Физиология и функции ретикулярной формации
Добавил пользователь Morpheus Обновлено: 29.01.2026
6. Роль ретикулярной формации мозгового ствола в регуляции функций. Варолиев мост
Ретикулярная формация (сетчатое образование) представляет собой скопление нейронов, начинающееся на уровне продолговатого мозга и продолжающееся на протяжении всего ствола мозга. В ретикулярной формации выделяют нисходящую часть , которая снижает деятельность спинного мозга и мышечный тонус (передняя часть продолговатого мозга и средняя часть моста), и восходящую часть – структуры ствола, среднего и промежуточного мозга – оказывает общее активирующее влияние на вышележащие отделы ЦНС; ей принадлежит важнейшая роль в регуляции уровня бодрствования и организации поведенческих реакций.
Нейроны ретикулярной формации имеют длинные маловетвящиеся дендриты и хорошо ветвящийся аксон, который образует синапсы более чем с 25 тысячами нейронов. В ретикулярную формацию поступают афферентные окончания:
- от температурных и болевых рецепторов,
- от сенсорной и частично от других зон коры,
- от ядер мозжечка.
Эфферентные волокна из ретикулярной формации идут:
- в спинной мозг (нисходящий путь),
- к неспецифическим ядрам таламуса, заднему гипоталамусу, полосатому телу (восходящие пути),
Ретикулярная формация координирует функционирование двигательных ядер черепных нервов, моторных спинальных центров и активность мышечных рецепторов.
Нисходящее влияние ретикулярной формации проявляется в центральном торможении рефлексов спинного мозга. Восходящее ее влияние может быть и активирующим, и тормозным. Активирующее влияние проявляется в том, что стимуляция ретикулярной формации приводит к активации коры больших полушарий и переходу из состояния сна в состояние бодрствования (пробуждению), тогда как тормозящее влияние переводит организм из состояния бодрствования в сон. Таким образом, важнейшей функцией восходящей части ретикулярной формации является регуляция цикла сон – бодрствование.
Кроме этого, ретикулярная формация поддерживает тонус вегетативных центров, интегрирует влияние симпатической и парасимпатической систем для обеспечения потребностей всего организма, передает модулирующие влияния от гипоталамуса к внутренним органам, содержит в себе такие жизненно важные центры, как сосудодвигательный, и дыхательный центр (в продолговатом мозге), пневмотаксический центр, осуществляющий переключение вдоха на выдох и наоборот (в среднем мозге).
Мост (варолиев мост) располагается выше продолговатого мозга.
Функции моста:
1- Сенсорные функции обеспечиваются:
- улитковыми и преддверными (треугольное, латеральное – Дейтерса, верхнее – Бехтерева) ядрами преддверно-улиткового нерва (первичный анализ вестибулярных раздражений, их силы и направленности);
- чувствительным ядром тройничного нерва (сигналы от рецепторов кожи лица, передних отделов волосистой части головы, слизистой оболочки носа и рта, конъюнктивы глазного яблока).
2 – Двигательные функции обеспечиваются:
- двигательным ядром тройничного нерва (V) – иннервирует жевательные мышцы, мышцы, натягивающие барабанную перепонку, мышцу, натягивающую небную занавеску;
- лицевой нерв (VII) иннервирует все мимические мышцы лица;
- отводящий нерв (VI) иннервирует прямую латеральную мышцу, отводящую глазное яблоко кнаружи.
3 – Проводящие функции:
- в покрышке моста – длинный медиальный и тектоспинальный пути;
- переднее и заднее ядро трапециевидного тела и латеральной петли обеспечивают первичный анализ информации от органов слуха и затем передают ее в задние бугры четверохолмий.
Функции ретикулярной формации ствола мозга
Ретикулярной формацией (РФ) называется сеть нейронов различных типов и размеров, имеющих многочисленные связи между собой, а также со всеми структурами ЦНС. Она располагается в толще серого вещества продолговатого, среднего и промежуточного мозга и регулирует уровень функциональной активности (возбудимости) всех нервных центров этих отделов ЦНС. Таким же образом она влияет на КБП.
В ЦНС выделяют две подсистемы, выполняющие разные организующие функции: специфическую и неспецифическую. Первая обеспечивает восприятие, проведение, анализ и синтез сигналов специфической чувствительности. К ним относятся все ее виды, т.е. зрительная, слуховая, болевая и т.д.
Неспецифической подсистемой является РФ. Она оказывает генерализованное возбуждающее или тормозящее влияние на многие структуры мозга. Следовательно, она может регулировать уровень функциональной активности моторной, сенсорной, висцеральных систем и организма в целом. Когда нервные импульсы идут по специфическим проводящим путям, по коллатералям этих путей они поступают и к нейронам РФ. Это приводит к их диффузному возбуждению. От нейронов РФ возбуждение передается на кору, что сопровождается возбуждением нейронов всех ее зон и слоев. Благодаря этому восходящему активирующему влиянию РФ, повышается активность аналитико-синтетической деятельности, увеличивается скорость рефлексов, организм подготавливается к реакции на неожиданную ситуацию. Поэтому РФ участвует в организации оборонительного, полового, пищедобывательного поведения. С другой стороны, она может избирательно активировать или тормозить определенные системы мозга. В свою очередь кора больших полушарий, через нисходящие пути, может оказывать возбуждающее действие на РФ.
Нисходящие ретикулоспинальные пути идут от РФ к нейронам спинного мозга. Поэтому она может оказывать нисходящие возбуждающие и тормозящие влияния на его нейроны. Например, ее гипоталамические и мезэнцефальные отделы повышают активность -мотонейронов спинного мозга. В результате этого растет тонус скелетных мышц, усиливаются двигательные рефлексы. Тормозящее влияние РФ на спинальные двигательные центры осуществляется через тормозные нейроны Реншоу. Это приводит к торможению спинальных рефлексов.
РФ контролирует передачу сенсорной информации через продолговатый, средний мозг, а также ядра таламуса. Она непосредственно участвует в регуляции бодрствования и сна, за счет синхронизирующих центров сна и бодрствования, находящихся в ней. На нейроны РФ оказывают влияние различные фармакологические вещества: амфетамины, кофеин, LSD-25, морфин (опыт Эдисона).
Функции мозжечка
Мозжечок состоит из 2-х полушарий и червя между ними. Серое вещество образует кору и ядра. Белое образовано отростками нейронов. Мозжечок получает афферентные нервные импульсы от тактильных рецепторов, рецепторов вестибулярного аппарата, проприорецепторов мышц и сухожилий, а также двигательных зон коры. Эфферентные импульсы от мозжечка идут к красному ядру среднего, ядру Дейтерса продолговатого мозга, к таламусу, а затем моторным зонам КБП и подкорковым ядрам.
Общей функцией мозжечка является регуляция позы и движений. Эту функцию он осуществляет путем координации активности других двигательных центров: вестибулярных ядер, красного ядра, пирамидных нейронов коры. Поэтому он выполняет следующие двигательные функции: 1. Регуляцию мышечного тонуса и позы.
2. Коррекцию медленных целенаправленных движений в ходе их выполнения, а также координацию этих движений с рефлексами положения тела.
3. Контроль за правильным выполнением быстрых движений, осуществляемых корой.
В связи с тем, что мозжечок выполняет данные функции, при его удалении у животного развивается комплекс двигательных нарушений, называемый триадой Лючиани. Он включает:
1. Атония и дистония - снижение и неправильное распределение тонуса скелетных мышц.
2. Астазия - невозможность слитного сокращения мышц, а как следствие, сохранения устойчивого положения тела при стоянии, сидении (покачивание).
3. Астения - быстрая утомляемость мышц.
4. Атаксия - плохая координация движений при ходьбе. Неустойчивая "пьяная" походка.
5. Адиадохокинез - нарушение правильной последовательности быстрых целенаправленных движений.
В клинике умеренные поражения мозжечка проявляются триадой Шарко:
1. Нистагм глаз в состоянии покоя.
2. Тремор конечностей, возникающий при их движениях.
3. Дизартрия - нарушения речи.
Л.А.Орбели установил, что мозжечок влияет и на различные вегетативные функции. Эти влияния могут быть возбуждающими и тормозящими. Например, при раздражении мозжечка увеличивается или снижается кровяное давление, изменяется частота сердцебиений, дыхание, пищеварение. Мозжечок влияет на обмен веществ. На эти функции он воздействует через вегетативные нервные центры, координируя их активность с движением. Функции внутренних органов изменяются в связи с изменением обменных процессов в них. Поэтому мозжечок оказывает на них адаптационно-трофическое влияние.
Ретикулярная формация ствола мозга
Ретикулярная формация - совокупность различных нейронов, расположенных на протяжении ствола мозга, оказывающих активирующее или тормозящее влияние на различные структуры центральной нервной системы, тем самым контролируя их рефлекторную деятельность.
Ретикулярная формация ствола мозга оказывает активирующее влияние на клетки коры головного мозга и тормозное действие на мотонейроны спинного мозга. Посылая в спинной мозг к его двигательным нейронам тормозящие и возбуждающие импульсы ретикулярная формация участвует в регуляции тонуса скелетных мышц.
Ретикулярная формация поддерживает тонус вегетативных центров, интегрирует симпатические и парасимпатические влияния, передает модулирующее влияние от гипоталамуса и мозжечка к внутренним органам.
Функции ретикулярной формации
Соматодвигательный контроль (активация скелетной мускулатуры), может быть прямым через tr. reticulospinalis и непрямым через мозжечок, оливы, бугорки четверохолмия, красное ядро, черное вещество, полосатое тело, ядра таламуса и даже соматомоторные зоны коры.
Соматочувствительный контроль, т.е. снижение уровней соматосенсорной информации — «медленная боль», модификация восприятия различных видов сенсорной чувствительности (слуха, зрения, вестибуляции, обоняния).
Висцеромоторный контроль состояния сердечно-сосудистой, дыхательной систем, активности гладкой мускулатуры различных внутренних органов.
Нейроэндокринная трансдукция через влияние на нейромедиаторы, центры гипоталамуса и далее гипофиз.
Биоритмы через связи с гипоталамусом и шишковидной железой.
Различные функциональные состояния организма (сон, пробуждение, состояние сознания, поведение) осуществляются посредством многочисленных связей ядер ретикулярной формации со всеми частями ЦНС.
Координация работы разных центров ствола мозга, обеспечивающих сложные висцеральные рефлекторные ответы (чихание, кашель, рвота, зевота, жевание, сосание, глотание и др.).
Строение ретикулярной формации
Ретикулярная формация образована совокупностью многочисленных нейронов, лежащих отдельно или сгруппированных в ядра (см. рис. 1 и 2). Ее структуры локализуются в центральных участках ствола, начиная с верхних сегментов шейного отдела спинного мозга до верхнего уровня ствола мозга, где они постепенно сливаются с ядерными группами таламуса. Ретикулярная формация занимает пространства между ядрами черепных нервов, другими ядрами и трактами, проходящими через ствол мозга.
Нейроны ретикулярной формации характеризуются большим разнообразием форм и размеров, но их общим признаком является то, что они образуют длинными дендритами и широко ветвящимися аксонами многочисленные синаптические контакты как между собой, так и с нейронами других ядер мозга. Эти ветвления формируют своеобразную сеть (ретикулум), откуда произошло название — ретикулярная формация. У нейронов, формирующих ядра ретикулярной формации, имеются длинные аксоны, образующие проводящие пути к спинному мозгу, ядрам ствола мозга, мозжечка, таламуса и других областей головного мозга.
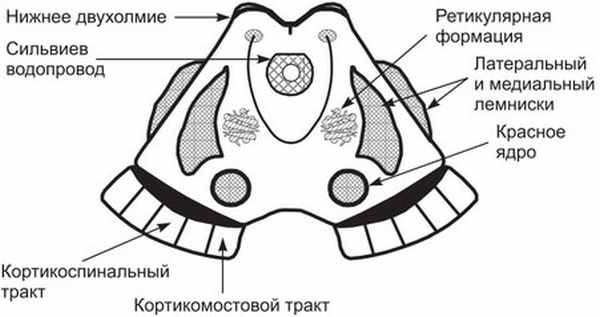
Рис. 1. Важнейшие структурные образования среднего мозга (поперечный срез)
К нейронам ретикулярной формации поступают многочисленные афферентные сигналы из различных структур ЦНС. Можно выделить несколько групп нейронов, к которым поступают эти сигналы. Это группа нейронов латерального ядра ретикулярной формации, расположенного в продолговатом мозге. Нейроны ядра получают афферентные сигналы от вставочных нейронов спинного мозга и входят в состав одного из непрямых спиномозжечковых путей. Кроме того, они получают сигналы от вестибулярных ядер и могут интегрировать информацию о состоянии активности вставочных нейронов, связанных с мотонейронами спинного мозга, и о положении тела и головы в пространстве.
Следующая группа — это нейроны ретикулотегментального ядра, расположенные на границе дорсального края моста. Они получают афферентные синаптические входы от нейронов претектальных ядер и верхних холмиков четверохолмия и посылают свои аксоны в структуры мозжечка, участвующие в контроле движений глаз.
Нейроны ретикулярной формации получают разнообразные сигналы через пути, связывающие их с корой головного мозга (кортикоретикулоспинальные пути), черной субстанцией, гипоталамусом и лимбической системой.
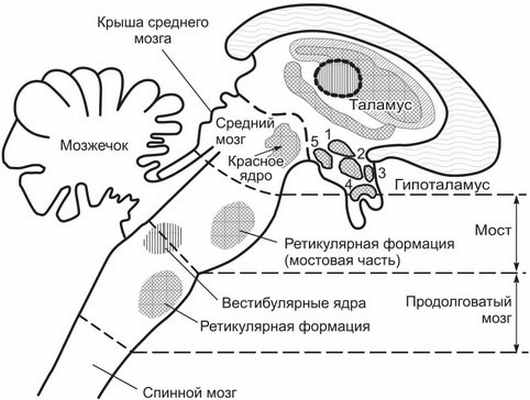
Рис. 2. Расположение некоторых ядер в стволе мозга и гипоталамусе: 1 — паравентрикулярное; 2 — дорсомедиальное: 3 — преоптическое; 4 — супраоптическое; 5 — заднее
Кроме описанных афферентных путей в ретикулярную формацию поступают сигналы по аксонным коллатералям проводящих путей сенсорных систем. При этом на один и тот же нейрон могут конвергировать сигналы от разных рецепторов (тактильных, зрительных, слуховых, вестибулярных, болевых, температурных, проприорецепторов, рецепторов внутренних органов).
Из приведенного перечня основных афферентных связей ретикулярной формации с другими областями ЦНС видно, что состояние ее тонической нейронной активности определяется притоком практически всех типов сенсорных сигналов от чувствительных нейронов, а также сигналов от большинства структур ЦНС.
Отделы
Характеристика
- дыхательный;
- сосудодвигательный;
- слюноотделительный и др.
- специфические центры, формирующие специфические ретикулоспинальные пути;
- неспецифические центры, формируют неспецифические ретикулоспинальные пути двух видов — активирующие, тормозные
Ретикулокортикальные: активирующие; гипногенные
Ядра ретикулярной формации и их функции
Долгое время считалось, что ретикулярная формация, строение которой характеризуется широкими межнейронными связями, интегрирует сигналы различной модальности, не выделяя при этом специфической информации. Однако становится все более очевидным, что ретикулярная формация является не только морфологически, но и функционально гетерогенной, хотя различия между функциями ее отдельных частей не столь очевидны, как это характерно для других областей мозга.
Действительно, многие нейронные группы ретикулярной формации формируют ее ядра (центры), выполняющие специфические функции. Это нейронные группы, формирующие сосудодвигательный центр продолговатого мозга (гигантоклеточное, парамедианное, латеральное, вентральное, каудальное ядра продолговатого мозга), дыхательный центр (гигантоклеточное, мелкоклеточное ядра продолговатого мозга, оральное и каудальное ядра моста), центры жевания и глотания (латеральное, парамедианное ядра продолговатого мозга), центры движений глаз (парамедианная часть моста, ростральная часть среднего мозга), центры регуляции тонуса мышц (ростральное ядро моста и каудальное — продолговатого мозга) и др.
Одной из важнейших неспецифических функций ретикулярной формации является регуляция общей нейронной активности коры и других структур ЦНС. В ретикулярной формации проводится оценка биологической значимости поступающих сенсорных сигналов, и в зависимости от результатов этой оценки она может активировать или тормозить через неспецифические или специфические нейронные группы таламуса нейронные процессы во всей коре головного мозга или в се отдельных зонах. Поэтому стволовая ретикулярная формация называется также активирующей системой ствола мозга. Благодаря этим свойствам ретикулярная формация может оказывать влияние на уровень общей активности коры, поддержание которой является важнейшим условием для сохранения сознания, состояния бодрствования, формирования направленности внимания.
Повышение активности ретикулярной формации (на общем высоком фоне) в отдельных сенсорных, ассоциативных областях коры обеспечивает возможность выделения и обработки специфической, наиболее важной в данный момент времени информации для организма и организации адекватных ответных поведенческих реакций. Обычно эти реакции, организуемые при участии ретикулярной формации ствола мозга, предваряются ориентационными движениями глаз, головы и тела в направлении источника сигнала, изменениями дыхания и кровообращения.
Активирующее влияние ретикулярной формации на кору и другие структуры ЦНС осуществляется по восходящим путям, идущим от гигантоклеточного, латерального и вентрального ретикулярных ядер продолговатого мозга, а также от ядер моста и среднего мозга. По этим путям потоки нервных импульсов проводятся к нейронам неспецифических ядер таламуса и после их обработки переключаются в таламических ядрах для последующей передачи к коре. Кроме того, от перечисленных ретикулярных ядер потоки сигналов проводятся к нейронам заднего гипоталамуса и базальных ганглиев.
Кроме регуляции нейронной активности высших отделов мозга ретикулярная формация может регулировать сенсорные функции. Это осуществляется путем влияния на проведение афферентных сигналов в нервные центры, на возбудимость нейронов нервных центров, а также на чувствительность рецепторов. Повышение активности ретикулярной формации сопровождается повышением активности нейронов симпатической нервной системы, иннервирующей органы чувств. В результате может повышаться острота зрения, слуха, тактильная чувствительность.
Наряду с восходящими активирующими и тормозящими влияниями на высшие отделы головного мозга ретикулярная формация принимает участие в регуляции движений, оказывая активирующие и тормозные воздействия на спинной мозг. На ее ядрах происходит переключение как восходящих путей, идущих от проприорецепторов и спинного мозга к головному мозгу, так и нисходящих двигательных путей от коры мозга, базальных ядер, мозжечка и красного ядра. Хотя восходящие нейронные пути, идущие из ретикулярной формации в таламус и кору, играют роль преимущественно в поддержании общего уровня активности коры больших полушарий мозга, но именно эта их функция важна для осуществления бодрствующей корой планирования, запуска, исполнения движений и контроля за их исполнением. Между восходящими и нисходящими через ретикулярную формацию путями имеется большое число коллатеральных связей, через которые они могут оказывать взаимное влияние. Существование такого тесного взаимодействия создает условия для взаимного воздействия области ретикулярной формации, влияющей через таламус на активность коры, планирующей и инициирующей движения, и области ретикулярной формации, влияющей на исполнительные нейронные механизмы спинного мозга. В ретикулярной формации имеются группы нейронов, которые посылают большинство аксонов в мозжечок, участвующий в регуляции и координации сложных движений.
По нисходящим ретикулоспинальным путям ретикулярная формация непосредственно воздействует на функции спинного мозга. Прямое влияние на его двигательные центры осуществляется по медиальному ретикулоспинальному тракту, идущему от ядер моста и активирующему преимущественно интер- и у-моторные нейроны разгибателей и тормозящему моторные нейроны мышц-сгибателей туловища и конечностей. По латеральному ретикулоспинальному тракту, начинающемуся от гигантоклеточного ядра продолговатого мозга, ретикулярная формация оказывает активирующее влияние на интер- и у-моторные нейроны мышц-сгибателей конечностей и тормозящее на нейроны мышц-разгибателей.
Из экспериментальных наблюдений на животных известно, что стимуляция более рострально расположенных нейронов ретикулярной формации на уровне продолговатого и среднего мозга, оказывает диффузное облегчающее влияние на спинальные рефлексы, а стимуляция нейронов каудальной части продолговатого мозга сопровождается торможением сиинальных рефлексов.
Активирующее и тормозное влияние ретикулярной формации на моторные центры спинного мозга может осуществляться через у-мотонейроны. При этом ретикулярные нейроны рострального участка ретикулярной формации активируют у-мотонейроны, которые своими аксонами иннервируют интрафузальные мышечные волокна, вызывают их сокращение, активируют рецепторы мышечных веретен. Поток сигналов от этих рецепторов активирует а-мотонейроны и вызывает сокращение соответствующей мышцы. Нейроны каудального участка ретикулярной формации тормозят активность у-мотонейронов спинного мозга и вызывают расслабление мышц. От баланса нейронной активности этих участков ретикулярной формации зависит распределение тонуса в больших мышечных группах. Поскольку этот баланс зависит от нисходящих влияний на ретикулярную формацию коры головного мозга, базальных ганглиев, гипоталамуса, мозжечка, то эти структуры мозга также могут через ретикулярную формацию и другие ядра ствола мозга воздействовать на распределение тонуса мышц и позу тела.
Широкое ветвление аксонов ретикулоспинальных путей в спинном мозге создает условия для влияния ретикулярной формации практически на все моторные нейроны и соответственно на состояние мышц различных частей тела. Такая особенность обеспечивает эффективное воздействие ретикулярной формации на рефлекторное распределение тонуса мышц, позу, ориентацию головы и тела в направлении действия внешних раздражителей и участие ретикулярной формации в осуществлении произвольных движений мышц проксимальных частей тела.
В центральной части ретикулярного гигантоклеточного ядра располагается участок, раздражение которого тормозит все двигательные рефлексы спинного мозга. Наличие такого торможения структур головного мозга на спинной мозг было открыто И.М. Сеченовым в опытах на лягушках. Суть опытов состояла в исследовании состояния рефлексов спинного мозга после перересечения ствола мозга на уровне промежуточного мозга и раздражении каудального участка разреза кристалликом поваренной соли. Оказалось, что двигательные спинальные рефлексы при раздражении не проявлялись или становились ослабленными и восстанавливались после устранения раздражения. Таким образом было впервые выявлено, что один нервный центр может тормозить активность другого. Это явление назвали центральным торможением.
Ретикулярная формация играет важную роль в регуляции не только соматических, но и вегетативных функций (ретикулярные ядра ствола мозга входят в структуру жизненно важных отделов дыхательного центра и центров регуляции кровообращения). Латеральная группа ретикулярных ядер моста и дорсолатеральное ядро покрышки формируют мочевыделительный центр моста. Аксоны нейронов ядер этого центра достигают преганглионарных нейронов крестцового отдела спинного мозга. Стимуляция нейронов этих ядер в мосту сопровождается сокращением мускулатуры стенки мочевого пузыря и мочевыделением.
В дорсолатеральном мосту расположено парабрахиальное ядро, на нейронах которого заканчиваются волокна сенсорных нейронов вкуса. Нейроны ядра, подобно нейронам голубоватого пятна и черной субстанции, содержат нейромеланин. Число таких нейронов в парабрахиальном ядре уменьшается при болезни Паркинсона. Нейроны парабрахиального ядра имеют связи с нейронами гипоталамуса, амигдалы, ядрами шва, одиночного тракта и другими ядрами ствола мозга. Предполагают, что парабрахиальные ядра имеют отношение к регуляции вегетативных функций и понижение их числа при паркинсонизме объясняет возникновение вегетативных нарушений при этом заболевании.
В опытах на животных было показано, что при раздражении некоторых локальных участков ретикулярных структур продолговатого мозга и моста можно вызвать торможение активности коры и сон. На ЭЭГ при этом возникают низкочастотные (1-4 Гц) волны. На основе описанных фактов считают, что важнейшими функциями восходящих влияний ретикулярной формации являются регуляция цикла сон — бодрствование и уровня сознания. Оказалось, что к формированию этих состояний имеют прямое отношение ряд ядер ретикулярной формации ствола мозга.
Так, с каждой стороны центрального шва моста располагаются парамедианные ретикулярные ядра, или ядра шва, содержащие серотонинергические нейроны. В каудальной части моста они включают нижнее центральное ядро, которое является продолжением ядра шва продолговатого мозга, а в ростральной части моста в состав ядер шва моста входит верхнее центральное ядро, называемое ядром Бехтерева, или срединным ядром шва.
В ростральной части моста на дорсальной стороне покрышки располагается группа ядер голубоватого пятна. В них имеется около 16 000-18 000 меланинсодержащих норадренергических нейронов, аксоны которых широко представлены в различных отделах ЦНС — гипоталамусе, гиппокампс, коре больших полушарий мозга, мозжечке и спинном мозге. Голубоватое пятно простирается в средний мозг, и его нейроны прослеживаются в ссром веществе околоводопроводного пространства. Число нейронов в ядрах голубоватого пятна уменьшается при паркинсонизме, болезни Альцгеймера и синдроме Дауна.
Как серотонинергические, так и норадреналинергические нейроны ретикулярной формации играют роль в контроле цикла сон — бодрствование. Подавление синтеза серотонина в ядрах шва ведет к развитию бессонницы. Предполагают, что серотонинергические нейроны являются частью нервной сети регуляции медленноволнового сна. При действии серотонина на нейроны голубоватого пятна возникает парадоксальный сон. Разрушение ядер голубоватого пятна у экспериментальных животных не ведет к развитию бессонницы, но вызывает на несколько недель исчезновение фазы парадоксального сна.
Функции ретикулярной формации
Ретикулярная формация расположена во внутренней части ствола на протяжении от нижних отделов промежуточного мозга до продолговатого мозга, где она проходит между двигательными и чувствительными ядрами.
Собственные ядра ретикулярной формации имеют низкую плотность нейронов, относительно высокий процент клеток нейроглии и слабо выраженные границы ядер, количество которых измеряется несколькими десятками. Большинство нейронов ретикулярной формации принимает коллатерали от нескольких сенсорных путей (полисенсорная конвергенция) и обладает тонической активностью покоя (5—10 импульсов в секунду), растущей в связи с поступлением афферентных сигналов. Аксоны нейронов ретикулярной формации обычно делятся на две ветви, следующие в восходящем и нисходящем направлениях. Восходящие влияния ретикулярной формации регулируют возбудимость нейронов коры в связи с циклом сон—бодрствование, они необходимы для сохранения сознания и восприятия сенсорных сигналов. Нисходящие влияния осуществляются посредством ретику- лоспинального тракта, входящего в состав медиального пути к спинному мозгу и регулирующего его двигательную функцию. Координационные центры ретикулярной формации обеспечивают согласование афферентной и эфферентной систем, участвующих в регуляции вегетативных функций (кровообращение, дыхание, пищеварение). Особенности нейронной организации ретикулярной формации
Афферентные сенсорные входы представлены волокнами спиноретикулярного тракта, образованного отростками нейронов задних рогов спинного мозга, коллатералями от аксонов интернейронов чувствительных ядер (тройничного, блуждающего, языкоглоточного, вестибулярного и слухового нервов), а также небольшим количеством волокон от других сенсорных проводящих путей, следующих через ствол. Остальные проекции на нейроны ретикулярной формации представлены окончаниями нейронов ядер мозжечка, гипоталамуса и коры, они регулируют активность нейронов ретикулярной формации.
Разные области и ядра ретикулярной формации образуют синаптические контакты между собой, как, например, каудальная область с ростральной, и могут реципрокно влиять друг на друга.
В дорсальном отделе продолговатого мозга находится ядро солитарного или одиночного тракта, получающее сенсорную информацию от внутренних органов. В составе волокон языкоглоточного и блуждающего нервов сюда поступают сигналы от периферических баро- и хеморецепторов. От нейронов ядра солитарного тракта отходят аксоны к парасимпатическим нейронам черепно-мозговых нервов (дорсальные ядра блуждающих нервов и обоюдные ядра) и к рострально-вентролатеральной области продолговатого мозга, регулирующей активность симпатических нейронов спинного мозга.
Вегетативные центры ретикулярной формации продолговатого мозга находятся под контролем гипоталамуса, который изменяет активность симпатического и парасимпатического отделов соответственно возникающим задачам, например поддержанию температуры тела, осуществлению оборонительного, пищевого или полового поведения. Гипоталамус обеспечивает функциональное объединение вегетативных центров ствола и спинного мозга в целостные системы, регулирующие функции кровообращения и дыхания, температуру тела. Гипоталамус подвержен влиянию лимбических структур мозга, обеспечивающих формирование эмоциональных реакций, при возникновении которых происходит комплексное изменение активности вегетативных центров, приспосабливающее внутренние процессы организма к решению текущих поведенческих задач.
В ретикулярной формации моста и продолговатого мозга располагаются структуры сосудодвигательного центра, состоящего из прессорного и депрес- сорного отделов. Прессорный отдел находится в рострально-вентральной области продолговатого мозга, где сосредоточены нейроны, имеющие решающее значение в повышении общей выходной активности симпатического отдела вегетативной нервной системы. Депрессорный отдел расположен в медиокаудальных отделах ствола, его активация сопровождается понижением тонуса симпатических нервов и одновременным повышением тонуса блуждающего нерва.
Поблизости от ядер одиночного тракта билатерально расположен дыхательный центр, состоящий из дорсальной и вентральной групп нейронов.
В зависимости от выполняемой функции нейроны дыхательного центра подразделяются на инспираторные, экспираторные, преинспираторные и постинспираторные. Инспираторные нейроны генерируют ритм дыхания, их нисходящие в спинной мозг аксоны возбуждают мотонейроны диафрагмы и наружных межреберных мышц, при сокращении которых происходит вдох. Экспираторные нейроны регулируют активность мотонейронов внутренних межреберных мышц, сокращения которых происходят при активном выдохе. Преинспираторные и постинспираторные нейроны регулируют активность инспираторных нейронов. Смена вдоха выдохом и наоборот происходит при участии нейронов пневмотаксического центра, расположенного в передней части моста. Нисходящие и восходящие влияния ретикулярной формации
Нисходящие влияния ретикулярной формации предназначены для регуляции рефлекторной деятельности спинного мозга (см. раздел 4.2.2). Восходящие дивергентные проекции ретикулярной формации к другим отделам головного мозга образованы ветвящимися аксонами моноаминергических нейронов среднего мозга, голубого пятна и ядер шва, а также холинергических нейронов. Метаботропное действие моноаминергических медиаторов влияет на биохимические процессы нейронов-мишеней, это действие проявляется медленно по сравнению с распространением сигналов, передающихся через синапсы с помощью ионотропных рецепторов. Результатом такого действия является модуляция функций многих нервных клеток, участвующих в формировании мотиваций, эмоций и целенаправленного поведения.
Норадренергические нейроны голубого пятна обладают фоновой активностью, изменяющейся в зависимости от сенсорной афферентации разных модальностей, а также под влиянием других центров ретикулярной формации, гипоталамуса и мозжечка. Аксоны нейронов голубого пятна проецируются почти во все области мозга и заканчиваются варикозными расширениями, из которых выделяется норадреналин, диффундирует через межклеточное пространство и через него действует на большие популяции нейронов.
Активность норадренергических нейронов необходима для поддержания бодрствования, при физиологическом повышении этой активности формируются двигательные и вегетативные компоненты апресс-реакции, чрезмерное повышение активности нейронов голубого пятна приводит к развитию панического синдрома, а при сниженной активности возникает депрессия.
Проекция нейронов черной субстанции среднего мозга к полосатому телу (нигростриатный путь) необходима для регуляции его активности при осуществлении произвольных движений. Дегенеративное поражение нейронов черной субстанции приводит к развитию болезни Паркинсона, проявляющейся возникновением ригидности мышц и тремором. Еще одна проекция дофаминергических нейронов направлена к структурам лимбической системы и фронтальной коре (мезолимбический и мезокортикальный пути), участвующим в образовании мотиваций и эмоций. Влияние дофамина на эти структуры усиливается при удовлетворении биологических потребностей, а переживание в этот момент положительных эмоций побуждает людей к повторению действий, приводящих к подобному результату. Роль дофамина состоит в модуляции возбудимости нейронов-мишеней лимбических структур и коры, чтобы их взаимодействие становилось оптимальным при создании программы поведения, а сами дофаминергические нейроны являются необходимым звеном образующихся поведенческих программ. При избытке дофамина вследствие патологически повышенной активности дофаминергических нейронов возникают грубые нарушения поведения и мышления, проявляющиеся, например, при заболевании шизофренией.
Нейроны ядер шва, синтезирующие серотонин, образуют проекции в спинной мозг, кору мозжечка, ядра таламуса и кору больших полушарий, что в совокупности создает восходящую и нисходящую системы, каждая из которых принимает участие в регуляции циклических процессов сна и бодрствования. Активация холинергических нейронов рострального отдела среднего мозга и гигантоклеточной области покрышки моста, образующих проекции к зрительному бугру и коре, необходима для поддержания сознания, переработки сенсорной и ассоциативной информации в коре и обеспечения познавательной интеллектуальной деятельности. Активность холинергических, норадренергических и серотонинергических нейронов в цикле сна и бодрствования периодически изменяется под влиянием супра- хиазмальных и вентромедиальных ядер гипоталамуса, играющих роль основных водителей суточной ритмики физиологических процессов.
РЕТИКУЛЯРНАЯ ФОРМАЦИЯ
В шейном отделе спинного мозга между боковыми и задними рогами имеются клетки с большим количеством отростков. Такие же клетки есть в продолговатом мозге, мосту, среднем и промежуточном мозге. Нейроны густо переплетаются между собой, поэтому структура похожа на сеть и называется сетчатой структурой, или ретикулярной формацией (рис. 11.8 [1] ). Между нейронами ретикулярной формации образуется особенно много контактов, один аксон на протяжении 2 см образует до 27 тыс. синапсов. Большинство нейронов имеют длинные дендриты и короткий аксон.
В ретикулярной формации человека выделяют 48 отдельных ядер. Наиболее крупное — гигантоклеточное ядро, которое составляет 2 /3 размера ретикулярной формации и отличается тем, что содержит гигантские нейроны, отсутствующие в других ядрах.
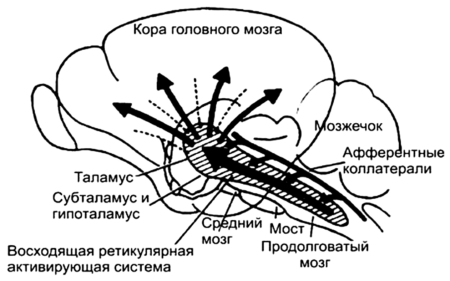
Рис. 11.8. Проекции восходящей ретикулярной активирующей системы
Сетевое строение обеспечивает высокую надежность функционирования ретикулярной формации, устойчивость к повреждениям, так как локальные повреждения всегда компенсируются за счет сохранившихся элементов сети. Высокая надежность функционирования ретикулярной формации проявляется в том, что раздражение любой ее части отражается на активности всей структуры за счет диффузности связей.
Основной функцией ретикулярной формации является регуляция уровня активности коры большого мозга, мозжечка, таламуса и спинного мозга.
В ретикулярной формации моста, продолговатого и среднего мозга имеются нейроны, которые реагируют на болевые раздражения, идущие от мышц или внутренних органов, что создает общее диффузное дискомфортное, не всегда четко локализованное болевое ощущение «тупой боли».
Ретикулярная формация ствола мозга принимает участие в регуляции мышечного тонуса, поскольку в сетевую структуру поступают нервные импульсы от зрительного и вестибулярного анализаторов и мозжечка. От ретикулярной формации к мотонейронам спинного мозга и ядрам черепных нервов поступают сигналы, обеспечивающие положение головы и туловища в пространстве.
К ядрам сетчатой структуры от всех афферентных систем по неспецифическому пути направляется поток чувствительных импульсов. Поэтому ретикулярная формация у бодрствующего человека постоянно находится в тонусе — возбуждении. От нее беспрерывно идут импульсы, которые обеспечивают оптимальный уровень функционирования коры головного мозга. Если фармакологически заблокировать ретикулярную формацию, то человек погружается в сон и кора перестает воспринимать раздражения из окружающего мира (зрение, слух).
Поток импульсов, идущий от ретикулярной формации к коре, называется «восходящей активирующей системой». Кора работает, только если к ней подходят импульсы от ретикулярной формации.
В ретикулярной формации заложены центры вегетативных функций (сердечно-сосудистой, дыхательной и пищеварительной систем).
Ретикулярная формация принимает участие в регуляции цикла «сон — бодрствование». При раздражении сетчатой структуры продолговатого мозга в ЭЭГ медленные волны с малой амплитудой, характеризующие состояние сна, сменяются быстрыми волнами, характеризующими состояние бодрствования.
В ретикулярной формации происходит взаимодействие афферентных и эфферентных импульсов, возможна их циркуляция по кольцевым связям, вследствие чего создается постоянный уровень возбуждения сетчатой структуры, благодаря которому обеспечиваются тонус и определенная степень готовности к деятельности различных отделов ЦНС.
Читайте также: