Биотоки улитки. Потенциал покоя и возбуждения улитки
Добавил пользователь Cypher Обновлено: 27.01.2026
Вопросы структуры и функции слуховой системы подробно изложены во многих монографиях и специальных руководствах. В связи с этим указанные вопросы будут рассматриваться лишь с позиций формирования сенильных изменений слуха.
Наружное ухо. Ушная раковина человека, за исключением мочки, состоит из эластического упругого хряща, покрытого с обеих сторон кожей, которая на передней поверхности раковины интимно спаяна с надхрящницей и покрыта нежным волосяным покровом (пушком). Последний более выражен в области козелка, противокозелка и межкозелковой вырезки. Ушная раковина обеспечивает оптимальный прием звуковых волн лишь при определенном к ним ориентировании. Кроме того, ей принадлежит роль и первичного усилителя звука.
Наружный слуховой проход, состоящий из перепончато-хрящевого и костного отделов, выстлан эпидермисом, содержащим в своих глубоких отделах серные железы. Кожа слухового прохода обладает способностью к самоочищению благодаря росту эпидермиса из глубоких отделов кнаружи. Считается, что горизонтальный (тангенциальный к поверхности) рост эпидермиса из генерационного центра в области барабанного кольца составляет у человека 0,05—0,07 мм в день. Такой рост эпидермиса поддерживает чистоту слухового прохода (W. Litton, 1968). К сожалению, закономерности миграции эпидермиса в этой области досконально не изучены, в особенности у лиц старших возрастных групп. Изменения же в характере роста, преимущественно вертикальной пролиферации, могут способствовать обструкции слухового прохода (кератозы, серные пробки). Слуховой проход имеет собственный резонанс в зоне 300÷4000 Гц. Благодаря этому звук в данной области усиливается в пределах 10 дБ (G. V. Bekesy, W. A, Rosenblith, 1951).
С возрастом эластические свойства тканей изменяются. Это не может не приводить к структурным перестройкам наружного уха. Последние, в свою очередь, могут вызывать изменения коэффициента звукопередачи.
Механические структуры среднего уха в основном предназначены для передачи звуковой энергии из воздушной среды в жидкую. Благодаря высокому по сравнению с воздухом импедансу жидкости, эффективность передачи акустической энергии достигается за счет механизмов усиления звукового давления (разница в площадях барабанной перепонки и подножной пластинки стремени, а также системы рычагов слуховых косточек). Рычажный механизм косточек контролируется работой двух внутрибарабанных мышц.
За счет сокращения мышц, что особенно проявляется при действии сильных звуков, меняется амплитуда и характер движения косточек относительно друг друга. Таким образом, обеспечивается изменение коэффициента передачи энергии звука (неодинаковое для разных частот) к внутреннему уху (аккомодационная и защитная функции).
Поскольку основной путь передачи звуковой энергии к рецептору составляет система слуховых косточек, то изменения в этой системе могут иметь определенное значение в нарушении звукопроведения (понижение эластичности связок, тугоподвижиость или анкилоз суставов, дегенеративные изменения в слуховых мышцах), что априорно не может быть исключено при пресбиакузисе.
Рецепторный отдел. Спиральный (кортиев) орган, или звуковой рецептор, представляет собой совокупность нейроэпителиальных клеток, расположенных на базилярной мембране улитки, которые осуществляют преобразование звукового раздражения в физиологический акт звуковой рецепции. Физиологическая активность спирального органа неотделима от колебательных процессов в прилежащих мембранах и окружающих жидкостях, а также от уровня метаболизма всего комплекса тканей улитки, в особенности сосудистой полоски. Но лишь разработка метода прижизненной изоляции перепончатой улитки позволила изучить цитохимические структуры рецептора в условиях адекватного раздражения (Я. А. Винников, Л. К, Титова, 1961). Электронно-микроскопически строение спирального органа изучено Г. Энгстромом (Н. Engstrom, 1960) и др.
Улитка представляет собой передний отдел внутреннего уха. Завитки улитки обвивают костный стержень, несущий в себе сосуды и нервы. На поперечном срезе в каждом завитке различают два перилимфатических канала — лестницу преддверия, находящуюся выше рейсснеровой мембраны, и лестницу барабана, расположенную ниже основной мембраны. Обе лестницы соединены у верхушки улитки отверстием. Энцолимфатическое пространство в пределах улитки ограничено снизу основной мембраной, сверху — рейссиеровой мембраной и латерально-сосудистой полоской, спиральным выступом и наружной спиральной бороздкой
Спиральный орган занимает большую часть эндолимфатической поверхности базилярной мембраны, тянущейся между костной спиральной пластинкой медиально и соединительнотканной спиральной связкой латерально. Базилярная мембрана расширяется по направлению к верхушке улитки. Она состоит из 4 слоев волокон, которые электронно микроскопически не имеют даже отдаленного сходства со струнами. Над спиральным органом нависает покровная перепонка, медиально связанная с соединительнотканным утолщением. Будучи тяжелее воды, она сохраняет свое положение благодаря наличию среди ее волокнистых структур прочных коллагеновых волокон (А. И. Неворотин, М. С. Плужников, 1973). Покровная перепонка имеет свободный контакт со стереоцилиями волосковых клеток.
На поперечном срезе клеточный массив спирального opганa разделен на две части — наружную и внутреннюю треугольным пространством кортиева туннеля, заполненного «третьей» лимфой или «кортилимфой», по своему химическому составу приближающейся к перилимфе. Через туннель проходят безмякотные волокна наружного спирального сплетения и, частично, эфферентного оливокохлеарного пучка.
Спиральный орган состоит из двух видов нейроэпителиальных клеток — чувствительных (волосковых) и опорных. По пространственному отношению к кортиеву туннелю волосковые клетки делятся на внутренние и наружные. У человека примерно 3500-4000 внутренних и 20 000 наружных волосковых клеток.
Наружные волосковые клетки имеют цилиндрическую форму. Их апикальная поверхность омывается эндолимфой, а боковые поверхности — кортилимфой пространства Нуэля. Волоски (стереоцилии) этих клеток погружены основаниями в кутикулу — электронно-плотное вещество апикальных частей волосковых клеток. Стереоцилии помимо сердцевины содержат многочисленные микрофиламенты — тончайшие плотные нити. Основная масса органоидов (митохондрии, канальцы гладкого эндоплазматического ретикулюма, микротрубочки, мультивезикулярные тельца, рибосомы и др.) распределена в цитоплазме наружных волосковых клеток в отличие от внутренних неравномерно (преимущественно в верхней их части, вдоль плазматической мембраны и в базальной части).
Характерным для волосковых клеток является наличие так называемых «ламеллированных» каналов, идущих параллельно боковой плазмолемме и связывающих апикальную и базальную части клетки. Последняя богата органоидами, включая так называемые «покрытые» пузырьки, и контактирует с афферентными и эфферентными нервными окончаниями. Тела афферентных окончаний небольшие, светлые, содержат единичные пузырьки диаметром 40—50 нм и митохондрии. Пре- и постсинаптические мембраны имеют примембранные утолщения, а у некоторых видов — и синаптическую полоску. Эфферентные нервные окончания крупные, темные от множества пузырьков диаметром 30—35 нм. Эфферентный синапс имеет субсинаптическую цистерну. Электронно-микроскопически прослежено вскрытие пузырьков эфферентного окончания с выбросом содержимого в синаптическую щель, а электронно-цитохимически — наличие ацетилхолинэстеразы в области эфферентных синапсов в базальной части волосковых клеток и в стереоцилиях.
Внутренние волосковые клетки имеют кувшинообразную форму и со всех сторон окружены клеточными элементами, за исключением апикальной поверхности, омываемой эндолимфой. Они имеют принципиально сходное строение с наружными волосковыми клетками, однако количественно гораздо беднее их органоидами. Внутренние волосковые клетки связаны с опорными клетками простой межклеточной связью, тогда как наружные волосковые клетки фиксированы плотной межклеточной связью в апикальной области и специализированным усиленным контактом в базальной области.
Опорные элементы спирального органа (клетки-столбы, клетки Дейтерса и Гензена) выполняют поддерживающую (в отношении волосковых клеток) функцию за счет развитой системы плотных межклеточных связей. Опорные клетки также несут трофическую функцию, обеспечивая транспорт веществ за счет аппарата микроворсинок.
Спиральный орган не имеет сосудов. Ближайший сосуд — vas spirale — располагается па тимпанальной поверхности базилярной мембраны. Основное значение в трофике спирального органа принадлежит сосудистой полоске, состоящей из трех видов эпителиальных клеток (маргинальных, интермедиарных и базальных), тесно связанных морфологически и функционально с эндотелиальными клетками. Эндолимфатическую поверхность сосудистой полоски составляют маргинальные клетки, цитоплазма которых насыщена органоидами, в особенности митохондриями. Детали этих клеток представляют собой сложнейшую разветвленную систему отростков, пронизывающих всю толщу сосудистой полоски и, как щупальцами, охватывающую капилляры. Энзиматическая активность сосудистой полоски чрезвычайно высока, в особенности ферментов окислительного метаболизма. По современным представлениям, stria vascularis обеспечивает насыщение эндолимфы кислородом, создает в улитке постоянный потенциал покоя и определяет состав эндолимфатической жидкости, в частности своеобразное «интрацеллюлярное» распределение в этой среде ионов калия и натрия.
Нарушение ионного равновесия в эндо- и перилимфе ведет к утрате слуховой функции. Конгенитальная патология сосудистой полоски лежит в основе врожденной глухонемоты, ее экспериментальное повреждение вызывает глубокое нарушение функции спирального органа, вплоть до его гибели.
Эндолимфатическая жидкость резорбцируется в эндолимфатическом мешке. Она изоосмотична с перилимфой, хотя и имеет отличный от нее количественный состав. Обмен между жидкостями возможен, главным: образом, через рейсснерову мембрану. В происхождении перилимфы основное значение принадлежит внутрилабиринтным источникам — процессу ультрафильтрации из сосудистых зон. Обе жидкости функционально едины и представляют собой целостную жидкостную систему внутреннего уха.
Нарушения циркуляции, химического состава, давления лабиринтной лимфы лежат в основе многих заболеваний, в частности болезни Меньера, кохлеарного неврита, старческой тугоухости, акустической травмы, интоксикаций и др. Гомеостаз внутренних лабиринтных сред обеспечивается функциональной активностью гематолабиринтного барьера. По сравнению с гематоэнцефалическим и гематоофтальчическим барьерами стабильность этого гистогематического барьера весьма высока, он является преградой для многих медикаментов, сохраняет свою инертность при значительных общих расстройствах гемодинамики. Селективное нарушение проницаемости барьера наступает при терапии антибиотиками группы аминогликозидов.
В физиологических условиях в наружных волосковых клетках возникает микрофонный потенциал — переменный биоток с частотой, синхронной тону озвучивания. Амплитуда потенциала в известных пределах пропорциональна интенсивности звукового раздражения. Форма звукового стимула в точности воспроизводится в форме волны микрофонного потенциала. Благодаря хорошей электропроводности лабиринтной лимфы регистрация микрофонного потенциала легко осуществима с круглого окна. Запись этого биотока (кохлеограмма) и ее анализ имели неоценимое значение для развития экспериментальной отологии и создания моделей заболеваний: акустической травмы, воспалительных процессов, гипоксии, оперативных вмешательств в области трансформационной системы среднего уха, воздействия медикаментов, включая анестетики, антибиотики, диуретики и др.
В улитке возникают также и другие биоэлектрические явления. Суммационный потенциал отражает функциональную активность внутренних волосковых клеток, акционный потенциал — ганглиозных слуховых клеток, а эндолимфатический потенциал покоя — клеток сосудистой полоски. X. Дэвис (Н. Davis, 1958) полагает, что эндолимфатический потенциал играет роль усилителя микроэлектрических явлений в спиральном органе и определяет его колоссальную чувствительность. Акционный потенциал представляет собой первичную кодированную информацию о звуке.
Механизм раздражения спирального органа (Н. Davis, 1958) состоит в сгибании стереоцилий волосковых клеток. Это предположение оспаривается цитохимической теорией слуха Я. А. Винникова и Л. К. Титовой (1961), которые усматривают механизм раздражения волосковых клеток спирального органа в деполяризации мембраны волосков ацетилхолином, растворенным в эндолимфе.
Спиральный (кортиев) орган — самый сложный прибор рецепции млекопитающих. Чувствительность органа близка к предельной, так как он способен реагировать на колебательный процесс, приближающийся по своим параметрам к броуновскому движению. В нем осуществляется весьма совершенный первичный анализ звука, то есть математическое разложение сложного сигнала на составляющие.
Нейрональный отдел включает восходящий и нисходящий слуховые пути. Восходящий путь представляет совокупность связанных между собой слуховых ядер, располагающихся в определенной последовательности: спиральный ганглий, кохлеарные ядра, верхняя олива, задние бугры четверохолмия, внутреннее коленчатое тело, слуховая кора. Нисходящий путь начинается в слуховой коре и достигает верхнеоливарной области, откуда идет хорошо прослеженный оливокохлеарный пучок, заканчивающийся на телах внутренних и наружных волосковых клеток крупными темными нервными окончаниями.
В слуховом ганглии выделяют три вида нейронов, из которых первый имеет миелинизированную оболочку у деидритов, а дендриты двух других лишены ее. За счет нейронов первого типа осуществляется иннервация внутренних волосковых клеток (в пропорции 1:20). Нейроны второго и третьего типов иннервируют наружные волосковые клетки, причем каждый нейрон обоих типов связан с 10 чувствительными клетками (В. Kellerhals et al., 1967). Таким образом, на уровне рецептора образуются иинервациониые, частично накладывающиеся поля, которые обеспечивают постоянство афферентации в случае дегенерации как отдельных волосковых, так и ганглиозных клеток.
Слуховые нейроны 2-го порядка сосредоточены в группе кохлеарных ядер продолговатого мозга (переднее и заднее вентральное ядро и дорзальное кохлеарное ядро или слуховой бугорок). Именно на уровне 2-го нейрона происходит перекрест основной массы волокон афферентного слухового пути, главная часть которого продолжает свой ход в составе трапециевидного тела и достигает верхней оливы. Меньшая часть волокон 2-го нейрона следует до заднего двухолмия и даже внутреннего коленчатого тела.
Комплекс верхней оливы (3-й слуховой нейрон) включает помимо латеральной и медиальной оливы скопление периоливарных ядер. На этом нейрональном уровне происходит конвергенция слуховых путей, подвергшихся или не подвергшихся ранее перекресту. Аксоны оливарных ядер и частично трапециевидного тела образуют латеральную петлю, достигающую заднего четверохолмия.
Задние бугры четверохолмия (заднее двухолмие) содержат в основном нейроны 4-го порядка, аксоны которых образуют пучок — ручку заднего бугра, которая и достигает внутреннего коленчатого тела ипсилатерально. Однако часть волокон переходит и на контралатеральную сторону. Аксоны нейронов 5-го порядка медиального коленчатого тела достигают височной коры (поля 41, 42 по Бродману у человека), где имеется шесть слоев клеток.
Функциональное значение нисходящего слухового пути изучено слабо. Считается (J. Fex, 1962), что оливокохлеарный пучок оказывает тормозное влияние в слуховой системе, способствует дифференцировке звуковых стимулов, уменьшению эффектов маскировки.
Биотоки улитки. Потенциал покоя и возбуждения улитки
Ткани и органы. Нервная ткань
Потенциал покоя и потенциал действия
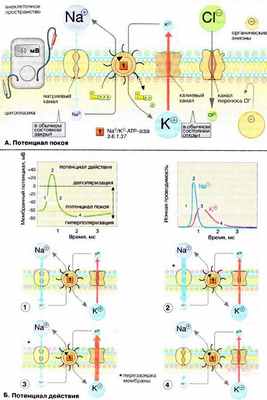
А. Потенциал покоя
Мембраны, в том чикле плазматические, в принципе непроницаемы для заряженных частиц. Правда, в мембране имеется Na + /K + -АТФ-аза (Nа + /К + -АТР-аза), осуществляющая активный перенос ионов Na + из клетки в обмен на ионы К + . Этот транспорт энергозависим и сопряжен с гидролизом АТФ (АТР) (см. рис. 221). За счет работы «Nа + ,К + -насоса» поддерживается неравновесное распределение ионов Na + и К + между клеткой и окружающей средой (см. с. 128). Поскольку расщепление одной молекулы АТФ обеспечивает перенос трех ионов Na + (из клетки) и двух ионов К + (в клетку), этот транспорт электрогенен, т. е. цитоплазма клетки заряжена отрицательно по отношению к внеклеточному пространству.
Электрохимический потенциал. Содержимое клетки заряжено отрицательно по отношению к внеклеточному пространству. Основная причина возникновения на мембране электрического потенциала (мембранного потенциала Δψ, см. c. 128) — существование специфических ионных каналов. Транспорт ионов через каналы происходит по градиенту концентрации или под действием мембранного потенциала. В невозбужденной клетке часть К + -каналов находится в открытом состоянии и ионы К + постоянно диффундируют из нейрона в окружающую среду (по градиенту концентрации). Покидая клетку, ионы К + уносят положительный заряд, что создает потенциал покоя равный примерно -60 мВ. Из коэффициентов проницаемости различных ионов (см. таблицу на рис. 129) видно, что каналы, проницаемые для Na + и Cl - , преимущественно закрыты. Ионы фосфата и органические анионы, например белки, практически не могут проходить через мембраны. С помощью уравнения Нернста (см. рис. 129) можно показать, что мембранный потенциал нервной клетки в первую очередь определяется ионами К + , которые вносят основной вклад в проводимость мембраны.
Ионные каналы. В мембранах нервной клетки имеются каналы, проницаемые для ионов Na + , К + , Са 2+ и Cl - . Эти каналы чаще всего находятся в закрытом состоянии и открываются лишь на короткое время. Каналы подразделяются на потенциал-управляемые (или электровозбудимые), например быстрые Na + -каналы, и лиганд-управляемые (или хемовозбудимые), например никотиновые холинэргические рецепторы. Каналы — это интегральные мембранные белки, состоящие из многих субъединиц. В зависимости от изменения мембранного потенциала или взаимодействия с соответствующими лигандами, нейромедиаторами и нейромодуляторами (см. рис. 343), белки-рецепторы могут находиться в одном их двух конформационных состояний, что и определяет проницаемость канала («открыт» — «закрыт» — и т.д.).
Б. Потенциал действия
Возбуждение нервной клетки под действием химического сигнала (реже электрического импульса) приводит к возникновению потенциала действия . Это означает, что потенциал покоя -60 мВ скачком изменяется на +30 мВ и спустя 1 мс принимает исходное значение. Процесс начинается с открывания Nа + -канала ( 1 ). Ионы Na + устремляются в клетку (по градиенту концентрации), что вызывает локальное обращение знака мембранного потенциала ( 2 ). При этом Na + -каналы тотчас закрываются, т. е. поток ионов Na + в клетку длится очень короткое время ( 3 ). В связи с изменением мембранного потенциала открываются (на несколько мс) потенциал-управляемые К + -каналы ( 2 ) и ионы К + устремляются в обратном направлении, из клетки. В результате мембранный потенциал принимает первоначальное значение ( 3 ), и даже превышает на короткое время потенциал покоя ( 4 ). После этого нервная клетка вновь становится возбудимой.
За один импульс через мембрану проходит небольшая часть ионов Na + и К + , и концентрационные градиенты обоих ионов сохраняются (в клетке выше уровень К + , а вне клетки выше уровень Na + ). Поэтому по мере получения клеткой новых импульсов процесс локального обращения знака мембранного потенциала может повторяться многократно. Распространение потенциала действия по поверхности нервной клетки основано на том, что локальное обращение мембранного потенциала стимулирует открывание соседних потенциал-управляемых ионных каналов, в результате чего возбуждение распространяется в виде деполяризационной волны на всю клетку.
Биотоки улитки. Потенциал покоя и возбуждения улитки
Биотоки улитки. Потенциал покоя и возбуждения улитки
Ввиду тех затруднений, с которыми встретилась теория пространственного расположения, некоторые авторы придерживаются дуалистического взгляда. Так, например, Ребул (Reboul, 1937) считает, что до 750 гц включительно основная мембрана колеблется целиком (наподобие телефонной мембраны) и соответствующая частота без искажения проводится до центров. Более высокие звуки (до 4000 гц) дают одно строго локализованное максимальное выпячивание. Уивер допускает, что ритмика звуков до 2000—3000 гц в неискаженном виде передается в центры. Правильная передача частот нарушается только для более высоких звуков, и для этой зоны пространственный фактор играет преобладающую роль.
Благодаря успехам электрофизиологии в настоящее время имеется возможность подойти к изучению процессов, совершающихся в нервных проводниках и корковых концах. При звуковом раздражении улитки в ней возникают электрические потенциалы — биотоки улитки, которые по силе, частоте и форме колебаний соответствуют звуковой кривой. Эти токи после усиления могут быть опять трансформированы (при помощи телефона или громкоговорителя) в звук и в точности повторяют поданный па ухо животного звук.
Токи улитки были получены у человека при помещении электрода в нишу круглого окна—через перфорацию барабанной перепонки или при операции (Г. В. Гершуни и др.).
Трактовка фактов, полученных путем регистрации токов улитки, затрудняется отчасти благодаря тому, что источник их не вполне выяснен. Имеются наблюдения, указывающие на то, что они возникают только при условии сохранности волосатых клеток (В. Ф. Ундриц и др.). Согласно новым данным Бекеши, мембраны перепончатой улитки поляризованы. При колебаниях этих структур возникают колебания электрических потенциалов. При повышении давления на овальное окно получается отрицательный заряд на мембране круглого окна. Высокая частота колебаний токов улитки объясняется суммарным действием многих клеток и волокон. Согласно теории залпов, суммарный эффект складывается из числа импульсов отдельного волокна (что соответствует частоте выстрелов из одного орудия) и количества одновременно работающих волокон (число орудий).
Троланд (Troland) допускает, что отдельные волокна обладают разными порогами возбудимости, почему начинают давать импульсы не одновременно, ввиду чего в единице времени получается суммация этих импульсов.
Большой интерес представляют новейшие данные, по которым в эндолимфатическом пространстве улитки даже вне деятельности существует потенциал покоя, который достигает огромной величины в 80 mV. Напрашивается мысль о значительной биологической роли высокого потенциала покоя в улитке. Наличие этого мощного усилителя, поставленного на месте трансформации звуковых колебаний в нервное возбуждение, вероятно, объясняет изумительную чувствительность звукового рецептора.
Токи центральных проводников отличается и их частота уменьшается при удалении от кортиева органа, и время возникновения все больше и больше запаздывает.
При помощи игольчатых электродов можно наблюдать первичный ответ на звуковое раздражение с различных участков звуковой зоны коры. У человека, как и у животных, электроэнцефалограмма с покровов черепа обнаруживает сложную кривую общей деятельности мозга, причем при звуковом раздражении наблюдается угнетение а-ритма и оживление бета-волн главным образом при височно-теменном отведении.
Для уточнения функции улитки, помимо изучения биотоков, применялось раздражение нервных элементов при помощи постоянного и переменного токов (А. М. Андреев, А. А. Волохов, Г. В. Гершуни). Хронаксия слухового нерва равняется 0,0001 секунды. При раздражении переменным током звуковой частоты наблюдаются звуковые ощущения, похожие на обычные (при адекватном раздражении). По-видимому, при этом возникают механические колебания в структурах уха (электрофонический эффект). При отсутствии кортиева органа раздражение ствола слухового нерва вызывает ощущение, резко отличное от звукового ощущения, получаемого при воздействии на кортиев орган. Это является новым доказательством о несостоятельности частотных теорий, т. е. теорий, которые связывают высоту воспринимаемого тона с частотой импульсов в нервных путях.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Гидродинамическая теория.
Исследования последнего времени показывают, что под влиянием звуков в лимфе улитки происходят сложные гидродинамические процессы. Это послужило основанием для создания Бекеши и Флетчером гидродинамической гипотезы слуха, которая значительно расширяет резонансную теорию Гельмгольца. Свои взгляды о механизме слуха Бекеши в I960 году изложи в виде теории "бегущей волны". Эта теория утверждает, что в ответ на звуковой раздражитель внутри улитки возникает бегущая волна, которая следует от основания к ее верхушке вдоль базиллярной мембраны. При этом колеблются не только волокна базиллярной мембраны, резонирующие на конкретную частоту, но и волокна на других участках мембраны. Расстояние, которое проходит бегущая волна по мембране, определяется частотой колебания стремени. Бегущая волна от высоких звуков проходит меньшее расстояние и обусловливает максимальную деформацию базиллярной мембраны, а следовательно, и максимальное раздражение волосковых клеток, преимущественно в области основного завитка улитки. Бегущая волна от низких звуков способна перемещаться на большие расстояния и таким образом вызывать деформацию мембраны по всей ее длине. Ощущение высоты звука определяется участком максимальной амплитуды колебаний базиллярной мембраны.
Поскольку базиллярная мембрана фиксирована по обоим краям, а покровная (текториальная) мембрана прикреплена только на одной стороне, по мнению Бекеши, одновременные движения обеих мембран вызывают скользящее действие на их соприкасающихся поверхностях. Это означает, что волосковые клетки подвергаются "режущемуся" воздействию текториальной мембраны. Сила такого воздействия зависит во многом от степени смещения мембран: чувствительные волосковые клетки подвергающиеся в этом случае максимальному смещению, вызывают наибольшее нервное возбуждение.
Таким образом, происходит трансформация механической энергии звуковых колебаний в нервное возбуждение, - на этот вопрос пытались и пытаются дать ответ многие исследователи. Значительный вклад в решение этой задачи сделан отечественными учеными. В основу электрофизиологического метода исследований данной проблемы положено учение Н.Е. Введенского о процессах нервного возбуждения. Согласно этим взглядам, ритм возбуждения нервной ткани соответствует ритму раздражения. В результате таких действий появляются электрические колебания, получившие название биотоков, или токов действия. Эти токи как оказалось, можно регистрировать. Оказалось, что улитка способна генерировать определённый переменный электрический потенциал в ответ на определенное звуковое раздражение.
В ответ на акустическую стимуляцию возникает пять электрических реакций, среди которых различают:
потенциал действия волокон VIIIнерва,
потенциал ствола мозга,
В 1958 г.JTasakiи соавторы сообщили, что улитка не ограничивается одной активностью в виде микрофонного потенциала, но обладает и другой активностью, генерируемой волосковыми клетками на акустический раздражитель - суммационным потенциалом. По мнению большинства авторов, суммационный потенциал, так же как и микрофонный, возникает в спиральном органе, однако его природа мало изучена.
Стимуляция улитки приводит к возникновению электрической энергии в волокнах слухового нерва. Эти потенциалы подобны тем, которые возникают в других нервах и названы потенциалами действия.
Применение электронной компьютерной техники в клинической практике сделало доступным регистрацию электрической активности улитки с помощью электродов, введенных в наружный слуховой проход, в область промонтория или окна улитки. Этот метод известен как электрокохлеография.
В последние годы благодаря успехам нейрофизиологии выявлен целый ряд новых данных, открывающих перспективу изучения сенсорных систем, в том числе и органа слуха.
До последнего времени считалось, что имеется один единственный (специфический) путь проведения импульсов: возбуждение начинается в рецепторе, затем распространяется на систему промежуточных нейронов и заканчивается локально в коре больших полушарий мозга.
Однако новейшие экспериментальные исследования показали, что этот классический путь проведения возбуждения не единственный, а существует и другой путь передачи возбуждения - неспецифический. Он обусловлен деятельностью ретикулярной формации. Доказано, что для возникновения слухового или любого другого ощущения недостаточно поступления в кору мозга импульсов, приходящих в неё по специфическому пути. Необходимо, чтобы эти импульсы были усилены волной неспецифического возбуждения из ретикулярной формации (Анохин П.К., 1968).Иными словами, в системе анализаторов не только имеет место односторонняя связь и передача импульсов от рецептора к клеткам головного мозга, но и существует центробежная ретикуляция возбудимости анализаторов, то есть происходит как восходящая (афферентная), так и нисходящая (эфферентная) передача импульсов от коры к периферическому отделу анализатора.
Таким образом, под влиянием деформации волосков рецепторных клеток при давлении покровной мембраны освобождается электрическая энергия синхронно со звуковыми колебаниями - эти потенциалы в виде микрофонных потенциалов отводятся наилучшим образом от самых волосковых клеток, а также от окна улитки благодаря электропроводимости ушной лимфы. Биотоки являются раздражителями тончайших окончаний веточек кохлеарного нерва, оплетающих волосковые клетки. Эти окончания имеют характер синапсов и возбуждение передаётся при помощи медиаторов (ацетилхолин). Следовательно, спиральный орган работает как детектор, отвечая только на определенный вид энергии (звук), и как трансформатор, превращая звуковую энергию в процесс нервного возбуждения.
Своими опытами по удалению височных долей мозга у собак И.П. Павлов установил, что отдельные элементы слухового анализатора рассеяны по всей коре больших полушарий и в них происходит низший анализ и синтез звуков. Однако высшие процессы анализа и синтеза звуковых раздражений возможны только в центре слухового анализатора, находящегося в коре височных долей мозга.
Физиология звуковосприятия
Звуковосприятие начинается с рецепторных клеток кортиева органа, которые, являясь вторичночувствующими волосковыми механорецепторными клетками, преобразуют механические звуковые колебания в электрические нервные импульсы.
Звуковосприятию соответствуют понятия слухового анализатора (по И.П.Павлову) и слуховой сенсорной системы, объединяющие слуховые рецепторы, проводящие пути и слуховые центры различных уровней ЦНС, включая кору височной доли головного мозга.
Существуют различные теории слуха, объясняющие механизм звуковосприятия в спиральном органе - рецепторе слуховой системы.
1. Теории периферического анализа звука.
- резонансная теория (Гельмгольц, 1863)
бегущей волны (Бекеши, 1960)
столба жидкости (Роаф-Флетчер, 1930)
- теория Флока (1977)
- теория Ухтомского (1945)
2. Теории центрального анализа звука.
- телефонная (Резерфорд, 1886)
- стоячих волн (Эвальд, 1899)
3. Дуалистическая теория (Ребул, 1938)
Теории периферического анализа звука предполагают возможность первичного анализа его свойств в улитке, благодаря ее анатомо-функциональным особенностям.
Резонаторная теория Гельмгольца заключается в том, что базилярная мембрана представляет собой набор "струн" разной длины и натянутости подобно музыкальному инструменту (например, роялю). "Струны" резонируют и реагируют на соответствующие им частоты звуковой волны, как, например, открытый рояль на человеческий голос. Теория Гельмгольца подтверждается морфологическим строением основной мембраны - у основания улитки струны короче (0,16мм), резонируют на высокие звуки, а у верхушки - они длиннее (0,52 мм) и реагируют на низкочастотные сигналы. При подаче сложных звуков одновременно колеблется несколько участков основной мембраны, чем объясняется тембр. От амплитуды колебаний мембраны зависит сила восприятия звука.
Теория Гельмгольца впервые позволила объяснить основные ñâîeñòâà oõà – îï?aäaëaièa âûñîòû, ñèëû è òaìa?à çâoêà, но она не объясняет явление маскировки высоких звуков низкочастотными звуками. Вместе с тем, современные знания не подтверждают возможность колебания отдельных “струн” основной мембраны, как и наличие их огромного числа на мембране длиной 35 мм, воспринимающей частоты в диапазоне 0,2-20 кГц.
По гидродинамической теории Бекеши звуковая волна, проходя в перилимфе обеих лестниц, вызывает колебания основной мембраны в виде бегущей волны. В зависимости от частоты звука происходит максимальный изгиб мембраны на ограниченном её участке. Низкие звуки вызывают бегущую волну по всей длине основной мембраны, вызывая максимальное смещение её около верхушки улитки. Среднечастотные тоны максимально смещают середину основной мембраны, а высокие звуки - в области основного завитка спирального органа, где базилярная мембрана более упругая и эластичная.
Гидродинамическая теория Роафа - Флетчера, построенная на опытах Лутца с U - образными трубками и жидкостью, подтверждает выводы Бекеши о том, что звуковые волны с высокой частотой распространяются вблизи основного завитка улитки, а с низкой частотой - до геликотремы.
Флок (1977) считает, что в формировании частотной избирательности главную роль играет базилярная мембрана с наружными волосковыми клетками, а не внутренними, как думают многие авторы. Эти клетки имеют эфферентные связи. Цилии их расположены в виде жесткой на изгиб W-конструкции, поэтому любые изменения длины клетки под воздействием разницы потенциалов будут приводить к смещению базилярной мембраны. В структуре наружных волосковых клеток найдены актин и миозин, необходимые компоненты любой сократительной системы. Биоэлектрическая активность наружных волосковых клеток в механическом раскачивании базилярной мембраны подтверждена опытами W.Brownell, G.Bander et al.
(1985). В настоящее время имеются математические и физические модели гидродинамики улитки, включающие нелинейные и активные механизмы (Шупляков B.C. с соавт., 1987; Zwicker E.,1986).
Теория "физиологического резонанса клеток" Ухтомского заключается в неодинаковой физиологической лабильности волосковых клеток, которые избирательно реагируют на разные частоты звуковых волн. При большой лабильности волосковых клеток, они реагируют на высокочастотные звуки и наоборот, что напоминает физиологический резонанс.
Центральные теории Резерфорда и Эвальда в отличие от предыдущих отрицают возможность первичного анализа звука в улитке.
По телефонной теории Резерфорда основой передаточного механизма для всех частот является кортиева покрышка наподобие телефонной мембраны с микрофонным эффектом. При давлении на волосковые клетки мембрана передаёт микрофонные потенциалы в синхронные по частоте сигналы в центры головного мозга, где происходит их анализ. Игнорируется роль механических колебаний основной мембраны.
По теории Эвальда под влиянием звука на основной мембране устанавливаются "стоячие" волны наподобие фигур Хладни (звуковые образы), которые анализируются в мозговых центрах, как соответствующие разнообразные слуховые ощущения.
Дуалистическая теория Ребула состоит в попытке соединить пространственную теорию с телефонной теорией. По его мнению, низкочастотные звуки передаются сразу в высшие слуховые центры, а высокие звуки имеют свою точную локализацию в определённом месте основной мембраны. Это противоречит фактам, так как импульсы высших отделов ЦНС не соответствуют частоте и характеру звуковой волны. Например, частота тонов круглого окна улитки составляет 16000 Гц, слухового нерва - 3500 Гц, продолговатого мозга – 2500 Гц и слуховой коры головного мозга – 100 Гц.
В колебательном процессе и раскачивании основной мембраны, возможно, имеет значение и отоакустическая эмиссия (Kemp D., 1978; Kemp D., Chum R., 1980). Она состоит в генерации акустических сигналов в улитке без звуковой стимуляции или после неё, которые регистрируются с помощью миниатюрного и высокочувствительного низкошумящего микрофона в наружном слуховом проходе.
Эти сигналы - различные по частоте и форме волн у разных людей. Индивидуальная картина эмиссии может соответствовать ндивидуальным отклонениям аудиограммы. При патологии внутреннего уха меняются "пороги" эмиссии. Спонтанной эмиссии, возникающей без акустической стимуляции, приписывают роль в острой частотной настройке улитки и активности слуховых рецепторов. Возможно, осуществляется одновременное влияние бегущей волны и резонанса на основную мембрану за счёт отоакустической эмиссии.
В спиральном органе механический процесс звукопроведения трансформируется в электрофизиологический механизм звуковосприятия.
Во время функционального покоя между наружной и внутренней сторонами клеточной мембраны существует мембранный потенциал рецепторной клетки или потенциал покоя (ПП), обусловленный неравномерным распределением ионов натрия и калия между цитоплазмой и окружающей средой. Внутренняя сторона мембраны заряжена отрицательно по отношению к наружной. Максимальный потенциал покоя внутренних волосковых клеток –42 мВ в апикальной части улитки (Dallos P., 1985) и –55 мВ – в ее базальной части (Rassel J., 1985), à наружных волосковых клеток – соответственно –71 мВ и –100 мВ. Таким большим потенциалом покоя не обладают другие клетки организма.
Вибрация ушной лимфы вызывает смещение основной мембраны бегущей волной и сгибание стереоцилий волосковых клеток. В результате этого в апикальной части клетки происходит деполяризация ее мембраны. При уменьшении ПП на 10-20% возникает градуальный рецепторный потенциал (РП), который с дектрементом затухания распространяется к основанию клетки и приводит к выбросу медиатора в синаптическую щель. На постсинаптической мембране нервного окончания слухового нерва возникает генераторный потенциал (ГП). Его величина тоже прямо пропорциональна силе раздражителя. ГП электротонически достигает электрогенного участка постсинаптической мембраны нерва, на котором периодически, при достижении ГП критического уровня, возникает потенциал действия (ПД), именуемый во вторичночувствующих рецепторах возбудительным постсинаптическим потенциалом (ВПСП).
Он подчиняется закону “âña èëè iè÷aãî”. Амплитуда его выше амплитуды ПП нерва. Возникновение ПД связано с устремлением ионов натрия в клетку при ее возбуждении. Нисходящая фаза поляризации зависит от потока ионов калия из клетки и исходный заряд восстанавливается. Нервные импульсы по проводящим путям слуховой системы достигают коркового центра слуха височной доли мозга.
Кроме электрических потенциалов рецепторных клеток и слухового нерва, выделяют микрофонный, суммационный и эндокохлеарный электрические потенциалы улитки.
Микрофонный потенциал (МП) представляет собой переменные электрические колебания в жидких средах улитки, повторяющие форму звуковой волны. Впервые их зарегистрировали в 1930 г. Уивер и Брей (Wever E., Bray C), а в 1937 г. - Г.В.Гершуни и ВФ.Ундриц. Эти потенциалы отводятся электродами из улитки, от круглого окна или от стенки наружного слухового прохода, барабанной перепонки. Они отражают функцию волосковых клеток, больше наружных, и подобны техническому микрофону, преобразующему импульсы давления в электрические сигналы.
Суммационный потенциал (СП), связанный с акустической стимуляцией, является постоянным и не воспроизводит форму звуковой волны. Он имеет небольшую величину и не зависит от кислородного обеспечения улитки и других факторов, влияющих на микрофонный эффект.
Эндокохлеарный потенциал (ЭКП) или эндолимфатический потенциал покоя обнаружен Бекеши (Bekesy G., 1952). Он не зависит от стимуляции и составляет от +60 до +80 мВ. Тасаки (Tasaki J., 1960) связывает его с функцией сосудистой полоски. Из всех жидкостей лабиринта по химическому составу одна перилимфа сходна с другими внеклеточными жидкостями, в частности со спинномозговой жидкостью и сывороткой крови, так как перилимфатическое пространство посредством улиткового протока (ductus cochlearis) сообщается с субарахноидальным ликворным пространством. В перилимфе высокая концентрация ионов натрия и низкая – калия.
Эндолимфа резко отличается от других внеклеточных жидкостей, так как в ней высокий уровень содержания ионов калия и очень низкий – натрия. Благодаря такому ионному соотношению создается постоянный потенциал покоя, являющийся усилителем всех микроэлектрических процессов в спиральном органе. Так как спиральный орган не имеет сосудов, сосудистая полоска обеспечивает его трофику, насыщая эндолимфу кислородом. Кортилимфа, находящаяся в туннеле и омывающая волосковые клетки и их нервные окончания, богата натрием. Она изолирована от эндолимфы кутикулярной пластинкой. Источником образования кортилимфы, вероятно, является капиллярная сеть, расположенная под базилярной мембраной. Гомеостаз внутрилабиринтных сред зависит от состояния гематолабиринтного барьера.
По мнению В.О.Самойлова (1986), ЭКП возникает благодаря активному транспорту водородный ионов (Н+) в эндолимфу из матрикса митохондрий маргинальных клеток. ЭКП имеет важное значение для нормального функционирования слухового рецептора. Он поддерживает большую разность потенциалов (до 150 мВ) между эндолимфой и цитоплазмой волосковых клеток у основания стереоцилий. Эта разность потенциалов возникает от суммирования положительного ЭКП с отрицатеьным потенциалом покоя рецепторной клетки. Падение его наблюдается при нарушении кислородного снабжения сосудистой полоски (Ланцов А.А., 1982).
Внутренние волосковые клетки (3500–4000) имеют связи преимущественно с афферентными нервными волокнами, а наружные (18000-20000) – c эфферентными волокнами. Слуховой нерв контактирует с волосковыми клетками на основной мембране через синапсы. От одной внутренней клетки отходит 10-20 афферентных волокон, а наружной - 1-2. Небольшая группа нервных волокон слухового нерва проводит импульсы преимущественно одной частоты. Если звуковой стимул представляет сложное колебание, то в слуховом нерве активируются все волокна, соответствующие спектру звука. На уровне слуховых рецепторов происходит частотный анализ звуков, а длительность их кодируется временем активации афферентных волокон слухового нерва.
В гуморальной регуляции функции кортиева органа определенное значение имеют особые клетки сосудистой полоски – апудоциты, являющиеся элементами системы эндокринной клеточной регуляции. Апудоциты продуцируют биогенные амины – серотонин, мелатонин и пептидные гормоны – адреналин, норадреналин.
Функция подкорковых слуховых центров изучена сравнительно мало. Через них осуществляется безусловная рефлекторная связь с двигательными реакциями на звук (повороты головы, глаз), кохлео-пальпебральный рефлекс Бехтерева, кохлео-пупиллярный рефлекс Шурыгина и др.
Основные сведения по локализации корковых центров получены при помощи условных рефлексов, опытов с экстирпацией коры и отведением биотоков. При поражении корковых центров слуха нарушается высший анализ звуковых сигналов, синтез их в слитный образ, плохая разборчивость речи при удовлетворительном восприятии чистых тонов (Ундриц В.Ф., 1923).
Роль высших центров слуха подтверждается в клинике, когда после тимпанопластики улучшается острота слуха не только на оперированное ухо, но и на другое за счёт снятия торможения слухой зоны коры (Хилов К.Л.; Белов И.М., 1963 /. Тесно связаны с функцией слуховых центров височной коры такие физиологические свойства слуховой системы, как бинауральный пространственный слух, адаптация, маскировка и др.
Пространственный слух, бинауральный слух и ототопика взамосвязаны (Руденко В.П., 1967; Альтман Я.А., 1981).Пространственный слух – способность локализовать и определять направление звука в пространстве, что связано с бинауральным слухом, основанном на двустороннем частичном перекресте слуховых путей. Пространственный слух обусловливается временем или интенсивностью поступления звука в каждое ухо. Со стороны какого уха звук будет интенсивнее или быстрее поступит в слуховую систему, с той стороны и будет локализоваться источник сигнала. Здесь имеет значение расстояние от источника звука до каждого уха, а также фаза и угол звукового луча.
Маскировкой называется явление, заключающееся в увеличении порога тестового сигнала в присутствии другого звука (маскера). Маскировка широко применяется в аудиометрии с целью заглушения лучше слышащего уха. Вследствие связей каждой улитки с обоими высшими центрами слуха часто отмечается маскирующее действие одних звуков на другие в окружающей обстановке. Действие маскера зависит от частоты и интенсивности сигнала. Низкие и более сильные звуки обладают большей маскировкой.
Под слуховой адаптацией понимают временное повышение слуховых порогов в результате звуковой стимуляции. Это приспособительная и защитная реакции. В условиях тишины или при действии очень слабых звуков чувствительность органа слуха может повышаться, что В.Ф.Ундриц (1962) назвал сенсибилизацией чувствительности. Адаптационная способность уха зависит от состояния центральных и периферических отделов слуховой системы, уравновешенности процессов возбуждения и торможения в коре головного мозга (Вартанян И.А., 1981). Утомление слуховой функции является патологической реакцией, теряющей при длительном (несколько месяцев) действии раздражителя способность обратного развития npoцecca. В кортиевом органе появляются необратимые изменения, приводящие к тугоухости.
Слух для человека является биологическим и социальным фактором развития речи и речевого общения. Центральным звеном всего аппарата речи является кора головного мозга, преимущественно левого полушария, где у правшей находятся речеслуховой и кинестезический центры. Физиологическое восприятие речи осуществляется слуховой и зрительной системами. Слуховая система контролирует интесивность, частоту, тембр и другие качества речи. Речевые сигналы - это совокупность элементов акустической энергии с быстро меняющимися амплитудами и частотами. Средняя частота основного тона речи у мужчин составляет 136 Гц, у женщин - 248 Гц, то есть на октаву выше (Ермолаев В.Г. с соавт. ,1970). Обычная речь у людей имеет частотный диапазон в пределах одной октавы, а у певцов, артистов - до двух октав. Встречаются певцы, имеющие звуковысотный диапазон до трёх и даже четырёх октав (Има Сумак, Маро Робен). Речевые сигналы ограничены частотным диапазоном 1000 - 10000 Гц и интесивностью от 50 до 80 дБ.
Чтобы услышать речь, её уровень должен быть выше порога слышимости или порога обнаружения. Только в этом случае человек начинает различать отдельные слова. С повышением интенсивности речевых сигналов разборчивость их увеличивается. Пороги разборчивости зависят от фонемного состава речевого материала, количества слогов в слове, частоты их употребления, а также наличия и характера шумовой помехи. Примером тесной взаимосвязи слуховой и речевой функций является опыт Ломбарда, когда при чтении текста вставляют в уши трещотки Барани и интенсивность речи резко возрастает, так как человек должен слышать и постоянно контролировать свою речь. С этой же целью в шумной обстановке люди говорят громче.
Следовательно, слуховой орган позволяет человеку воспринимать и адекватно реагировать на звуковые изменения окружаюжей среды. Каждому участку слуховой системы свойственна определённая функция, нарушение которой ведёт к частичной или полной потере слуха.
Читайте также: