
Какова роль сухожильных рецепторов в контроле мышечных действий
Большинство рефлексов, имеющих важное значение для самосохранения, поддержки положения тела, быстрого восстановления равновесия, осуществляется на основе "быстродействующих механизмов" и минимальным количеством причастных нейронных цепей. Сухожильные рефлексы, нервные центры которых расположены на разных уровнях спинного мозга, представляют большой интерес в клинической практике, как тест функционального состояния организма в целом и локомоторного аппарата в частности, а также для топической диагностики при повреждениях спинного мозга.
Сухожильные рефлексы называются еще миотатичнимы, или Т-рефлексами (от лат. Tendo - сухожилия), поскольку вызываются растяжением мышц при ударе неврологическим молоточком в месте проекции соответствующего сухожилия. Однако сухожильные рецепторы при постукивании молоточком НЕ раздражаются, потому что есть высокопороговых, их активация происходит только при растяжении мышцей. Клинически важные рефлексы на растяжение мышечных веретен (миотатични) представлены в табл. 4.1 и на рис. 4.11.
К сухожильных рефлексов также относят:
■ Периостальный пястно-лучевой рефлекс, вызываемый ударом молоточка по сухожилию шиловидного отростка лучевой кости. реакция

РИС. 4.10. Контур рефлекса на растяжение (А) и рефлекторная дуга спинального миотатичного рефлекса (Б): 1 - мотонейроны мышц-разгибателей; 2 - мотонейроны мышц-сгибателей; 3 - тормозной нейрон; 4 - мышечное веретено. Знак "+" - возбуждения. Знак "-" - торможение
ТАБЛИЦА 4.1. Миотатични сухожильные рефлексы
Название рефлекса ке растяжения
Раздражение, что приводит к активации рецепторов мышц - мышечных веретен
Характер рефлекторного ответа
Место нахождения нервного центра рефлекторной дуги в спинном мозге
Локтевой изгибающий рефлекс предплечья
Постукивание неврологическим молоточком по сухожилию т. biceps brachii
V-IV шейные сегменты
Разгибательный рефлекс предплечья
Постукивание неврологическим молоточком по сухожилию т. triceps brachii
VII-VIII шейные сегменты
Легкий удар неврологическим молоточком по сухожилию ниже надколенника
III-IV поясничные сегменты
Удар неврологическим молоточком по пяточном (ахилловом) сухожилию
Подошвенное сгибание стопы
1-II крестцовые сегменты

РИС. 4.11. Рефлексы с верхних и нижних конечностей
в ответ - сгибание руки в локтевом суставе, пронация кисти и сгибание пальцев. Составные части рефлекторной дуги: нервы - срединный, лучевой, мышечно-кожный; V-VIII шейные сегменты спинного мозга, иннервирующих мышцы-пронаторы, плече-лучевой мышцу, сгибатели пальцев, двуглавая мышца плеча.
■ Н-рефлекс растяжения (Гофмана) вызывается у человека электрическим раздражением в подколенной ямке большеберцовой нерва. Эффектор - Камбалообразные мышцу. Регистрация электромиографического.
Сухожильные рецепторы Гольджи и двигательные спинальные рефлексы с рецепторов сухожилий
Сухожильные рецепторы Гольджи - это веретенообразные структуры, состоящие из сухожильных (коллагеновых) нитей, которые отходят от 9-13 экстрафузальных мышечных волокон и многочисленных ветвей безмиелиновых нервных окончаний, которые являются продолжениями толстых миелиновых волокон типа lb (Αβ), окруженных капсулой, граничащей с мышцей. Нервные волоконца содержатся среди пучков сухожильных нитей в заполненном жидкостью пространстве (рис. 4.12).
Рефлексы с рецепторов Гольджи возникают при их растяжении сокращенным скелетных мышц, с которым соединено сухожилия. Информация от сухожильных рецепторов передается афферентными нервными волокнами группы Αβ в задние рога спинного мозга и через интернейронов вызывает торможение альфа мотонейронов, иннервирующих мышцы, которые сокращаются; это приводит к их расслаблению. Мотонейроны, которые иннервируют мышцы-антагонисты, возбуждаются, возникает их сокращение (рис. 4.13).

РИС. 4.12. Строение сухожильного рецептора Гольджи
Таким образом, каждый скелетная мышца имеет две системы обратной связи с нервным центром при осуществлении рефлекторной регуляции:
■ первая система обратной связи - это сигнализация от мышечных веретен о скорости сокращения и длину мышцы;
■ вторая система обратной связи - информация от сухожильных рецепторов Гольджи о степени напряжения (силу сокращения) скелетной мышцы.

РИС. 4.13. Строение рефлекторных дуг при раздражении сухожильных рецепторов Гольджи. Знак "+" - возбуждение, знак "-" торможения
Таким образом, с рецепторами Гольджи связано как сокращение, так и расслабление мышцы. Если интрафузальных волокна определяют длину мышцы и изменения его длины, то сухожильные - напряжение мышцы, которая изменяет собственное напряжение рецептора.
Физиологическую роль рефлексов с рецепторов сухожилий определяют как защитные рефлексы, потому сильное сокращение мышц, что приводит к растяжению сухожилий и активации высокопороговых сухожильных рецепторов, может привести к повреждению как мышц, так и сухожилий. Хотя эти рефлексы относятся к защитным рефлексов, основная их роль заключается в регуляции напряжения мышц при нормальном их сокращении. Рефлексы с рецепторов сухожилий показывают, что сила сокращения мышцы является стимулом, который приводит к своего собственного расслабления.
Здравствуйте? уважаемые читатели!
Из статьи Вы узнаете, почему при физических нагрузках мышцы человека не всегда подчиняются воле человека, какую роль в этом играет сухожильный орган Гольджи, где он находится и как устроен, а также как его обмануть и стать сильнее.
Благодаря чему мы чувствуем свое тело?
Человек способен воспринимать положение своего тела в пространстве и координировать движения благодаря кинестетическим проприорецепторам. К ним относятся мышечные веретена, сухожильный орган Гольджи, рецепторы суставных связок и капсул, тактильные рецепторы кожи. Все кинестетические рецепторы по-разному участвуют в регуляции положения тела в пространстве. Например, импульсы от рецепторов суставов идут непосредственно в кору головного мозга человека, поэтому движения в суставах хорошо осознаются. Сигналы прочих кинестетических рецепторов поступают в мозжечок, где они обрабатываются без прямого участия сознания. Общая картина положения тела в пространстве, его движений, деформаций и смещения, складывается в коре головного мозга путем слияния данных об импульсах, поступивших со всей системы проприорецепции.

Мышечные веретёна и сухожильные органы Гольджи
Как наш мозг узнает о том, что происходит в мышцах?
Продолговатые мышечные веретена, также называемые рецепторами растяжения, находятся в глубине мышечной ткани. Количество их в разных мышцах неодинаково, на один грамм ткани может приходиться от нескольких штук, до сотни мышечных веретён. Длиной они обычно всего несколько миллиметров, шириной десятые доли миллиметра. Подобно датчикам, эти веретёна воспринимают скорость и силу растяжения мышечных волокон, колебания их длины, активность двигательных нейронов.

Сухожильный орган Гольджи
Строение сухожильного органа Гольджи
Во всех мышцах человеческого тела есть рецепторы, воспринимающие степень их напряжения, которые называются органами Гольджи, в честь учёного, впервые их открывшего. Рассмотрим, каково их строение.
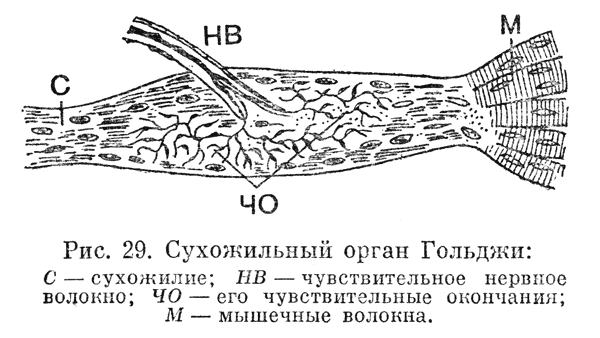
Сухожильный орган Гольджи
Каждый такой орган представляет собой сплетение лежащих параллельно тонких нервных волокон и коллагеновых сухожильных волокон, заключенное в капсулу цилиндрической формы, образованную соединительной тканью. Длина органа Гольджи составляет всего один миллиметр. Через этот сухожильный рецептор проходит порядка десяти-пятнадцати волокон мышечного сухожилия, которое представляет собой соединительную ткань, проходящую через мышцу и прикрепляющую её к кости. Нервные волокна, пролегающие в органе Гольджи, лишены своей миелиновой оболочки, что улучшает их контакт с мышечным волокном.
Сухожильный орган Гольджи
Когда мышца приходит в напряжение – сокращается или растягивается – сухожильные волокна напрягаются и оказывают давление на расположенные между ними нервные окончания. Орган Гольджи передает сигнал о степени и скорости нарастания напряжения в центральную нервную систему. Импульсы от сухожильных рецепторов идут по крупным нервным волокнам. Их диаметр составляет шестнадцать мкм (микрон), что лишь немногим меньше толщины самих мышечных волокон. Подобная толщина обеспечивает высокую скорость передачи информации, которая поступает сначала в спинной мозг, затем по его каналам в мозжечок, а через него - в кору головного мозга. Рецепторы Гольджи непрерывно передают информацию о состоянии любого участка мускулатуры организма, даже когда человек полностью расслаблен (фоновые сигналы), что способствует формированию мышечно-суставного чувства (мы чувствуем своё тело).
Зачем нам нужен тормозящий Гольджи-рефлекс?
Если поступившая в спинной мозг информация о напряжении мышцы распознается как угрожающая её целостности, то возбуждается тормозной вставочный нейрон, который тормозит соответствующий передний двигательный нейрон, контролирующий непосредственно мышцу. Так возникает локальный тормозной рефлекс, называемый аутогенным торможением или Гольджи-рефлексом. Он приводит к мгновенному расслаблению перенапрягшейся мышцы, но никак не сказывается на прилегающих к ней.
Этот процесс защищает организм от травм, ведь чрезмерное напряжение может стать причиной разрыва мышечной ткани или отрыва слабо тянущихся сухожилий от кости. В лабораторных условиях подобное травмирование происходило в отсоединенных от спинного мозга конечностях, избыточное напряжение которых достигалось путем проведения разряда электрического тока. А при экспериментальном выключении рецепторов Гольджи препаратом, лабораторные животные теряли способность двигаться и поддерживать устойчивые позы.
Каждый из нас много раз испытывал тормозящий Гольджи-рефлекс. Очень ярко он выражается, когда мы хотим опустить на пол что-то тяжелое. Допустим, вы держите двумя руками у груди большую гирю, которую хотите опустить на пол. При этом у вас напряжены бицепсы. Чтобы аккуратно опустить гирю, вам нужно разогнуть руки, то есть растянуть мышечные волокна. Но тормозящий Гольджи-рефлекс не позволяет сильно напряженным мышцам растянуться еще больше, вместо этого они наоборот расслабляются. Ваша гиря с грохотом падает на пол, соседям снизу становится невесело. Зато у вас сухожилия от костей не оторвались! Защитная функция организма сработала!
Помимо защиты мышц от возможных травм, сигналы, поступающие с рецепторов Гольджи, помогают организму равномерно напрягать мускулатуру, которая должна действовать синхронно. Благодаря постоянному контролю состояния мышц, происходит перераспределение напряжения в них – чрезмерно возбужденные волокна расслабляются, а чересчур расслабленные – сокращаются, и мышцы более эффективно выполняют свои функции. Представьте теперь, как будто вы одной рукой поднимаете и опускаете легкую для вас гирю. Вы сгибаете и разгибаете локоть, при этом разные мышцы вашей руки синхронно то напрягаются, то расслабляются. Координируется этот процесс при участии нервных импульсов, поступающих с сухожильных органов Гольджи.
Как обмануть сухожильный рефлекс и стать сильнее?
В организме человека заложено много возможностей для активации его резервов. Путем систематических тренировок, можно улучшить работу любого органа, даже маленького сухожильного органа Гольджи, который можно разглядеть лишь в микроскоп.
Считается, что лучший способ тренировки органа Гольджи – это статические нагрузки.
То есть, вы берёте штангу и просто её удерживаете, пока ваши пальцы не разожмутся от перенапряжения. Разумеется, для безопасности этого упражнения вам понадобится ограничивающая падение штанги силовая рама или страхующий вас напарник (если штанга над Вами, например, в жиме лёжа). Чем больший вес на штанге, которую вы удерживает, тем сильнее напрягаются Ваши мышечные волокна, и органы Гольджи соответственно. Если вы регулярно будете совершать упражнение с данным весом, Ваша мускулатура начнет привыкать к подобной нагрузке, напряжение перестанет восприниматься как излишнее, и блокирующий сигнал будет подан гораздо позже. Однако мы рекомендуем вам делать это упражнение не чаще раза в неделю, так как оно вызывает достаточно сильное напряжение центральной нервной системы, что при злоупотреблении может привести к перенапряжению.
Если через неделю Вы возьмете штангу с большим весом, а потом снова первую, то она покажется Вам гораздо легче, чем была в прошлый раз, и Вы сможете удерживать ее гораздо дольше. Причина этому – сравнение центральной нервной системой силы импульсов, сообщающих о перенапряжении мышц, которые поступают с проприорецепторов. Вы сможете дольше удерживать вес за счет тренировки органа Гольджи.
Этот способ использован в моём силовом цикле по увеличению жима лёжа, в котором раз в неделю используются субмаксимальные нагрузки.
Феноменальная сила. Обратная сторона
Возможно и самопроизвольное отключение тормозящего сухожильного рефлекса в стрессовой ситуации. Вы могли слышать о подобных историях, когда, например, не занимающийся спортом человек вдруг приподнимает автомобиль, чтобы спасти зажатого ребенка, или перескакивает с разбега через двухметровый забор, спасаясь от собак.
Не рекомендую Вам пытаться вызвать подобный эффект целенаправленно. Регулярные тренировки с субмаксимальными или статическими нагрузками в зале не столь героичны, зато намного более безопасны для Вашего организма. Ведь в случае опасности организм рискует здоровьем ради спасения своей жизни или кого-то из близких, и только в таких случаях это оправдано.
Практически в каждой мышце находятся рецепторы растяжения, называемые из-за своей формы мышечными веретенами. Эти образования называют интрафузальными мышечными волокнами, все прочие, составляющие основную массу мышцы и обеспечивающие ее работу – экстрафузальными. Различия в их размерах весьма значительны: диаметр первых 15-30 мкм, длина-4-7 мм, тогда как у вторых диаметр примерно 50-100 мкм и длина от нескольких миллиметров до многих сантиметров. Каждым своим концом мышечное веретено прикрепляется к соединительнотканной оболочке пучка экстрафузальных волокон при помощи напоминающих сухожилия полосок соединительной ткани длиной 0,5–1 мм.
Основная сенсорная иннервация мышечных рецепторов растяжения обеспечивается афферентными волокнами, ветви которых образуют аннулоспиральные (первичные) окончания. Эти толстые миелинизированные афференты называются волокнами 1а (классификация Ллойда-Ханта для сенсорных нервов). Многим (хотя и не всем) веретенам свойственна также вторичная сенсорная иннервация. Ее афферентные волокна (волокна группы II) тоньше. По аналогии с первичными окончаниями, названными так из-за принадлежности к волокнам 1а, волокнам группы II соответствуют вторичные окончания мышечных веретен. По форме они сходны с первичными, но гораздо менее однородны – часто напоминают спираль, но иногда ветвятся наподобие соцветия.
Наряду с сенсорной, у интрафузальных мышечных волокон (как и у экстрафузальных) есть двигательная иннервация. Эти двигательные аксоны тоньше обычных (волокон Аα, или сокращенно α-волокон) и называются волокнами Аγ (сокращенно γ-волокнами). Плотность мышечных веретен, т.е. их число на 1 г мышечной ткани, особенно высока в мелких мышцах, участвующих в выполнении тонких движений, например в мелких мышцах кисти.
В сухожилиях всех мышц вблизи от сухожильно-мышечного соединения находятся особые рецепторы – сухожильные органы Гольджи. Они состоят из сухожильных нитей, заключенных в соединительнотканную капсулу. К ней подходят одно или два толстых миелинизированных афферентных аксона, которые называются волокнами 1b.
По типу адекватного раздражителя мышечные веретена и сухожильные органы относятся к рецепторам растяжения. Однако их расположение различно; первые лежат параллельно экстрафузальным волокнам, а вторые соединены с ними последовательно. В результате неодинаков и характер их активации, особенно во время сокращения мышцы.
Следовательно, мышечные веретена регистрируют главным образом длину мышцы, а сухожильные органы – ее напряжение.
1а-волокна мышечных веретен образуют возбуждающие синапсы на α-мотонейронах их собственных мышц. Активация первичных окончаний мышечных веретен при растяжении мышцы ведет, таким образом, к возбуждению мотонейронов. (миотатический рефлекс). Сила рефлекса существенно колеблется в зависимости от активности других (облегчающих или тормозных) входов к участвующим в нем нейронам. Поэтому, за исключением крайних случаев, клиническим показателем служит не столько ее абсолютная величина, сколько разница между рефлексами с правой и с левой стороны тела или аномалии некоторых из них, выявляемые при сравнении с общим рефлекторным поведением.
Физиологическое значение миотатического рефлекса в том, что он регулирует длину мышц, что особенно важно для сохранения тонуса позных мышц. Например, когда человек стоит, каждое сгибание в коленном суставе, даже такое слабое, что его невозможно ни увидеть, ни почувствовать, сопровождается растяжением четырехглавой мышцы и соответствующим усилением активности первичных окончаний мышечных веретен. В результате происходит дополнительная активация α-мотонейронов четырехглавой мышцы и повышение ее тонуса, противодействующее сгибанию. И наоборот, слишком сильное сокращение мышцы ослабляет стимуляцию ее рецепторов растяжения. Частота их импульсации, возбуждающей мотонейроны, уменьшается, и мышечный тонус ослабевает. Такова регулирующая цепь, поддерживающая постоянство мышечной длины.
Волокна 1а образуют не только возбуждающие связи с мотонейронами собственной мышцы, но и тормозные связи с мотонейронами-антагонистами. Тормозная рефлекторная дуга включает центральный интернейрон, следовательно относится к дисинаптическому типу – в ней два центральных синапса: возбуждающий между волокнами 1а и интернейронами и тормозной между аксонами интернейронов и мотонейронами. Это реципроктное антагонистическое торможение – за счет такой рефлекторной дуги мотонейроны каждой мышцы-антагониста в паре (например, сгибателя и разгибателя в данном суставе) тормозятся во время активации другого компонента пары.
Мышечные веретена могут вызывать сокращения экстрафузальных волокон в двух случаях не только при растяжении мышцы, но и в результате сокращения интрафузальных волокон, вызванного γ-мотонейронами. В этом случае активность волокон 1а непосредственно возбуждает α–мотонейроны. Данный механизм получил название γ-петли.
Обычно α-мотонейроны активируются супраспинальными центрами. При этом движения быстрые, но их недостаток состоит в нарушении тонкого равновесия системы регуляции мышечной длины, включающей рефлекс растяжения. В результате растяжение веретен соответствующей мышцы может стать недостаточным (подпороговым) или избыточным (насыщающим). В отличие от этого, активация γ-петли обеспечивает укорочение мышцы с минимальными изменениями частоты импульсации афферентов мышечных веретен.
Вслед за сокращением интрафузальных мышечных волокон следует усиление сокращения экстрафузальных мышечных волокон до тех пор, пока не восстановится исходная частота импульсации первичных аффе-рентов веретен. Значит, γ-петля, включающая дугу рефлекса растяжения, представляет собой сервомеханизм, в котором длина мышечных веретен (регулирующая переменная) управляет длиной мышцы (регулируемая переменная). α- и γ-мотонейроны возбуждаются одновременно. Такая коактивация α - и γ-мотонейронов называется также α-γ-сопряжением. При этом чувствительность рецептора регулирующей системы (первичных окончаний мышечных веретен) поддерживается в адекватном диапазоне, а движение развивается плавно, автоматически учитывая нагрузку на мышцу (поднос с посудой и т.д.).
Действие афферентов группы IIа мышечных веретен в отличие от волокон 1а не ограничивается только синергистами и антагонистами одного и того же сустава, а регулирует движение всей конечности.
Сухожильные органы образуют ди- и трисинаптические тормозные связи с мотонейронами своей мышцы. Так как сухожильные органы – рецепторы напряжения мышцы, его увеличение при сокращении экстрафузальных волокон приводит за счет активации афферентов 1b к торможению мотонейронов. И напротив, ослабление мышечного тонуса вызывает их растормаживание (уменьшение степени торможения) и тем самым активацию. Другими словами, рефлекторная дуга сухожильных органов служит для поддержания постоянства напряжения мышцы.
Следовательно, у каждой мышцы две регуляторные системы обратной связи: регуляции длины с мышечными веретенами в качестве рецепторов и регуляции напряжения, рецепторами в которой служат сухожильные органы.
С точки зрения теории систем управления не вполне понятно, зачем нужно регулировать одновременно и длину, и напряжение. При идеальной организации системы контроля длины развиваемая мышцей сила всегда пропорциональна частоте эфферентной импульсации α-мотонейронов, и контроль напряжения оказывается излишним. Но сила, развиваемая мышцей, зависит и от других факторов – скорости сокращения, утомления. Обусловленное этими факторами отклонение мышечного напряжения от нужной величины регистрируется сухожильными органами и корректируется системой регуляции напряжения.
| | | следующая лекция ==> | |
| III. Проводниковые функции спинного мозга. | | | VI. Функциональные возможности изолированного спинного мозга. Спинальный шок. |
Дата добавления: 2017-09-19 ; просмотров: 606 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Представляют собой небольшие продолговатые образования, напоминающие своим внешним видом прядильные капсулы мышечного веретена находится пучок мышечных волокон, которые называются интрафузальными, т. к. они расположенными внутри веретена в отличие от обычных мышечных волокон, которые называются зкстрафузальными.
Каждое интрафузальное волокно состоит из трех частей:
• его центральная часть называется ядерной сумкой, в которой находятся ядра мышечной клетки;
• два периферических участка, которые имеют поперечную исчерченность и обладают способностью сокращаться;
• миотрубки, расположенные между ядерной сумкой и периферическими участками.
Ядерную сумку в виде спирали окружают нервные волокна чувствительного нейрона-первичные рецепторные окончания. В области миотрубок нервные окончания афферентных нейронов гроздевидно ветвятся, образуя вторичные рецепторные окончания.
В мышце мышечное веретено одним концом прикрепляется к экстрафузальному мышечному волокну, а другим - к сухожилию этого волокна. Таким образом, мышечное веретено расположено в мышце параллельно экстрафузальным мышечным волокнам
· Сухожильные органы Гольджи.
Располагаются в местах соединения мышечных волокон с коллагеновыми пучками сухожилий. Весь орган окружён тонкой соединительнотканной капсулой. Нервное волокно, подходящее к веретену утрачивает миелиновый слой и, распадаясь на терминальные ветви, проходит между пучками коллагеновых волокон сухожилия.
Реагируют на изменение уровня напряжения мышцы и натяжения коллагеновых волокон сухожилий. Участвуют в реализации рефлекторной регуляции напряжения мышц.
Расположенные в фасциях, сухожилиях, связках. Информируют ЦНС о степени сгибания или разгибания в этом суставе.
Состоит из отростков изменённых клеток реснитчатого эпителия с вторичночувствующими ресничками. Тела клеток отделены от контактной зоны капсулой. Реснички вторичночувствующих клеток располагаются между наружной и внутренней капсулами, контактируя с внутренней поверхностью наружной капсулы. Наружная капсула имеет строение, аналогичное внутренней.
2) Децеребрационная ригидность, её механизмы и проявления.
Децеребрационная ригидность - повышенние тонуса мышц экстензоров (разгибателей) и относительное расслабление мышц-сгибателей, возникает в результате перерезания стволовой части головного мозга (на уровне среднего мозга) — децеребрации , объясняется прекращением тормозящего действия красного ядра на ядро Дейтерса.
Похожие симптомы развиваются при сдавлении ствола мозга односторонним, ограничивающим внутричерепное пространство процессом в полушариях головного мозга, особенно в его задних отделах. Это прежде всего опухоли и абсцессы мозга и мозжечка, внутримозговые гематомы. Также может быть вызвана отеком и набуханием мозга при обширном инфаркте мозга, ушибе мозга, менингите и менингоэнцефалите, при токсических энцефалопатиях, почечной коме и др.
Наиболее характерная поза больного: голова запрокинута назад, спина выгнута дугой, руки разогнуты, предплечья ротированы внутрь, кисти и пальцы согнуты, ноги выпрямлены и повернуты внутрь, стопы разогнуты. Сознание утрачено или затемнено. Может сопровождаться остановкой дыхания.
3) Сенсорные системы человека, классификация, общие свойства и различия. Явление адаптации в сенсорных системах.
Сенсорные системы - часть его нервной системы, способная воспринимать внешнюю для мозга информацию, передавать ее в мозг и анализировать.
1. Соматовисцеральная (тактильная, проприорецептивная, температурная, болевая, висцеральная).
- Формирование ощущений и восприятие действующих стимулов;
- Контроль произвольных движений;
- Контроль деятельности внутренних органов;
- Необходимый для бодрствования человека уровень активности мозга.
Все сенсорные системы состоят из периферических рецепторов, проводящих путей и переключательных ядер, первичных проекционных областей коры и вторичной сенсорной коры. Организованы иерархически. Низший уровень переработки обеспечивают первичные сенсорные нейроны, которые расположены в специализированных органах чувств или в чувствительных ганглиях и предназначены для проведения возбуждения от периферических рецепторов в центральную нервную систему. Периферические рецепторы — это чувствительные высокоспециализированные образования, способные воспринять, трансформировать и передать энергию внешнего стимула первичным сенсорным нейронам. Центральные отростки первичных сенсорных нейронов оканчиваются в головном или спинном мозге на нейронах второго порядка, тела которых расположены в переключательном ядре. В нем имеются не только возбуждающие, но и тормозные нейроны, участвующие в переработке передаваемой информации. Представляя более высокий иерархический уровень, нейроны переключательного ядра могут регулировать передачу информации путем усиления одних и торможения или подавления других сигналов. Аксоны нейронов второго порядка образуют проводящие пути к следующему переключательному ядру, общее число которых обусловлено специфическими особенностями разных сенсорных систем. Окончательная переработка информации о действующем стимуле происходит в сенсорных областях коры.
1. Действие раздражителя на периферические рецепторы;
2. Преобразование энергии стимула в электрические сигналы — потенциалы действия, возникающие в первичном сенсорном нейроне;
3. Последующую переработку передаваемых сигналов на всех иерархических уровнях сенсорной системы;
4. Возникновение субъективной реакции на раздражитель, представляющей собой восприятие или внутреннее представительство действующего стимула в виде образов или словесных символов.
При длительном действии стимула порог чувствительности рецепторов к нему может повышаться, что определяется как адаптация рецепторов. Механизмы адаптации не одинаковы в рецепторах разных модальностей, среди них различают быстро адаптирующиеся (например, тактильные рецепторы кожи) и медленно адаптирующиеся рецепторы (например, проприоцепторы мышц и сухожилий). Быстро адаптирующиеся рецепторы сильнее возбуждаются в ответ на быстрое нарастание интенсивности стимула (фазический ответ), а их быстрая адаптация способствует освобождению восприятия от биологически незначительной информации (например, контакт между кожей и одеждой). Возбуждение медленно адаптирующихся рецепторов мало зависит от скорости изменения стимула и сохраняется при его длительном действии (тонический ответ), поэтому, например, медленная адаптация проприоцепторов позволяет человеку получать нужную ему для сохранения позы информацию в течение всего необходимого времени.
Какие же рецепторы есть в двигательном аппарате и как они возбуждаются при собственных рефлексах?
Двигательный аппарат отличается наличием очень специализированных и сложно устроенных рецепторных структур, которые могут быть разделены на два основных типа. Рецепторы одного из них (мышечные веретена) расположены в мышцах, а рецепторы другого - в сухожилиях и суставах. Мышечные веретена расположены между мышечными волокнами и одеты капсулой, более широкой в средней части и утончающейся по концам. Внутри каждого веретена также есть интрафузальные мышечные волокна, отличающиеся по строению от обычных (они имеют другое содержание миофибрилл и саркоплазмы и приближаются по строению к эмбриональным волокнам). Важной особенностью мышечных волокон, расположенных внутри веретена, является наличие у них наряду с двигательными окончаниями рецепторных структур. Последние могут быть двух типов - в виде спирали, обвивающей интрафузальное волокно (аннуло-спиральные рецепторы), и в виде грозди бляшек. Аннуло-спиральные рецепторы соединены с более толстыми афферентными волокнами, а гроздьевидные - с более тонкими.
Двигательные нервные волокна, иннервирующие интрафузальные мышечные волокнам имеют свои особенности. Они значительно тоньше, чем обычные двигательные волокна. Если обычные двигательные волокна относятся в группе А - альфа, по классификации Й..Эрлангера и X.Гассера (1927), то двигательные волокна, иннервирующие мышечные волокна, относятся к группе А -гамма. Они являются аксонами особых мотонейронов, соответственно также обозначаемых как гамма - мотонейроны.
Рецепторные образования, расположенные в сухожилиях, имеют совершенно другой тип строения. Наиболее характерными здесь являются образования Гольджи. Они состоят из сложной соединительнотканной капсулы, внутрь которой входит афферентное волокно, свивающееся затем в довольно густой клубок. Капсула расположена среди соединительнотканных волокон сухожилия. Инкапсулированные окончания есть также в суставных поверхностях и в фасциях двигательного аппарата.
Кроме этих основных типов, в двигательном аппарате есть и другие, более простые рецепторы, в том числе свободные нервные окончания, Они, по-видимому, не связаны с собственными рефлексами мышц, а представляют собой болевые окончания.
Подробные исследования особенностей афферентного разряда от мышечных рецепторов были проведены в 1933 г. английским физиологом Б. Мэттьюсом, который впервые применил для этой цели отведение потенциалов действия от отдельных афферентных волокон в составе мышечного нерва. Оказалось, что афферентные волокна, идущие от мышцы, по характеру разряда делятся на две основные группы. Если вызвать в мышце одиночное мышечное сокращение, то в одной из этих групп разряд развивается, в основном, в момент укорочения или расслабления мышцы и замолкает в период максимального натяжения, у другой группы, наоборот, разряд усиливается тогда, когда мышца развивает максимальное натяжение.
Несомненно, что эти два типа разрядов соответствуют описанному выше подразделению рецепторов на мышечные веретена и сухожильные рецепторы. Можно себе представить несколько схематически, что механическое действие мышечного сокращения на собственно мышечные и сухожильные рецепторы в какой-то степени противоположно. Сухожильные рецепторы включены как бы последовательно с сократительными элементами, которые механически растягиваются при сокращении. Следовательно, максимальное раздражение таких рецепторов будет наблюдаться как раз на высоте сокращения. Мышечные же веретена расположены по существу параллельно обычным мышечным волокнам. Поэтому при сокращении мышцы интрафузальное волокно окажется, наоборот, расслабленным, и раздражение его рецепторов уменьшится.
Конечно, если мышца растягивается внешней силой, а не сокращается самостоятельно, то особенности раздражения рецепторов изменятся. В этом случае растягиваются как сухожильные, так и интрафузальные структуры, и все типы окончаний будут давать разряд импульсов (хотя порог их раздражения может быть различным).
Специальные исследования зависимости между различными видами собственных рефлексов реакций мышц и особенностями импульсации, поступающей от различных групп рецепторов двигательного аппарата, показали, что основой возникновения как сухожильных рефлексов, так и рефлексов растяжения являются сигналы от рецепторов мышечных веретен. На первый взгляд может показаться странным, почему в таком случае сухожильные рефлексы вызываются воздействием не на мышцы, а на сухожилия, как это подчеркивается и в самом их названии. Однако можно довольно просто убедиться, что к возникновению этих рефлексов рецепторы сухожилия отношения не имеют и что основой рефлекса является вызываемое ударом кратковременное растяжение мышцы. Так, при анестезировании сухожилия удар по нему вызывает такой же сухожильный рефлекс, как и в обычных условиях.
Может показаться странным и то обстоятельство, что возбуждение одних и тех же мышечных рецепторов вызывает две очень различные по своему проявлению рефлекторные реакции. При сухожильном рефлексе это кратковременное фазное движение, а при рефлексе растяжения - длительное тоническое напряжение. Причина различия в этом случае заключается, очевидно, только в том, что в связи с различным видом механического действия афферентная импульсация, поступающая от рецепторов мышечных веретен, оказывается различным образом организованной во времени. Когда наносится удар по сухожилию, то это вызывает кратковременное сильное растяжение и почти синхронный, кратковременный разряд импульсов в соответствующих афферентных волокнах. Если же мышечное веретено растягивается длительно, то в афферентных волокнах возникают не кратковременные синхронные разряды, а постоянная импульсация низкой частоты, длящаяся все время, пока продолжается растяжение мышцы. Постоянная афферентная импульсация и поддерживает тоническое возбуждение соответствующих мотонейронов. Не исключено, конечно, что различные типы окончаний мышечных веретен могут быть в различной степени ответственны за фазный и тонический характер возникающей рефлекторной реакции. Аннуло-спиральные рецепторы сигнализируют в зависимости от скорости растяжения мышцы, они и быстрее адаптируются, гроздьевидные окончания более чувствительны к изменению длины мышцы и медленнее адаптируются, поэтому они дают более стойкий разряд афферентных импульсов, способствуют поддержанию тонического характера рефлекторной реакции.
Если таким образом и сухожильные рефлексы и рефлексы растяжения связаны с возбуждением рецепторов мышечных веретен, то какова же функция рецепторов, расположенных в сухожилиях и, в частности, рецепторов Гольджи? Есть основания считать, что возбуждение рецепторов Гольджи является причиной аутогенного торможения собственных рефлексов мышц. Возбуждение афферентных волокон, которые идут от рецепторов Гольджи, вызывает развитие тормозящих постсинаптических потенциалов в соответствующих мотонейронах и прекращает рефлекторный тонус мышцы при ее чрезмерном растяжении. Известно, что порог возбуждения рецепторов Гольджи при механическом растяжении мышцы выше, чем аналогичный порог возбуждения мышечных веретен. Это различие хорошо согласуется с теми условиями раздражения, при которых развивается аутогенное торможение.
Переходя к рассмотрению центральных механизмов собственных рефлексов мышц, необходимо в первую очередь отметить уже упоминавшуюся характерную особенность сухожильных рефлексов - исключительно короткий скрытый период. Конечно, у рефлексов растяжения измерить скрытый период нельзя, так как это тонический рефлекс, который, постепенно нарастая, длится все время раздражения. Однако можно думать, что поскольку оба рефлекса вызываются возбуждением одних и тех же рецепторов, следовательно, одних н тех же афферентных волокон, то они связаны с деятельностью одних и тех же центральных механизмов.
Одним из условий, которые обеспечивают исключительную быстроту передачи соответствующих нервных влияний, является строение афферентных волокон, которые идут от мышечных рецепторов. Они относятся к самым толстым нервным волокнам (группа А -альфа), имеющим скорость проведения до 120 м/с. Для проприоцептивных афферентных волокон млекопитающих Д. Ллойд и А. Мак-Пита в 1950 г. предложили специальную классификацию, в которой различные группы обозначаются не буквами, а римскими цифрами (группы I, II, III и IV). Группа I соответствует примерно волокнам А - альфа по классификации Й. Эрлангера и X. Гассера и объединяет быстропроводящие волокна, обслуживающие мышечные и сухожильные рецепторы. Афферентные волокна, идущие от сухожильных рецепторов, несколько тоньше по диаметру, чем афферентные волокна, идущие от мышечных рецепторов. Поэтому группа I подразделяется на подгруппу Iа и подгруппу Ib; афферентные волокна от рецепторов мышечных веретен относятся к группе Iа, а афферентные волокна от рецепторов сухожилий - к группе Ib. От части рецепторов мышечных веретен идут и более тонкие волокна, относящиеся к группе II.
Большая скорость осуществления собственных рефлексов мышц обеспечивается, кроме быстроты проведения афферентной волны, также и тем, что рефлекторная дуга таких рефлексов не включает в себя вставочных нейронов.
Трудно сказать, почему в процессе эволюции именно эта рефлекторная дуга оказалась такой быстродействующей. Вероятно, быстрота осуществления собственных рефлексов очень важна для обеспечения нормального функционирования мышечного аппарата и предохранения его от перерастяжения и разрывов при различных двигательных актах.
Сказанное выше касается только возбуждающих эффектов при собственных рефлексах. Центральный скрытый период появления аутогенного торможения всегда оказывается несколько большим, чем центральный скрытый период возбуждающих эффектов (на несколько десятых долей миллисекунд). Это согласуется с общим положением о том, что тормозящие эффекты в центральной нервной системе всегда осуществляются с помощью хотя бы одного дополнительного вставочного нейрона. По-видимому, афферентные волокна, берущие начало от сухожильных рецепторов Гольджи (волокна группы Ib), не контактируют прямо с мотонейронами. Этот путь имеет вставочный нейрон, который превращает возбуждающий эффект в тормозящий. Единственным типом окончаний, которые прямо контактируют с мотонейронами, являются окончания волокон группы Iа от мышечных веретен.
Читайте также: