
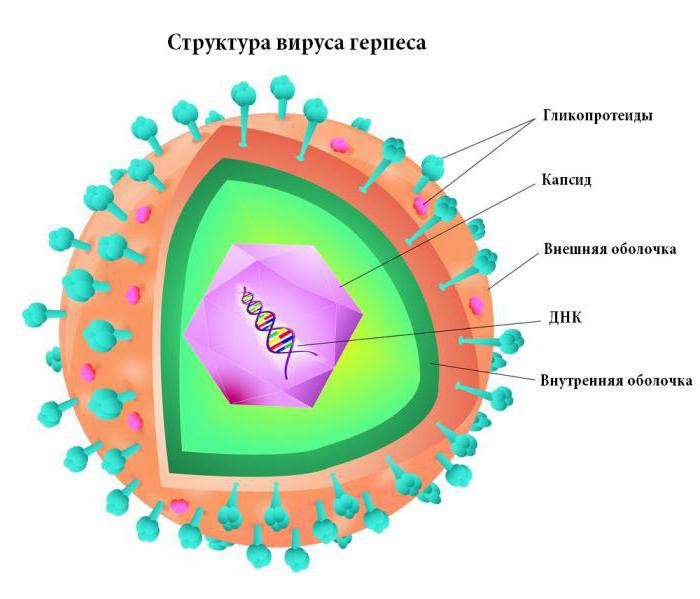
Однонитевая днк у вирусов
Продуктивный тип взаимодействия вируса с клеткой, т. е. репродукция вируса (лат. re — повторение, productio — производство), проходит в 6 стадий:
1) адсорбция вирионов на клетке;
2) проникновение вируса в клетку;
4) синтез вирусных компонентов;
5) формированиевирионов;
6) выход вирионов из клетки.
У различных вирусов эти стадии отличаются
Специфические рецепторы клеток имеют различную природу, являясь белками, липидами, углеводными компонентами белков, липидов и др. Так, рецепторами для вируса гриппа является сиаловая кислота в составе гли-копротеинов и гликолипидов (ганглиозидов) клеток дыхательных путей. Вирусы бешенства адсорбируются на ацетилхолиновых рецепторах нервной ткани, а вирусы иммунодефицита человека — на СО4-рецепторах Т-хелперов, моноцитов и дендритных клеток. На одной клетке находится от десяти до ста тысяч специфических рецепторов, поэтому на ней могут адсорбироваться десятки и сотни вирионов.
Наличие специфических рецепторов лежит в основе избирательности поражения вирусами определенных клеток, тканей и органов. Это так называемый тропизм(греч. tropos — поворот, направление). Например, вирусы, репродуцирующиеся преимущественно в клетках печени, называются гепатотропными, в нервных клетках — нейротропными, в иммунокомпетентных клетках — иммунотропными и т. д.
Проникновение вирусов в клетку.Вирусы проникают в клетку путем рецептор-зависимого эндоцитоза (виропексиса), или слияния оболочки вируса с клеточной мембраной, или же в результате сочетания этих механизмов.
2. Слияние обточки вириона с клеточной мембраной характерно только для некоторых оболочечных вирусов (парамиксовирусов, ретровиру-сов, герпесвирусов), в составе которых имеются белки слияния. Происходит точечное взаимодействие вирусного белка слияния с липидами клеточной мембраны, в результате чего вирусная липопротеиновая оболочка интегрирует с клеточной мембраной, а внутренний компонент вируса попадает в цитозоль.
В) Синтез вирусных компонентов. Синтез белков и нуклеиновых кислот вируса, который разобщен во времени и пространстве. Синтез осуществляется в разных частях клетки, поэтому такой способ размножения вирусов называется дизъюнктивным (от лат. disjunctus — разобщенный).
С)Синтез вирусных белков. В зараженной клетке вирусный геном кодирует синтез двух групп белков:
1. неструктурных белков, обслуживающих внутриклеточную репродукцию вируса на разных его этапах;
2. структурных белков, которые входят в состав вириона (геномные, связанные с геномом вируса, капсидные и су-перкапсидные белки).
К неструктурным белкамотносятся: 1) ферменты синтеза РНК или ДНК (РНК- или ДНК-полимеразы), обеспечивающие транскрипцию и репликацию вирусного генома; 2) белки-регуляторы; 3) предшественники вирусных белков, отличающиеся своей нестабильностью в результате быстрого нарезания на структурные белки; 4) ферменты, модифицирующие вирусные белки, например, протеиназы и протеинкиназы.
I. ДНК-содержашие вирусы реализуют генетическую информацию так же, как и клеточный геном, по схеме:
Причем ДНК-содержашие вирусы используют для этого процесса клеточную полимеразу (вирусы, геномы которых транскрибируются в ядре клетки — аденовирусы, па-повавирусы, герпесвирусы) или собственную РНК-полимеразу (вирусы, геномы которых транскрибируются в цитоплазме, например поксвирусы).
II. Плюс-нитевые РНК-содержашие вирусы(например, пикорнавирусы, флавивирусы, тогавирусы) имеют геном, выполняющий функцию иРНК; он распознается и транслируется рибосомами. Синтез белков у этих вирусов осуществляется без акта транскрипции по схеме:
геномная РНК вируса-> трансляция белка вируса.
III. Геном минус-однонитевых РНК-содержаших вирусов (ортомиксовирусов, парамиксовирусов, рабдовирусов) и двунитевых (реовирусов) служит матрицей, с которой транскрибируется иРНК, при участии РНК-полимеразы, связанной с нуклеиновой кислотой вируса. Синтез белка у них происходит по схеме:
IV. Ретровирусы(вирусы иммунодефицита человека, онкогенные ретровирусы) имеют уникальный путь передачи генетической информации. Геном ретровирусов состоит из двух идентичных молекул РНК, т. е. является диплоидным. В составе ретровирусов есть особый вирусоспецифический фермент — обратная транскриптаза, или ревертаза, с помощью которой осуществляется процесс обратной транскрипции, т. е. на матрице геномной РНК синтезируется комплементарная однонитевая ДНК (кДНК). Комплементарная нить ДНК копируется с образованием двунитевой комплементарной ДНК, которая интегрирует в клеточный геном и в его составе транскрибируется в иРНК с помощью клеточной ДНК-зависимой РНК-полимеразы. Синтез белков для этих вирусов осуществляется по схеме:
Репликация вирусных геномов,т. е. синтез вирусных нуклеиновых кислот, приводит к накоплению в клетке копий исходных вирусных геномов, которые используются при сборке вирионов. Способ репликации генома зависит от типа нуклеиновой кислоты вируса, наличия вирусоспецифических или клеточных полимераз, а также от способности вирусов индуцировать образование полимераз в клетке.
Механизм репликации отличается у вирусов, имеющих:
1) двунитевую ДНК;
2) однонитевую ДНК;
3) плюс-однонитевую РНК;
4) минус-одноните-вую РНК;
5) двунитевую РНК;
6) идентичные плюс-нитевые РНК (ретровирусы).
1. Двунитевые ЛНК-вирусы. Репликация двунитевых вирусных ДНК происходит обычным полуконсервативным механизмом: после рас-
плетения нитей ДНК к ним комплементарно достраиваются новые нити. Каждая вновь синтезированная молекула ДНК состоит из одной родительской и одной вновь синтезированной нити. К этим вирусам относится большая
группа вирусов, которые содержат двунитевую ДНК в линейной (например, герпесвирусы, аденовирусы и поксвирусы) или в кольцевой форме, как папилломавирусы. У всех вирусов, кроме поксвирусов, транскрипция вирусного генома происходит в ядре.
Уникальный механизм репликации характерен для гепаднавирусов (вируса гепатита В). Геном гепаднавирусов представлен дву-нитевой кольцевой ДНК, одна нить которой короче (неполная плюс-нить) другой нити. Первоначально достраивается (рис. 3.7). Затем полная двунитевая ДНК с помощью клеточной ДНК-зависимой РНК-полимеразы транскрибируется с образованием небольших молекул иРНК и полной однонитевой плюс-РНК. Последняя называется прегеномной РНК; она является матрицей для репликации генома вируса. Синтезированные иРНК участвуют в процессе трансляции белков, в том числе вирусной РНК-зависимой ДНК-полимеразы (обратной транскриптазы). С помощью этого фермента мигрирующая в цитоплазму прегеномная РНК обратно транскрибируется в минус-нить ДНК, которая, в свою очередь, служит матрицей для синтеза плюс-нити ДНК. Этот процесс заканчивается образованием двунитевой ДНК, содержащей неполную плюс-нить ДНК.
2.Однонитевые ДНК-вирусы. Единственными представителями однонитевых ДНК-вирусов являются парвовирусы. Парвовирусы
используют клеточные ДНК-полимеразы для создания двунитевого вирусного генома, так называемой репликативной формы послед
него. При этом на исходной вирусной ДНК (плюс-нить) комплементарно синтезируется минус-нить ДНК, служащая матрицей для синтеза плюс-нити ДНК нового вириона. Параллельно синтезируется иРНК, происходит трансляция вирусных пептидов.
3.Плюс-однонитевые РНК-вирусы. Эти вирусы включают большую группу вирусов — пикорнавирусы, флавивирусы, тогавирусы (рис.3.8), у которых геномная плюс-нить РНК выполняет функцию иРНК. Например, РНК
полиовирусов после проникновения в клетку связывается с рибосомами, работая как иРНК, и на ее основе синтезируется большой полипептид, который расщепляется на фрагменты:
РНК-зависимую РНК-полимеразу, вирусные протеазы и капсидные белки. Полимераза на основе геномной плюс-нити РНК синтезирует минус-нить РНК; формируется временно двойная РНК, названная промежуточным
репликативным звеном. Это промежуточное репликативное звено состоит из полной плюс-нити РНК и многочисленных частично завершенных минус-нитей. Когда образованы все минус-нити, они используются как шаблоны
для синтеза новых плюс-нитей РНК. Этот механизм используется как для размножения геномной РНК вируса, так и для синтеза большого количества вирусных белков.
4.Минус-однонитевые РНК-вирусы. Минус -однонитевые РНК-вирусы (рабдовирусы, парамиксовирусы, ортомиксовирусы) имеют в своем составе РНК-зависимую РНК-полимеразу. Проникшая в клетку геномная минус-
нить РНК трансформируется вирусной РНК-зависимой РНК-полимеразой в неполные и полные плюс-нити РНК. Неполные копии выполняют роль иРНК для синтеза вирусных белков. Полные копии являются матрицей (промежуточная стадия) для синтеза минус-нитей геномной РНК потомства
5.Двунитевые РНК-вирусы. Механизм репликации этих вирусов (реовирусов и ротави-русов) сходен с репликацией минус-однонитевых РНК-вирусов. Отличие состоит в том, что образовавшиеся в процессе транскрипции плюс-нити функционируют не только как иРНК, но и участвуют в репликации: они являются матрицами для синтеза минус-нитей
РНК. Последние в комплексе с плюс-нитями РНК образуют геномные двунитевые РНК вирионов. Репликация вирусных нуклеиновых кислот этих вирусов происходит в цитоплазме клеток.
6. Ретровирусы (плюс-нитевые диплоидные РНК-содержащие вирусы). Обратная транс-криптаза ретровирусов синтезирует (на матрице РНК-вируса) минус-нить ДНК, с которой копируется плюс-нить ДНК с образованием двойной нити ДНК, замкнутой в кольцо (рис. 3.10). Далее двойная нить ДНК интегрирует с хромосомой клетки, образуя провирус. Многочисленные вирионные РНК образуются в результате транскрипции одной из нитей интегрированной ДНК при участии клеточной ДНК-зависимой РНК-полимеразы.
Формирование вирусов.Вирионы формируются путем самосборки: составные части вириона транспортируются в места сборки вируса — участки ядра или цитоплазмы клетки. Соединение компонентов вириона обусловлено наличием гидрофобных, ионных, водородных связей и стерического соответствия.
Существуют следующие общие принципы сборки вирусов:
Формирование вирусов— многоступенчатый процесс с образованием промежуточных форм, отличающихся от зрелых вирионов по
составу полипептидов.
□ Сборка просто устроенных вирусов заключается во взаимодействии вирусных нуклеиновых кислот с капсидными белками и в образовании нуклеокапсидов.
□ У сложноустроенных вирусов сначала формируются нуклеокапсиды, которые взаимодействуют с модифицированными мембранами клеток (будущей липопротеиновой оболочкой вируса).
Причем сборка вирусов, реплицирующихся в ядре клетки, происходит с участием мембраны ядра, а сборка вирусов, репликация которых идет в цитоплазме, осуществляется с
участием мембран эндоплазматической сети или плазматической мембраны, куда встраиваются гликопротеины и другие белки оболочки вируса.
□ У ряда сложноустроенных вирусов минус-нитевых РНК-вирусов (ортомиксовирусов, парамиксовирусов) в сборку вовлекается так называемый матриксный белок (М-белок), который расположен под модифицированной клеточной ембраной. Обладая гидрофобными свойствами, он выполняет роль посредника между нуклеокапсидом и вирусной липопротеиновой оболочкой.
□ Сложноустроенные вирусы в процессе формирования включают в свой состав некоторые компоненты клетки хозяина, например липиды и углеводы.
Выход вирусов из клетки.Полный цикл репродукции вирусов завершается через 5—6 ч (вирус гриппа и др.) или через несколько суток (гепатовирусы, вирус кори и др.). Процесс репродукции вирусов заканчивается выходом их из клетки, который происходит взрывным путем или почкованием, экзоцитозом.
□ Взрывной путь: из погибающей клетки одновременно выходит большое количество вирионов. По взрывному пути выходят из клетки просто устроенные вирусы, не имеющие липопротеиновой оболочки.
□ Почкование, экзоцшпт присущи вирусам, имеющим липопротеиновую оболочку, которая является производной от клеточных мембран. Сначала образовавшийся нуклеокапсид или сердцевина вириона транспортируется к клеточным мембранам, в которые уже встроены вирусоспецифические белки. Затем в области контакта нуклеокапсида или сердцевины вириона с клеточной мембраной начинается выпячивание этих участков. Сформировавшаяся почка отделяется от клетки в виде сложно устроенного вируса. При этом клетка способна длительно сохранять жизнеспособность и продуцировать вирусное потомство.
Почкование вирусов, формирующихся в цитоплазме, может происходить либо через плазматическую мембрану (например, парамиксовирусы, тогавирусы), либо через мембраны эндоплазматической сети с последующим их выходом на поверхность клетки (например, буньявирусы).
Вирусы, формирующиеся в ядре клетки (например, герпесвирусы), почкуются в перинуклеарное пространство через модифицированную ядерную мембрану, приобретая таким образом липопротеиновую оболочку. Затем они транспортируются в составе цитоплазма-тических везикул на поверхность клетки.
Одни вирусы способны интегрироваться в геном клетки-мишени и таким образом оставаться во всех дочерних клетках, которые будут в будущем получены после ее деления. К таким вирусам относятся гаммаретровирусы и лентивирусы. Другие делать этого не умеют (например, адено- и аденоассоциированные вирусы). Но для производства белков и репликации (размножения) все они используют клетку и ее синтетический аппарат.
Как это работает?
Для того чтобы вирус мог проникнуть в клетку, белки его оболочки должны связаться с мембранными белками клетки-мишени. Важно отметить, что проникает вирус только в те клетки, которые могут в дальнейшем помочь его репликации. Вирус ВИЧ живет в клетках иммунной системы, вирус гепатита С — в клетках печени. Есть особые вирусы, которые поражают только растения или даже только бактерии.
Содержащие неактивный вирус и оставшиеся в живых клетки иногда сохраняют возможность нормального функционирования. В этом случае клетки могут быть заражены, но вирус проявит себя спустя длительный период времени. Так устроен герпес.
В зависимости от того, каким типом нуклеиновой кислоты представлен генетический материал, выделяют ДНК-содержащие вирусы и РНК-содержащие вирусы. И тут стоит остановиться на классификации.
Типы вирусов. Коротко о главном
Современная типология вирусов содержит 7 классов и была предложена Дэвидом Балтимором еще в 1971 году. С тех пор, впрочем, она была уточнена и расширена, в том числе советскими учеными. И выглядит в настоящее время таким образом:
- Класс I
Вирусы, содержащие двухцепочечную ДНК
Описание
Для репликации вирусу необходимо попасть в ядро клетки-мишени и воспользоваться ее ДНК-полимеразой. Иногда вирус вызывает незапланированное деление самое клетки, то есть становится онкогенным. Эти вирусы хорошо изучены.
Пример: Вирус герпеса, адено- и папилломавирусы
Вирусы, содержащие одноцепочечную ДНК
Описание
Попадая в ядро клетки, вирусы образуют двухцепочечную ДНК, после чего реплицируются так же, как вирусы класса I.
Пример: Парво- и цирковирусы
Вирусы, в которых РНК способна к репликации (редупликации)
Описание
Вирусы этого класса могут размножаться в цитоплазме клетки, им не нужна молекула ДНК. Каждый ген, находящийся в РНК вируса, кодирует только один вирусный белок.
Пример: Бирна- и реовирусы
Вирусы, содержащие одноцепочечную (+) РНК
Описание
Из геномной (+) РНК на рибосомах хозяина создаются вирусные белки. В одном фрагменте РНК могут быть закодированы разные белки, что увеличивает сложность вируса без удлинения генов.
Пример: Пикорнавирусы (полиомиелит, гепатит А) и коронавирусы
Вирусы, содержащие одноцепочечную (–) РНК
Описание
(–) РНК этих вирусов предварительно должна быть транскрибирована в (+) РНК вирусными РНК-полимеразами, после чего может начаться синтез вирусных белков. Вирусы этого класса делятся еще на две группы, в зависимости от их генома и места его репликации (цитоплазма или ядро).
Пример: Филовирусы, аренавирусы (геморрагическая лихорадка Ласса), ортомиксовирусы (вирусы гриппа) и так далее.
Вирусы, содержащие одноцепочечную (+) РНК, реплицирующиеся через стадию ДНК
Описание
Такие вирусы используют фермент обратную транскриптазу для превращения (+) РНК в ДНК, которая встраивается в геном хозяина ферментом интегразой. Дальнейшая репликация происходит при помощи полимераз клетки хозяина.
Пример: Ретровирусы (в том числе ВИЧ)
Вирусы, содержащие двухцепочечную ДНК, реплицирующиеся через стадию одноцепочечной РНК
Описание
Молекула ДНК замкнута в кольцо и является матрицей для синтеза мРНК и дополнительных молекул РНК, которые используются при репликации вирусного генома обратными транскриптазами.
Пример: Колимовирусы (вызывают инфекции растений) и гепаднавирусы (например, гепатит В)
Вакцинация и лечение
Как правило, организмы умеют бороться с паразитирующими на них вирусами. На примере млекопитающих и человека мы обычно говорим о главном инструменте — врожденном иммунитете.
Впрочем, наиболее эффективен этот вид защиты в отношении бактериальных инфекций и не может обеспечить продолжительную и надежную защиту, особенно от инфекций вирусных.
Именно поэтому огромное значение имеет приобретенный иммунитет, в результате которого клетки иммунной системы обучаются вырабатывать специфические к вирусу антитела, способные уничтожать как саму вирусную частицу, так и зараженные ею клетки.
Еще одна врожденная система борьбы с вирусными инфекциями — внутриклеточная. Как правило, клетка способна распознать чужеродную РНК в своей цитоплазме, куда ее сперва и доставляют многие вирусы, и имеет специальные комплексы для ее деградации. Но часть вирусов научились обходить и эту ловушку. К примеру, ротавирусы, которые даже внутри клетки сохраняют капсид с геномной РНК.
Миссия: уничтожить
Основная сложность в лечении вирусных заболеваний заключается в том, что они используют естественные функции клеток-мишеней для своего размножения, поэтому ученым зачастую оказывается не так-то просто придумать препарат, который будет токсичен для вируса и безопасен для самой клетки. Если такой безопасности достичь не удастся, лекарство будет иметь слишком много побочных эффектов, повреждающих сам организм, что окажется нецелесообразно для использования.
По принципу действия противовирусные препараты подразделяются на две группы: стимулирующие иммунную систему атаковать вирусы (например, за счет индукции синтеза белков-интерферонов) и атакующие вирусы напрямую. Препараты второй группы различаются по этапу жизненного цикла вируса, на котором они активны: это препараты, препятствующие проникновению вируса в клетку, препятствующие размножению вируса внутри клетки и препятствующие выходу копий вируса из клетки.
Чтобы помешать проникновению вируса, препарат должен заблокировать рецептор на клетке, с которым связывается вирусная частица. Так работает, например, ибализумаб — зарегистрированный в США новый препарат против ВИЧ, о котором мы недавно писали.
Еще один класс противовирусных препаратов блокирует ферменты, необходимые для создания и модификаций белков вируса. Такие лекарства называют протеазными ингибиторами.
Вместо заключения: а могут ли вирусы приносить пользу?
Безусловно, да. Несмотря на то, что вирусы ассоциируются у большинства людей с однозначным вредом, они могут приносить и пользу — если речь идет о так называемых вирусных векторах и терапевтических подходах на их основе.
Исследователи давно научились помещать в белковую оболочку вируса интересующие их нуклеиновые кислоты, чтобы доставлять нужный ген в клетки, а также убирать те гены, которые делают вирус опасным для организма.
Это позволило сделать возможной генную терапию, помогающую бороться с заболеваниями, вызванными известными генетическими мутациями. Создание вирусных векторов — достаточно непростая задача, к тому же ограниченная свойствами самих вирусных частиц: количеством помещающейся генетической информации, местом ее вставки, стабильностью. Кроме того, вирусный вектор, используемый в медицине, не должен вызывать иммунного ответа или критично влиять на жизнедеятельность клетки. Тем не менее эти сложности решаются, поэтому уже одобрен ряд вполне успешных и безопасных генных терапий. А в качестве основы для вирусных векторов чаще всего используются ретро-, ленти-, адено- и аденоассоциированные вирусы.
Продуктивный тип взаимодействия вируса с клеткой - репродукция вируса проходит
• 1) адсорбция вирионов на клетке;
• 2) проникновение вируса в клетку;
• 3) “раздевание” и высвобождение вирусного генома (депротеинизация вируса);
• 4) синтез вирусных компонентов;
• 5) формирование вирусов;
• 6) выход вирионов из клетки.
Репродукция вирусов
• Механизм репродукции отличается у вирусов, имеющих: 1) двунитевую ДНК; 2) однонитевую ДНК; 3) плюс однонитевую РНК; 4) минус однонитевую РНК; 5) двунитевую РНК; 6) идентичные плюс нитевые РНК (ретровирусы).
• Двунитевые ДНК-вирусы – вирусы, содержащие двунитевую ДНК в линейной (например, герпесвирусы, аденовирусы и поксвирусы) или в кольцевой форме (как паповавирусы). Репликация двунитевых вирусных ДНК проходит обычным полуконсервативным механизмом: после расплетения нитей ДНК к ним комплементарно достраиваются новые нити. У всех вирусов, кроме поксвирусов транскрипция вирусного генома происходит в ядре.
Репродукция вирусов
• Плюс однонитевые РНК-вирусы . Эти вирусы включают большую группу вирусов (пикорнавирусы, флавивирусы, тогавирусы), у которых геномная плюс- нить РНК выполняет функцию иРНК.
• Минус однонитевые РНК–вирусы (рабдовирусы, парамиксовирусы, ортомиксовирусы) имеют в своем составе РНК-зависимую РНК-полимеразу. Проникшая в клетку геномная минус нить РНК трансформируется вирусной РНК-зависимой РНК-полимеразой в неполные и полные плюс нити РНК. Неполные копии выполняют роль иРНК для синтеза вирусных белков. Полные копии являются матрицей (промежуточная
стадия) для синтеза минус-нитей геном ной РНК

Однонитевые ДНК-вирусы. Единственными представителями однонитевых ДНК-вирусов являются парвовирусы
Поглощенный вирус поставляет геном в ядро клетки. Парвовирусы используют клеточные ДНК-полимеразы для создания двунитевого вирусного генома, так называемой репликативной формы последнего. При этом на исходной вирусной ДНК (плюс нить) комплементарно синтезируется минус нить ДНК, служащая матрицей в синтезе плюс нити ДНК для новых поколений вирусов. Параллельно синтезируется иРНК, происходит трансляция вирусных белков, которые возвращаются в ядро, где собираются вирионы.

Плюс однонитевые РНК-вирусы. Эти вирусы включают большую группу вирусов (пикорнавирусы, флавивирусы, тогавирусы), у которых геномная плюс-нить РНК выполняет функцию иРНК.
Вирус (1), после эндоцитоза, освобождает в цитоплазме (2) геномную плюс РНК, которая как иРНК, связывается с рибосомами (3): транслируется полипротеин (4), который расщепляется на 4 структурных белка (NSP 1-4), включая РНК-зависимую РНК-полимеразу. Эта полимераза транскрибирует геномную плюс РНК в минус нить РНК (матрицу), на которой (5) синтезируются копии РНК двух размеров: полная плюс нить геномной РНК; неполная нить иРНК, кодирующая С-белок капсида (6) и гликопротеины оболочки E1-3. Гликопротеины синтезируются на рибосомах, связанных с мембранами эндоплазматического ретикулума, затем включаются в мембрану и гликози- лируются. Дополнительно гликозилируясь в аппарате Гольджи (7), они встраиваются в плазмалемму. С-белок образует с геномной РНК нуклеокапсид, который взаимодействует с модифицированной плазмалеммой (8). Вирусы выходят из клетки почкованием (9).

Минус однонитевые РНК–вирусы (рабдовирусы, парамиксовирусы, ортомиксовирусы) имеют в своем составе РНК-зависимую РНК-полимеразу.
Схема репродукции парамиксовирусов
Вирус связывается гликопротеинами оболочки с поверхностью клетки и сливается с плазмалеммой (1). С геномной минус нити РНК вируса транскрибируются неполные плюс нити РНК, являющиеся иРНК (2) для отдельных белков и полная минус нить РНК – матрица для синтеза геномной минус РНК вируса (3). Нуклеокапсид связывается с матриксным белком и гликопротеин- модифицированной плазмалеммой. Выход вирионов – почкованием (4).

Ретровирусы (плюс-нитевые диплоидные РНК-вирусы).
Схема репродукции вируса иммунодефицита человека (ВИЧ)
ВИЧ связывается гликопротеином gp120
(1) с рецептором CD4 T-хелперов и других клеток. После слияния оболочки ВИЧ с плазмалеммой клетки в цитоплазме освобождается геномная РНК и обратная транскриптаза вируса, которая на матрице геномной РНК синтезирует комплементарную минус нить ДНК (линейная кДНК). С последней
(2) копируется плюс нить с образовани- ем двойной нити кольцевой кДНК (3), которая интегрирует с хромосомной ДНК клетки. С рекомбинантной ДНК-провиру- са (4) синтезируются геномная РНК и иРНК, которые обеспечивают синтез компонентов и сборку вирионов. Вирионы выходят их клетки почкованием (5): сердцевина вируса “одевается” в модифицированную плазмалемму клетки.
Культивирование и индикация вирусов
• Вирусы культивируют в организме лабораторных животных, в развивающихся куриных эмбрионах и культурах клеток (тканей).
• Индикацию вирусов проводят на основе следующих феноменов: цитопатогенного действия (ЦПД) вирусов, образования внутриклеточных включений, образования “бляшек”, реакции гемагглютинации, гемадсорбции или “цветной” реакции
Вирусы представляют собой форму жизни, которая погибает через некоторое время после попадания в окружающую организм среду, то есть не может существовать вне тела носителя. По факту их можно назвать паразитами внутриклеточного уровня, которые размножаются в клетках, вызывая тем самым различные заболевания. Вирусы способны поражать как РНК (рибонуклеиновую кислоту), так и ДНК (дезоксирибонуклеиновую кислоту). ДНК-содержащие вирусы признаются более консервативными с точки зрения генетики и наименее подверженными каким-либо изменениям.
Теории о происхождении
Существует несколько теорий о происхождении вирусов. Приверженцы одной теории утверждают, что зарождение вирусов происходит спонтанно и обусловлено рядом факторов. Другие считают вирусы потомками простейших форм. Однако эта теория бездоказательна и необоснованна, так как сама паразитарная суть вирусов предполагает наличие более высокоорганизованных существ, в клетках которых они могли бы существовать.
Еще одна версия происхождения вирусов предполагает преобразование более сложных форм. Эта теория говорит о вторичности простоты вируса, так она является следствием адаптации к паразитическому образу жизни. Подобное упрощение свойственно всем паразитирующим микроорганизмам. Они утрачивают способность к самостоятельному питанию, приобретая при этом склонность к быстрому размножению.
Устройство и размеры ДНК-содержащих вирусов
Самые простые вирусы имеют в составе нуклеиновую кислоту, которая выступает в качестве генетического материала как самого микроорганизма, так и его капсида, представляющего собой чехол из белка. Состав некоторых вирусов дополнен жирами и углеводами. У вирусов отсутствует часть ферментов, которая отвечает за репродуктивную функцию, поэтому размножаться они могут, только попав в клетку живого организма. Метаболизм зараженной клетки после этого перестраивается на производство вирусных, а не собственных компонентов. В каждую клетку заложена определенная генетическая информация, которая при определенных обстоятельствах может рассматриваться в качестве инструкции по синтезу конкретного вида белка внутри клетки. Зараженная клетка воспринимает данную информацию как руководство к действию.
Размеры
Что касается размеров ДНК- и РНК-содержащих вирусов, то он находится в пределах 20-300 нм. Вирусы в большинстве своем имеют меньший размер, чем бактерии. Эритроцитные клетки, например, на порядок больше вирусных. Способная к заражению, полноценная инфекционная вирусная частица вне здорового организма носит название вирион. В ядро вириона входят одна или несколько молекул нуклеиновой кислоты. Капсид представляет собой оболочку из белка, которая покрывает вирионовую нуклеиновую кислоту, обеспечивая защиту от пагубного воздействия окружающей среды. Нуклеиновая кислота, входящая в вирион, считается геномом вируса и выражается в дезоксирибонуклеиновой кислоте, или ДНК, а также рибонуклеиновой (РНК). В отличие от бактерий, у вирусов не встречается комбинация этих двух видов кислоты.
Рассмотрим основные стадии размножения ДНК-содержащих вирусов.
Размножение вирусов
Чтобы иметь возможность размножаться, вирусам необходимо внедриться в клетки носителя. Некоторые вирусы могут существовать у большого количества хозяев, у других имеется склонность к конкретным видам. На первоначальной стадии заражения вирус внедряет в клетку генетический материал в виде ДНК или РНК. Репродуктивная его функция, а также дальнейшее развитие клеток напрямую зависят от деятельности и выработки генов и белков вируса.
Для производства клеток ДНК-содержащим вирусам бывает недостаточно собственных белков, поэтому используются аналогичные вещества носителя. Спустя некоторое время после заражения в клетке остается только небольшая часть изначальных вирусов. Данная фаза называется эклипсной. Геном вируса в этот период тесно взаимодействует с носителем. Затем, спустя несколько этапов, начинается накопление во внутриклеточном пространстве потомства вируса. Это называется фазой созревания. Рассмотрим последовательность стадий размножения ДНК-содержащих вирусов.
Цикл жизни
Цикл жизни вирусов состоит из нескольких этапов, являющихся обязательными:
1. Адсорбция на клетке носителя. Это первоначальный и важный этап распознания клеток-мишеней по рецепторам. Адсорбция может происходить на клетках органов или тканей. Процесс запускает механизм дальнейшей интеграции вируса в клетку. Для связывания клеток требуется определенное количество ионов. Это необходимо для снижения электростатического отталкивания. Если проникновение в клетку не удается, вирус ищет новую мишень для интеграции и процесс повторяется. Это явление объясняет определенность в путях проникновения вируса в организм человека.
Так, например, слизистая оболочка верхних дыхательных путей обладает рецепторами к вирусу гриппа. Клетки кожи, напротив, таковых не имеют. По этой причине невозможно заразиться гриппом через кожный покров, это возможно только при вдыхании частиц вируса. Бактериальные вирусы в форме нитей или не имеющие отростков не могут прикрепляться на стенках клетки, поэтому они адсорбируются на фимфибриях. На начальном этапе адсорбция происходит за счет электростатического взаимодействия. Данная фаза обратима, так как частица вируса легко отделяется от клетки, выбранной в качестве мишени. Начиная со второй фазы, отделение не представляется возможным.
2. Следующая стадия размножения ДНК-содержащих вирусов характеризуется попаданием целого вириона или нуклеиновой кислоты, им выделяемой внутри клетки носителя. В организм животного вирус интегрируется проще, так как клетки в данном случае не снабжены оболочкой. Если у вириона есть снаружи липопротеидная мембрана, то она сталкивается при контакте с аналогичной защитой клетки носителя и вирус попадает в цитоплазму. Вирусы, проникающие в бактерии, растения и грибы интегрируются сложнее, так как в данном случае они вынуждены проходить сквозь жесткую стенку клетки. Для этого бактериофаги, к примеру, обеспечены ферментом лизоцимом, который помогает растворить твердые клеточные стенки. Примеры ДНК-содержащих вирусов рассмотрим ниже.
3. Третья стадия именуется депротеинизацией. Она характеризуется высвобождением нуклеиновой кислоты, являющейся носителем генетической информации. У некоторых вирусов, например, бактериофагов, данный процесс сочетается со второй стадией, так как белковая оболочка вириона остается за пределами клетки носителя. Вирион способен проникать в клетку путем захвата последней. При этом возникает вакуоль-фагосома, которая вбирает в себя первичные лизосомы. При этом расщепление на ферменты происходит только у белковой части вирусной клетки, а нуклеиновая кислота остается в неизменном состоянии. Именно она в дальнейшем существенно переформирует функционирование здоровой клетки, вынуждая ее производить нужные вирусу вещества. Сам вирус необходимыми для таких процедур механизмами не обеспечен. Существует такое понятие, как стратегия вирусного генома, которая подразумевает реализацию генетической информации.
4. Четвертая стадия размножения ДНК-содержащих вирусов сопровождается выработкой нужных для жизнедеятельности вируса веществ, которая осуществляется под влиянием нуклеиновой кислоты. Сначала вырабатывается ранняя мРНК, которая станет основой для белков вируса. Ранними называются молекулы, которые возникли до высвобождения нуклеиновой кислоты. Молекулы, возникшие после репликации кислоты, называют поздними. Важно понимать, что выработка молекул напрямую зависит от вида нуклеиновой кислоты конкретного вируса. ДНК-содержащие вирусы во время биосинтеза придерживаются определенной схемы, включающей конкретные этапы – ДНК-РНК-белок. Мелкие вирусы используют в процессе транскрипции РНК-полимеразы. Крупные, такие как оспяной вирус, синтезируются не в клеточном ядре, а в цитоплазме.
К ДНК-содержащим вирусам относятся вирусы гепатита В, герпес, вирусы оспы, паповавирусы, гепаднавирусы, парвовирусы.
Группы РНК-вирусов
Вирусы, содержащие РНК, подразделяются на несколько групп:
1. Первая группа устроена наиболее просто. В нее входят корона-, тога- и пикорнавирусы. Транскрипция у этих видов вируса не осуществляется, поскольку однонитчатая РНК вириона самостоятельно реализует функцию матричной кислоты, то есть представляет собой основу для выработки белков на уровне клеточных рибосом. Таким образом, схема биовыработки у них выглядит как РНК-белок. Вирусы данной группы называют также позитивно геномными или плюснитевыми.
2. Вторая группа ДНК и РНК-содержащих вирусов включает в себя минуснитевые вирусы, то есть они обладают негативным геномом. Это вирусы кори, гриппа, паротита и многие другие. В них также содержится однонитчатая РНК, но она не подходит для непосредственной трансляции. По этой причине сначала происходит перенос данных на РНК вириона, а полученная матричная кислота будет служить в дальнейшем основой для выработки белков вируса. Транскрипция в данном случае определяется зависимой от рибонуклеиновой кислоты полимеразой РНК. Данный фермент приносится вирионом, так как он отсутствует в клетке изначально. Это связано с тем, что у клетки не возникает необходимости перерабатывать РНК для получения другого РНК. Так, схема биовыработки в данном случае будет выглядеть как РНК-РНК-белок.
3. Третью группу составляют так называемые ретровирусы. Они же входят в разряд онковирусов. Биосинтез у них происходит по более сложному пути. В исходной матричной РНК однонитчатого типа на начальном этапе происходит выработка ДНК, что является уникальным явлением, аналогов которому нет в природе. Процесс находится под контролем специального фермента, а именно зависимой от РНК полимеразой ДНК. Данный фермент также носит название ревертазы или обратной транкриптазы. Молекула ДНК, полученная в результате биосинтеза, получает форму кольца и обозначается как провирус. Далее молекула внедряется в клетки хромосом носителя и транскрибируется несколько раз посредством полимеразы РНК. Созданные копии совершают следующие действия: представляют собой матрицу РНК, с помощью которой вырабатывается вирусный белок, а также вирион РНК. Схема синтеза представляется таким образом: РНК-ДНК-РНК-белок.
4. Четвертая группа образуется из вирусов, РНК которых имеет двухнитчатую форму. Их транскрипция реализуется посредством фермента вируса зависимой РНК полимеразы РНК.
5. В пятой группе выработка составляющих частицы вируса, а именно капсидных белков и нуклеиновой кислоты происходит многократно.
6. Шестая группа включает в себя вирионы, которые возникают как результат самосборки на основе множества копий белков и кислоты. С этой целью концентрация вирионов должна достичь критического значения. При этом компоненты частицы вируса вырабатываются отдельно друг от друга в разных областях клетки. Сложные вирусы также создают защитную оболочку из веществ, входящих в плазменную клеточную мембрану.
7. На заключительном этапе новые частицы вируса высвобождаются из клетки носителя. Данный процесс происходит разными способами в зависимости от вида вируса. Некоторые клетки после этого погибают, так как клеточный лизис освобождается. В других вариантах возможно отпочковывание от клетки, однако и этот способ не предотвращает ее дальнейшее отмирание, так как повреждается мембрана плазмы.

Период до момента выхода вируса из клетки называют латентным. Продолжительность данного промежутка может находиться в пределах от нескольких часов до пары дней.
Геномные вирусы, содержащие ДНК
Вирусы, содержание ДНК геномного вида подразделяются на четыре группы:
1. Такие геномы, как адено-, папова- и герпесвирусов, переносятся и копируются в клеточном ядре носителя. Это вирусы, содержащие двухцепочечную ДНК. Капсиды, попав в клетку, переносятся к мембране клеточного ядра, чтобы потом, под воздействием определенных факторов, ДНК вируса перешло в нуклеоплазму и накопилось там. Вирусы при этом пользуются матрицей РНК и клеточными ферментами носителя. А-белки переносятся первыми, за ними следуют b-белки и g-белки. Матрица РНК возникает на основе а-22 и а-47. Полимераза РНК реализует перенос ДНК, который размножается по принципу перекатывающегося кольца. Капсид, в свою очередь, возникает из g-5 белка. Какие еще существуют геномы ДНК-содержащих вирусов?
2. Поксивирусы входят во вторую группу. На начальном этапе действия осуществляются в цитоплазме. Там происходит высвобождение нуклеотидов и начало транскрипции. Затем образуется матрица РНК. На ранних стадиях выработки создается полимераза ДНК и около 70 белков, а также двухцепочная ДНК расщепляется полимеразой. С обоих сторон генома начинается репликация в тех местах, где на начальном этапе осуществлялось расплетение и расщепление цепей ДНК.
3. В третью группу включены парвовирусы. Размножение осуществляется в клеточном ядре носителя и находится в зависимости от функций клетки. В данном случае ДНК образует так называемую шпилечную структуру и выступает в роли затравки. 125 первых пар оснований переходят с начальной цепи на смежную, которая и служит матрицей. Таким образом, происходит инверсия. Для синтезирования нужна полимераза ДНК, благодаря которой происходит транскрипция вирусного генома.
8. В четвертую группу входят гепаднавирусы. Сюда относится ДНК-содержащий вирус гепатита. ДНК вируса кольцевого типа работает в качестве основы для выработки мРНК вируса и плюс-цепи РНК. Она, в свою очередь, становится матрицей для синтеза минус-цепи ДНК.
Методы борьбы
ДНК - содержащие вирусы, безусловно, представляют опасность для здоровья человека. Основным методом борьбы с ними может стать проведение профилактических мер, направленное на укрепление иммунитета, а также регулярная вакцинация.
Как правило, антитела, направленные на борьбу с теми или иными вирусами вырабатываются как результат вторжения вредоносных микроорганизмов в систему носителя. Однако усилить выработку антител можно заранее, сделав профилактическую прививку.
Виды вакцинации
Существует несколько основных видов вакцинации, включающих:
1. Введение в организм ослабленных клеток вируса. Это провоцирует выработку увеличенного количества антител, что позволяет бороться с нормальным вирусным штаммом.
2. Введение уже мертвого вируса. Принцип действия аналогичен первому варианту.
3. Пассивная иммунизация. Данный метод заключается в введении уже синтезированных антител. Это может быть как кровь человека, перенесшего заболевание, против которого делается прививка, так и животного, например, лошадей. Последовательность размножения ДНК-содержащих вирусов мы рассмотрели.
Чтобы избежать заражения организма различными видами вирусов, опасных для здоровья человека, следует беречь тело от потенциального контакта с патогенными микроорганизмами. Вполне по силам избежать токсоплазмы, микоплазмы, герпеса, хламидий и других распространенных форм вируса, просто соблюдая определенные рекомендации. Это особенно касается детей до 15 лет.
Если тело ребенка не было заражено вышеперечисленными штаммами вирусов, то у него вырабатывается в подростковом периоде здоровый и усиленный иммунитет. Главная опасность вирусов - не всегда в том, как они выражаются, а в том, какое влияние оказывают на защитные свойства нашего организма. Примеры ДНК- и РНК-содержащих вирусов интересуют многих.
Герпесный вирус, который присутствует в теле 9 из 10 жителей Земли, снижает иммунные свойства примерно на 10 процентов в течение всей жизни, хотя может и никак не проявлять себя.
Заключение
Помимо подобной вирусной нагрузки, которая подчас не ограничивается только герпесом, условия современной жизни далеки от идеала, что также сказывается на защитных барьерах организма. В данный пункт можно отнести форсированный городской ритм жизни, плохую экологию, неправильное питание и т.д. На фоне снижения общего состояния здоровья человека его организм становится менее резистентным к различным вирусам и, соответственно, частым болезням.
Читайте также: