоплазме. Именно здесь находятся органоиды, на которых осуществляется данный процесс. Рибосома представляет собой две части: малую и большую. Чтобы биосинтез белка начался, необходимо доставить информацию из ядра в цитоплазму.

Ядро эукариот хранит информацию о первичной структуре природных полимеров. Её называют наследственной. Эта важная информация должна быть без искажения перенесена к месту синтеза белка.
С этой целью в ядре идут матричные реакции. На одной из цепей ДНК синтезируется и-РНК. Именно она является посредником между двумя частями клетки.
Этапы биосинтеза белка
Процесс протекает в ядре. ДНК образована большим количеством нуклеотидов. Это единица макромолекулы. Она включает в свой состав 3 компонента:
углевод, представленный пентозой – дезоксирибозой;
минеральную кислоту – фосфорную;
органическое соединение, относящееся к классу азотистых оснований.
В составе ДНК могут содержаться 4 разных основания. Они имеют краткое обозначение, по первой букве названия:
Именно этими основаниями и отличаются нуклеотиды. Чередование 3 нуклеотидов образует триплет. Один триплет соответствует одной аминокислоте. Вопрос соответствия аминокислот триплетам изучен и указан в таблице генетического кода.
Последовательность триплетов в молекуле дезоксирибонуклеиновой кислоты, отвечающей за синтез одного белка, называют геном. Между разными генами расположены триплеты, которые не соответствуют аминокислотам. Их называют стоп-кодонами. Они служат сигналом начала и окончания гена.
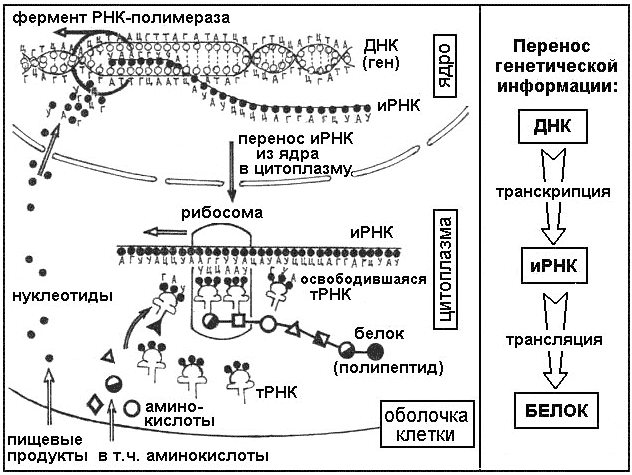
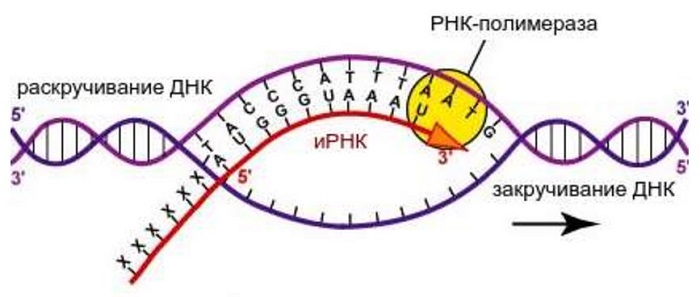
Для осуществления транскрипции, участок макромолекулы ДНК раскручивается. Он выполняет роль матрицы. На нём выстраивается и-РНК. Осуществляется синтез по принципу соответствия. Еще его называют комплементарностью.
РНК также имеет нуклеотидное строение. Вместо дезоксирибозы присутствует углевод рибоза. Содержится остаток ортофосфорной кислоты. Третьим компонентом является азотистое основание. Три основания одинаковые – А, Г, Ц в ДНК и РНК. Четвертое основание рибонуклеиновой кислоты – урацил (У).
Комплементарными основаниями являются: Т – А, А – У, Г – Ц, Ц – Г. В парах комплементарных оснований первое соответствует ДНК, второе – РНК. Таким образом, на макромолекуле ДНК по принципу соответствия выстраивается и-РНК. В дальнейшем цепь РНК транспортируется через ядерную мембрану к месту синтеза белка.
Процесс идет на органоидах – рибосомах. Они нанизываются на цепь и-РНК, передвигаются по ней не плавно, а прерывисто. Располагаются таким образом, что внутри рибосомы находится полностью 1-2 триплета. На одну РНК может одновременно нанизываться большое количество рибосом.
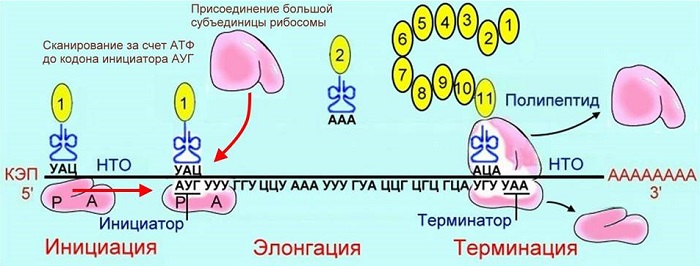
В процессе принимают участие т-РНК. Они имеют пространственную структуру, принимают форму трилистника. Верхняя часть листа, то есть молекулы, содержит антикодон. Это триплет, распознающий кодон (один триплет) и-РНК.
Каждая т-РНК транспортирует к рибосоме строго определенную аминокислоту. Если триплет-антикодон т-РНК распознает триплет-кодон и-РНК, тогда аминокислота встраивается в макромолекулу белка. Следующая т-РНК подтаскивает другую аминокислоту, снова идет процесс распознавания. В данном случае также идет матричный процесс сборки белка. РНК служит матрицей для синтеза белка.
Как только белковая молекула синтезирована, она освобождается от рибосомы. Правильное чередование аминокислот в макромолекуле образует первичную структуру белковой молекулы. Она является определяющей, поэтому так важен матричный синтез белков. Другие структуры белковые макромолекулы приобретают самопроизвольно.
Схема биосинтеза белка
Процессы, ведущие к синтезу белка, можно кратко изобразить на схеме:
Первый этап – реакции, идущие в кариоплазме. Раскручивание ДНК. Транскрипция. Образование м-РНК.
Второй этап – транспорт м-РНК к рибосомам.
Третий этап – реакции, идущие в цитоплазме. Трансляция. Биосинтез белковой молекулы, протекающий при участии РНК, клеточных органоидов – рибосом.
Заключение
В реакциях матричного синтеза происходит реализация наследственной информации. В каждом организме синтезируются специфичные белковые молекулы. Они вместе с углеводами и жирами накапливаются в плодах растений. В организмах животных выполняют множество разнообразных функций.
Читайте также: