Гуморальная регуляция секреции пищеварительных соков и моторики желудка и кишечника. Гормональная регуляция пищеварительного тракта.
Добавил пользователь Владимир З. Обновлено: 30.01.2026
Гуморальная регуляция секреции пищеварительных соков и моторики желудка и кишечника. Гормональная регуляция пищеварительного тракта.
Центральные, периферические и местные рефлексы осуществляются в тесном взаимодействии с гуморальным механизмом регуляции миоцитов, гландулоцитов и нервных клеток.
В слизистой оболочке желудочно-кишечного тракта и в поджелудочной железе имеются эндокринные клетки, которые вырабатывают гастроинте-стинальные гормоны (регуляторные пептиды, энтерины). Эти гормоны через кровоток и местно (паракринно, диффундируя через межклеточную жидкость) оказывают влияние на миоциты, гландулоциты, интрамураль-ные нейроны и эндокринные клетки. Их выработка запускается рефлекторно (через блуждающий нерв) во время приема пищи и длительное время поддерживается за счет раздражающего влияния продуктов гидролиза пищевых веществ и экстрактивных веществ.
Таблица 11.1. Гормоны желудочно-кишечного тракта, место их образования и вызываемые ими эффекты
| Название гормона | Место выработки гормона | Типы эндокринных клеток | Эффект действия гормонов |
| Соматостатин | Желудок, проксимальный отдел тонкой кишки, поджелудочная железа | D-клетки | Тормозит выделение инсулина и глюкагона, большинства известных желудочно-кишечных гормонов (секретина, ГИПа, мотилина, гастрина); тормозит активность париетальных клеток желудка и ацинарных клеток поджелудочной железы |
| Вазоактивный интестинальный (ВИП) пептид | Во всех отделах желудочно-кишечного тракта | D-клетки | Тормозит действие холецистокинина, секрецию соляной кислоты и пепсина желудком, стимулированную гистамином, расслабляет гладкие мышцы кровеносных сосудов, желчного пузыря |
| Панкреатический полипептид (ПП) | Поджелудочная железа | D2-клетки | Антагонист ХЦК-ПЗ, усиливает пролиферацию слизистой оболочки тонкой кишки, поджелудочной железы и печени; участвует в регуляции обмена углеводов и липидов |
| Гастрин | Антральная часть желудка, поджелудочная железа, проксимальный отдел тонкой кишки | G-клетки | Стимулирует секрецию И выделение пепсина желудочными железами, возбуждает моторику расслабленного желудка и двенадцатиперстной кишки, а также желчного пузыря |
| Гастрон | Антральный отдел желудка | G- клетки | Снижает объем желудочной секреции и выход кислоты в желудочном соке |
| Бульбогастрон | Антральный отдел желудка | G- клетки | Тормозит секрецию и моторику желудка |
| Дуокринин | Антральный отдел желудка | G- клетки | Стимулирует выделение секрета бруннеровых желез двенадцатиперстной кишки |
| Бомбезин (гастринвысвобождающий пептид) | Желудок и проксимальный отдел тонкой кишки | Р-клетки | Стимулирует высвобождение гастрина, усиливает сокращение желчного пузыря и выделение ферментов поджелудочной железой, усиливает выделение энтероглюкагона |
| Секретин | Тонкий кишечник | S-клетки | Стимулирует секрецию бикарбонатов и воды поджелудочной железой, печенью, железами Бруннера, пепсина; тормозит секрецию в желудке |
| Холецистокинин-панкреозимин (ХЦК-ПЗ) | Тонкий кишечник | I-клетки | Возбуждает выход ферментов и в слабой степени стимулирует выход бикарбонатов поджелудочной железой, тормозит секрецию соляной кислоты в желудке, усиливает сокращение желчного пузыря и желчевыделение, усиливает моторику тонкой кишки |
| Энтероглюкагон | Тонкий кишечник | ЕС1-клетки | Тормозит секреторную активность желудка, снижает в желудочном соке содержание К+ и повышает содержание Са2+, тормозит моторику желудка и тонкой кишки |
| Мотилин | Проксимальный отдел тонкой кишки | ЕС2-клетки | Возбуждает секрецию пепсина желудком и секрецию поджелудочной железы, ускоряет эвакуацию содержимого желудка |
| Гастроингибирующий пептид (ГИП) | Тонкий кишечник | К-клетки | Тормозит выделение соляной кислоты и пепсина, высвобождение гастрина, моторику желудка, возбуждает секрецию толстой кишки |
| Нейротензин | Дистальный отдел тонкой кишки | N-клетки | Тормозит секрецию соляной кислоты железами желудка, усиливает высвобождение глюкагона |
| Энкефалины (эндорфины) | Проксимальный отдел тонкой кишки и поджелудочная железа | L-клетки | Тормозит секрецию ферментов поджелудочной железой, усиливает высвобождение гастрина, возбуждает моторику желудка |
| Субстанция Р | Тонкая кишка | ЕС1-клетки | Усиливает моторику кишечника, слюноотделение, тормозит высвобождение инсулина |
| Вилликинин | Двенадцатиперстная кишка | ЕС1-клетки | Стимулирует ритмические сокращения ворсинок тонкой кишки |
| Энтерогастрон | Двенадцатиперстная кишка | ЕС1-клетки | Тормозит секреторную активность и моторику желудка |
| Серотонин | Желудочно-кишечный тракт | ЕС1,ЕС2-клетки | Тормозит выделение соляной кислоты в желудке, стимулирует выделение пепсина, активирует секрецию поджелудочной железы, желчевыделение, кишечную секрецию |
| Гистамин | Желудочно-кишечный тракт | ЕС2-клетки | Стимулирует выделение секрета желудка и поджелудочной железы, расширяет кровеносные капилляры, оказывает активирующее влияние на моторику желудка и кишечника |
| Инсулин | Поджелудочная железа | Бета-клетки | Стимулирует транспорт веществ через клеточные мембраны, способствует утилизации глюкозы и образованию гликогена, тормозит липолиз, активирует липогенез, повышает интенсивность синтеза белка |
| Глюкагон | Поджелудочная железа | Альфа-клетки | Мобилизует углеводы, тормозит секрецию желудка и поджелудочной железы, тормозит моторику желудка и кишечника |
Место выработки основных гастроинтестинальных гормонов, вызываемые ими эффекты и продуцирующие их клетки представлены в табл. 11.1. В настоящее время обнаружено около 30 регуляторных пептидов. Как следует из представленной таблицы, они оказывают стимулирующее, тормозное и модулирующее влияние на секрецию пищеварительных соков, моторику гладкой мускулатуры желудочно-кишечного тракта, всасывание, выделение энтеринов эндокринными элементами слизистой оболочки желудка, кишечника и поджелудочной железы.
Выделение гастроинтестинальных гормонов имеет каскадный характер. Например, под влиянием гастрина обкладочные клетки желез желудка увеличивают выработку соляной кислоты, которая в слизистой оболочке тонкой кишки стимулирует выделение S- и J-клетками секретина и холеци-стокинна — панкреозимина. Секретин усиливает секрецию воды и бикарбонатов поджелудочной железой и печенью, а холецистокинин — панкреозимин — возбуждает выделение ферментов поджелудочной железой и тормозит секрецию соляной кислоты обкладочными клетками, усиливает моторику тонкой кишки и желчного пузыря.
Регуляторные пептиды, поступая в кровоток, быстро разрушаются в печени и почках и тем самым создают условия для осуществления эффектов других гастроинтестинальных гормонов.
Выработка некоторых энтеринов носит циклический характер и может осуществляться и при отсутствии пищевого раздражителя. Например, мо-тилин, вырабатываемый ЕС2-клетками проксимального отдела тонкой кишки, вызывает сокращения мышц желудка и кишечника, совпадающие с периодами «голодной» активности пищеварительного тракта.
Вопрос 4. Гуморальная регуляция жкт. Собственные гормоны жкт и их роль. Эффекты гастроинтестинальных гормонов.
В основном осуществляют гастроинтестиналъные гормоны: гастрин, секретин, холицистокинин, мотилин и др. (см. тему «Гуморальная регуляция»). Эти вещества продуцируются эндокринными клетками слизистой оболочки желудка, двенадцатиперстной кишки, поджелудочной железы, нейросекреторными клетками гипоталамуса и относятся к АПУД-системе. Гастроинтестинальные гормоны участвуют в регуляции секреции, моторики, всасывания, а также оказывают общие эффекты: изменения в обмене веществ, деятельности сердечно-сосудистой и эндокринной систем, пищевом поведении.
Гормоны поджелудочной железы
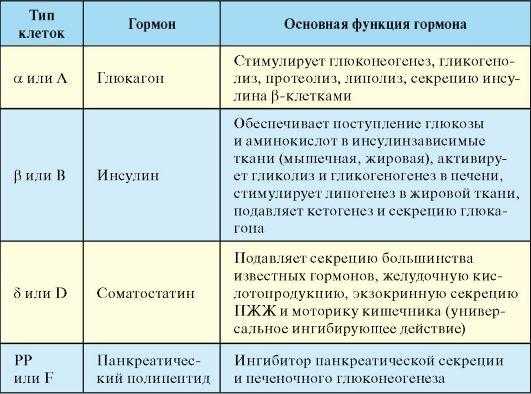
Эффекты гастроинтестинальных гормонов.
Место образования
Антральный отдел желудка и проксимальный отдел тонкой кишки (G-клетки)
Усиление секреции соляной кислоты и пепсиногена желудком и сока поджелудочной железы. Стимуляция моторики желудка, тонкой и толстой кишки, желчного пузыря.
Тонкая кишка, преимущественно в проксимальном отделе (S-клетки)
Увеличение секреции бикарбонатов поджелудочной железой, торможение секреции соляной кислоты в желудке, усиление желчеобразования и секреции тонкой кишки. Торможение моторики желудка, усиление моторики кишечника и сокращения пилорического сфинктера.
Тонкая кишка, преимущественно проксимальный отдел (1-клетки)
Усиление моторики желчного пузыря и секреции ферментов поджелудочной железой, торможение секреции соляной кислоты в желудке и его моторики, усиление секреции пепсиногена, стимуляция моторики тонкой и толстой кишки, расслабление сфинктера Одди. Угнетение аппетита.
Желудок и проксимальный отдел тонкой кишки (Р-клетки)
Стимуляция секреции желудка путем усиления высвобождения гастрина. Усиление сокращений желчного пузыря и секреции ферментов поджелудочной железой путем стимуляции высвобождения ХЦК-ПЗ, усиление высвобождения энтероглюкагона, нейротензина и ПП.
Желудок, тонкая кишка, преимущественно проксимальный отдел, (D-клетки) поджелудочная железа
Торможение выделения секретина, ГИПа, мотилина, гастрина, инсулина и глюкагона.
Тонкая кишка, преимущественно проксимальный отдел (ЕС2- клетки)
Усиление моторики желудка и тонкой кишки, усиление секреции пепсиногена желудком.
Панкреатический пептид (ПП)
Поджелудочная железа (ПП-клетки).
Антагонист ХЦК-ПЗ. Уменьшение секреции ферментов ибикарбонатов поджелудочной железой, усиление пролиферации тонкой кишки, поджелудочной железы и печени, усиление моторики желудка. Участие в обмене углеводов и липидов слизистой оболочки.
Желудочно-кишечный тракт (ECL-клетки)
Стимуляция секреции соляной кислоты желудком, сока поджелудочной железы. Усиление моторики желудка и кишечника. Расширение кровеносных капилляров.
Тонкая кишка, преимущественно дистальный отдел (N-клетки)
Уменьшение секреции соляной кислоты желудком, усиление секреции поджелудочной железы.
Тонкая кишка (ЕС1-клетки)
Усиление моторики кишечника, слюноотделения, торможение высвобождения инсулина и всасывания натрия
Проксимальный отдел тонкой кишки (ЕС1- клетки)
Стимуляция сокращений ворсинок тонкой кишки.
Тонкая кишка, немного в поджелудочной железе (G-клетки)
Торможение секреции ферментов поджелудочной железой.
Мобилизация углеводов. Торможение секреции желудка и поджелудочной железы, моторики желудка и кишечника. Пролиферация слизистой оболочки тонкой кишки (индукция гликогенолиза, липолиза, глюконеогенеза и кетогенеза.
Желудочно-кишечный тракт (ЕС1, ЕС2-клетки)
Торможение выделения соляной кислоты в желудке, стимуляция выделения пепсина. Стимуляция секреции поджелудочной железы, кишечной секреции желчевыделения.
Вазоактивный интестиналъный пептид (ВИП)
Желудочно-кишечный тракт (Д1-клетки)
Расслабление гладких мышц кровеносных сосудов, желчного пузыря, сфинктеров. Торможение секреции желудка, усиление секреции бикарбонатов поджелудочной железой и кишечной секреции. Торможение действия ХЦК-ПЗ.
5. Общие принципы регуляции пищеварения
Причем пищевой центр имеет отношение не только к формированию пищевого поведения (обеспечивается в основном латеральными ядрами гипоталамуса – центром голода), но и к регуляции деятельности органов пищеварительной системы (обеспечивается гипоталамусом, лимбической системой и ретикулярной формацией ствола головного мозга). Ведущую роль в формировании чувства голода и пищедобывательного поведения играют латеральные ядра гипоталамуса, возбуждение которых и обуславливает многие субъективные проявления голода (головокружение, чувство "жжения или сосания под ложечкой", головную боль, общую слабость) и побуждает к поиску и приему пищи. Вопрос относительно природы факторов, вызывающих возбуждение нейронов этих ядер и соответственно возникновение чувства голода, является окончательно нерешенным. Очевидно, возбуждение латеральных ядер гипоталамуса может возникать
как в результате местного их раздражения измененным химическим составом крови или церебральной жидкости, омывающих гипоталамус,
так и вследствие поступления афферентной информации от различных периферических хеморецепторов сосудистого русла и механорецепторов самого пищеварительного тракта.
Так, в опытах на животных показано, что сильное возбуждение латеральных ядер гипоталамуса и соответственно чувство голода возникает
при значительном уменьшении уровня глюкозы в периферической крови (отсюда возникла глюкостатическая теория голода)
или при уменьшении содержания аминокислот в крови (аминоацидостатическая теория голода)
или в случае уменьшения концентрации продуктов гидролиза жиров в крови (липостатическая теория голода).
при поступлении к латеральным ядрам гипоталамуса афферентной информации от рецепторов жировых депо, возбуждающихся в момент мобилизации жира из них (липонервостатическая теория)
при увеличении содержания в крови промежуточных метаболитов цикла Кребса, образующихся в результате расщепления различных питательных веществ (метаболическая теория)
наконец, известна зависимость между возникновением у человека чувства голода и водными ресурсами организма: при снижении запасов воды в организме уменьшается и потребление пищи (гидростатическая теория голода).
Предполагается участие в формировании пищевого поведения и определенных гормонов, продуцируемых эндокринными клетками желудочно-кишечного тракта. Так, например, некоторые гормоны кишечника – арентерин, холецистокинин-панкреозимин – угнетают аппетит. Немаловажное значение в формировании чувства голода и насыщения имеет и та информация, которая поступает от рецепторов самого желудочно-кишечного тракта. В частности, наполнение желудка и кишечника пищевым содержимым оказывает тормозное влияние на активность латеральных ядер гипоталамуса и возбуждает вентромедиальные ядра, в результате чего у человека еще до всасывания продуктов гидролиза пищевых веществ в кровь возникает ощущение насыщения (это первичное или сенсорное насыщение). На смену ему приходит вторичное (истинное или обменное насыщение), возникающее в результате поступления в кровь продуктов гидролиза белков, жиров и углеводов, и как следствие этого, изменения ее химизма и уменьшения степени активности хеморецепторов сосудистого русла, реагирующих на понижение содержания каких-то пищевых компонентов в крови.
Регуляция моторной и секреторной функции желудочно-кишечного тракта осуществляется нервным и гуморальным путем. В основе нервных механизмов регуляции лежат условно- и безусловно-рефлекторные реакции, реализующиеся через посредство симпатического и парасимпатического отделов вегетативной нервной системы. Так, обстановка приема пищи, ее вид, запах, звук тарелок и вилок условнорефлекторным путем приводят к повышению активности пищевого центра гипоталамуса и соответственно к усилению секреторной функции слюнных, желудочных и отчасти поджелудочной желез. Благодаря условным пищевым рефлексам пищеварительный тракт еще до поступления пищи в ротовую полость оказывается заранее подготовленным к перевариванию пищи (опережающее значение условных рефлексов). Поступление пищи в ротовую полость и связанное с этим раздражение вкусовых, механо- и терморецепторов сопровождается безусловнорефлекторным усилением секреции не только слюнных, но и других пищеварительных желез, и некоторым повышением моторики ниже расположенных отделов пищеварительного тракта, что подготавливает их к переработке новых порций пищи. Рефлекторные дуги безусловных рефлексов, регулирующих секреторную и моторную деятельность пищеварительной системы, могут замыкаться на уровне
центральной нервной системы (т.н. "длинные" рефлекторные дуги), как правило, обеспечивают координированную перестройку деятельности многих отделов пищеварительного тракта, оказывают пусковое влияние на секреторную активность пищеварительных желез
или периферических интрамуральных парасимпатических ганглиев (т.н. "короткие" рефлекторные дуги), оказывающие корригирующие влияние и обеспечивающие преимущественно местную регуляцию секреторной или моторной функции какого-то отдела пищеварительного тракта с учетом не только той информации, которая поступают из ц.н.с, но и особенностей функционирования органа (объема поступившего пищевого содержимого, его кислотности и некоторых других). Именно периферические рефлексы делают возможным четкое соответствие между качеством и количеством принятой пищи и продолжительностью, объемом секреции, а также ферментативной активностью вырабатываемых пищеварительных соков.
Гуморальные механизмы регуляции деятельности желудочно-кишечного тракта обеспечиваются преимущественно гастроинтестинальными гормонами, продуцируемыми эндокринными клетками его слизистой оболочки. Гастроинтестинальные гормоны по своей химической природе являются пептидами и способны оказывать
как паракринное действие (вблизи места своей секреции),
так и поступать в общий кровоток и воздействовать на секреторную и моторную функции каких-то отделов пищеварительной трубки на расстоянии от места образования (телекринное действие).
Наряду с регуляцией пищеварения гастроинтестинальные гормоны оказывают воздействие и на другие функции и системы организма (обмен веществ всего организма, состояние сердечно-сосудистой системы, деятельность других эндокринных желез). В связи с тем, что некоторые гастроинтестинальные гормоны встречаются в качестве медиаторов в центральной нервной системе (энкефалин, вещество Р, нейротензин и некоторые другие) существует предположение об общности нейронов и эндокринных клеток желудочно-кишечного тракта и возможном происхождении этих эндокринных клеток путем выселения их предшественников в период эмбриогенеза из нервной трубки.
В регуляции деятельности пищеварительного тракта, особенно дистальных его отделов, определенную роль играют и местные механизмы регуляции. Так, механическое и химическое раздражение рецепторов какого-то отдела кишечника вызывает непосредственное повышение секреторной и моторной активности этого же отдела как за счет периферических ("коротких") рефлексов, так и в результате усиления секреции каких-то гастроинтестинальных гормонов.
Роль нервных и гуморальных механизмов регуляции пищеварения неоднозначна для различных отделов пищеварительного тракта. В частности, в проксимальных отделах пищеварительной трубки основное значение в регуляции имеют нервные механизмы, тогда как в дистальных отделах – гуморальные и особенно местные нейрогуморальные механизмы. Так, вид, запах пищи, сама обстановка ее приема, а затем и возбуждение рецепторов ротовой полости поступившей пищей рефлекторным путем усиливают секрецию слюны и желудочного сока; рефлекторные же влияния на секрецию панкреатического, кишечного соков и желчи при этом выражены гораздо слабее, поскольку основную роль в регуляции их секреции имеют гуморальные механизмы (гастроинтестинальные гормоны) и местные периферические рефлексы. Таким образом, в регуляции пищеварительного тракта существует некий градиент распределения нервных и гуморальных влияний: в начальных его отделах велика роль рефлекторных механизмов, тогда как в дистальных – гуморальных и местных нейрогуморальных механизмов регуляции.
Секреция пищеварительных соков железами зависит от многих факторов, среди которых определенную роль играют как первоначальные пусковые механизмы преимущественно рефлекторного характера, так и последующие корригирующие, в основе которых лежат местные нервные и гуморальные влияния. По механизму изменения секреции пищеварительных желез в процессе пищеварения И.П. Павлов выделил следующие основные фазы секреции:
1. сложнорефлекторную или мозговую фазу, в основе которой лежат условные и безусловные рефлексы, возникающие еще до поступления пищи в тот отдел пищеварительной трубки, куда выделяет свой секрет железа. Так, например, возбуждение обонятельных, зрительных и слуховых рецепторов запахом, видом пищи, обстановкой ее приема условнорефлекторным путем стимулирует секрецию желудочного сока. Поступление пищи в ротовую полость и связанное с этим возбуждение вкусовых, тактильных и температурных рецепторов ротовой полости продолжает, но уже безуловнорефлекторным путем стимулировать желудочную и отчасти поджелудочную секрецию (хотя пища еще в желудок и кишечник не поступила)
2. нервно-гуморальная фаза, проявляющаяся в коррекции начатой еще в мозговую фазу секреции пищеварительного сока железой после поступления пищевого содержимого в тот отдел пищеварительного тракта, куда выделяет свой секрет железа. Ведущую роль в осуществлении этой фазы играют местные периферические рефлексы, возникающие в ответ на раздражение собственных рецепторов данного отдела пищеварительного тракта, в который поступает сок пищеварительной железы, а также действие гастроинтестинальных гормонов. Причем в составе нейрогуморальной фазы секреции выделяют две фазы:
желудочную, когда пищевое содержимое уже поступило в желудок, и возбуждение рецепторов желудка рефлекторным или гуморальным (через определенные гастроинтестинальные гормоны) путем корригирует секреторную активность самого желудка и стимулирует секрецию кишечных желез, поджелудочной железы и отчасти печени.
кишечную, в момент которой часть пищевой массы перешла в кишечник. Возбуждение рецепторов кишечника поступившим пищевым содержимым, а также рецепторов кровеносных сосудов продуктами гидролиза пищевых веществ, всасывающихся в кровь, рефлекторным (преимущественно по типу периферических рефлексов) и в основном гуморальным (через изменение секреции определенных гастроинтестинальных гормонов) путям корригирует как секрецию соков, поступающих в просвет самого тонкого кишечника, так и секреторную и моторную активность желудка, обеспечивая достижение соответствия между качеством пищи и количеством и ферментативным составом желудочного, панкреатического и кишечного соков, а также как можно лучшее переваривание пищи в каждом из этих отделов пищеварительной трубки.
Таким образом, влияния на секреторную активность пищеварительных желез в первую мозговую фазу секреции носят преимущественно пусковой характер и осуществляются рефлекторным путем, тогда как в последующие желудочную и кишечную фазы – играют корригирующую роль и обеспечиваются местными периферическими рефлексами и гастроинтестинальными гормонами.
В зависимости от своего характера, нейрогуморальные влияния на органы пищеварительного тракта, могут быть:
функциональными; вегетативная нервная система или какие-то гуморальные факторы оказывают непосредственное стимулирующее или тормозное влияние на определенные органы. Чаще всего такой тип влияний реализуется на отделы собственно пищеварительной трубки (а не на железы) путем воздействия на гладкомышечные волокна их стенки (т.е. такой тип влияний нервная система оказывает преимущественно на моторику желудочно-кишечного тракта). Так, парасимпатический отдел вегетативной нервной системы, как правило, стимулирует моторику желудочно-кишечного тракта, а симпатический – ее тормозит. Причем действие парасимпатического отдела на моторную функцию пищеварительной трубки во многом зависит не только от влияний, поступающих из ц.н.с., но и от состояния самого регулируемого органа. Отмеченная зависимость обусловлена тем, что вторые эфферентные парасимпатические нейроны интрамуральных ганглиев пищеварительной трубки являются общим конечным путем для эфферентной информации, поступающей из ц.н.с. по преганглионарным парасимпатическим нервным волокнам, а также для чувствительной информации от рецепторов самого иннервируемого органа. Тормозные влияния симпатического отдела на моторику желудочно-кишечного тракта не всегда могут реализоваться, поскольку постганглионарные симпатические волокна образуют синапсы не на самих гладкомышечных клетках пищеварительной трубки, а на вторых эфферентных парасимпатических нейронах интрамуральных ганглиев. В результате этого симпатический отдел вегетативной нервной системы зачастую лишь ослабляет активирующее влияние парасимпатического отдела на моторную функцию желудочно-кишечного тракта. Гуморальные факторы могут оказывать как стимулирующее, так и тормозное влияние на моторную и секреторную функции пищеварительного тракта. Так, одни гастроинтестенальные гормоны (мотилин, гастрин) стимулируют моторику желудка, а другие – (секретин, холецистокинин-панкреозимин) – ее тормозят. Аналогично, одни гуморальные факторы (гастрин) усиливают желудочную секрецию (в основном стимулируя выделение соляной кислоты), а другие (секретин, холецистокинин-панкреозимин) – тормозят секрецию соляной кислоты, но усиливают секцию пищеварительных ферментов в составе желудочного сока.
сосудодвигательными; вегетативная нервная система может влиять на функциональную активность органа косвенно путем изменения уровня его кровоснабжения. Так, возбуждение парасимпатических нервных волокон, иннервирующих какую-то пищеварительную железу, оказывает сосудорасширяющее действие на ее артериолы, что увеличивает уровень кровоснабжения железы, а, следовательно, и ее функциональную активность. Такой тип влияний, как правило, реализуется в регуляции активности пищеварительных желез и в меньшей мере в косвенной регуляции моторики желудочно-кишечного тракта. Изменение уровня кровоснабжения органов пищеварительной системы может происходить как в результате непосредственных влияний из ц.н.с. по эфферентным симпатическим или парасимпатическим нервам на гладкомышечные клетки сосудов, так и возникать по причине накопления в усиленно работающих органах местных продуктов метаболизма, оказывающих местное сосудорасширяющее действие
трофическими; заключаются в том, что вегетативная нервная система и ряд гуморальных факторов могут изменять функциональную активность органов пищеварительной системы путем изменения интенсивности анаболических процессов в них (синтеза белков, нуклеиновых кислот), что будет отражаться на функциональной активности или общем количестве составляющих эти органы клеток. Так, гастрин стимулирует пролиферацию париетальных клеток в желудочных железах, вырабатывающих соляную кислоту, а холецистокинин-панкреозимин – гландулоцитов в ацинусах поджелудочной железы. Эфферентные влияния симпатического отдела вегетативной нервной системы, как правило, способствуют нормальной трофике большинства органов организма, в том числе и пищеварительного тракта.
Между функциональной активностью различных отделов пищеварительного тракта существуют очень сложные взаимоотношения, которые обычно проявляются в следующей закономерности. Механическое или химическое раздражение рецепторов какого-то отдела пищеварительного тракта компонентами поступившего в него пищевого содержимого, как правило, приводит к повышению секреторной и моторной активности этого отдела и дистальнее расположенных отделов, но угнетает моторику проксимальнее расположенных отделов пищеварительной трубки, при этом определенным образом изменяя и их секреторную активность. Так, поступление кислого содержимого из желудка в кишечник, с одной стороны, через усиление секреции секретина эндокринными клетками слизистой кишечника стимулирует образование и выделение из поджелудочной железы сока, богатого бикарбонатами, что способствует нейтрализации поступившего пищевого комка. С другой стороны, сам секретин, а также рефлекторные реакции, возникающие в ответ на возбуждение механо- и хеморецепторов двенадцатиперстной кишки, несколько подавляют секрецию соляной кислоты в составе желудочного сока (путем торможения образования гастрина G-клетками желудка) и тормозят моторику желудка, но при этом стимулируют моторную и секреторную функцию нижерасположенных отделов пищеварительной трубки, тем самым ускоряя перерабатывание находящихся в них пищевых масс и подготавливая к приему новых. Тормозное же влияние на моторику вышерасположенных отделов пищеварительной трубки со стороны нижерасположенных способствует более длительному нахождению там пищи и лучшему ее перевариванию.
Таким образом, пищеварительный конвейер функционирует как единое целое, а находящаяся в нем смесь пищевого содержимого с пищеварительными соками является не только объектом пищеварительных воздействий, но и средством регуляции самого пищеварительного процесса (поскольку консистенция пищевого содержимого, его рН в различных отделах пищеварительного тракта, степень гидролиза питательных веществ и другие факторы через посредство нервных и гуморальных механизмов во многом определяют объем и особенности состава секретируемых пищеварительных соков, а также моторику различных отделов пищеварительного тракта).
66. Принципы регуляции деятельности пищеварительной системы. Роль рефлекторных, гуморальных и местных механизмов регуляции. Гормоны жкт, их классификация.
Регуляция деятельности пищеварительной системы осуществляется тремя механизмами:
Рефлекторный компонент вызывает торможение или активацию моторной деятельности при возбуждении рецепторов. Повышает моторную функцию парасимпатический отдел: для верхний части – блуждающие нервы, для нижней – тазовые. Тормозное влияние осуществляется за счет чревного сплетения симпатической нервной системы. При активации нижележащего отдела желудочно-кишечного тракта происходит торможение выше расположенного отдела. В рефлекторной регуляции выделяют три рефлекса:
гастроэнтеральный (при возбуждении рецепторов желудка активируются другие отделы);
энтеро-энтеральный (оказывают как тормозное, так и возбуждающие действие на нижележащие отделы);
ректо-энтеральный (при наполнении прямой кишки возникает торможение).
Гуморальные механизмы преобладают в основном в двенадцатиперстной кишке и верхней трети тонкого кишечника.
Возбуждающее действие оказывают:
мотилин (вырабатывается клетками желудка и двенадцатиперстной кишки, оказывает активирующее влияние на весь желудочно-кишечный тракт);
гастрин (стимулирует моторику желудка);
бамбезин (вызывает отделение гастрина);
холецистокинин-панкреозинин (обеспечивает общее возбуждение);
секретин (активирует моторику, но тормозит сокращения в желудке).
Тормозное влияние оказывают:
вазоактивный интестинальный полипептид (вызывает расслабление гладких мышц кровеносных сосудов (снижение артериального давления), желчного пузыря, сфинкторов, торможение секреции желудка);
гастроингибирующий полипептид (тормозит секрецию соляно кислоты желудка, пепсина);
соматостатин (вызывает торможение высвобождения желудочно-кишечных гормонов и секреции желез желудка);
энтероглюкагон (тормозит секрецию соляной кислоты желудка, пепсина).
Гормоны желез внутренней секреции также влияют на моторную функцию. Так, например, инсулин ее стимулирует, а адреналин тормозит.
Местные механизмы осуществляются за счет наличия метсимпатической нервной системы и преобладают в тонком и толстом кишечнике. Стимулирующее действие оказывают:
грубые непереваренные продукты (клетчатка);
конечные продукты расщепления белков и углеводов.
Тормозное действие возникает при наличии липидов.
67. Пищеварение в полости рта: состав и физиологическая роль слюны. Слюноотделение и его регуляция.
Пищеварение в полости рта продолжается 15-20 секунд. Там она подвергается физической обработке, измельчению, растиранию, смачиванию слюной, частично – химической обработке и органолептическому анализу, т.е. определению вкуса пищи. Это обуславливает положительную или отрицательную реакцию на пищу. В ротовой полости пища раздражает вкусовые, тактильные, болевые, температурные рецепторы. Вкусовые рецепторы преимущественно располагаются на сосочках языка (сладкое – кончик языка, соленое и кислое – боковая поверхность языка, горькое – корень языка). Тактильные, температурные, болевые рецепторы располагаются по все поверхности слизистой ротовой полости.
Состав и свойства слюны:
Слюна выделяется околоушной, поднижнечелюстной и подъязычной железами, а также мелкими железами, расположенными на поверхности языка, слизистой оболочки щёк и неба. Околоушная железа состоит из серозных клеток, выделяет серозную слюну, которую также выделяют железы, расположенные на боковых поверхностях языка. Поднижнечелюстная и подъязычная железы смешанные, состоят из слизистых и серозных клеток, выделяют смешанную слюну. Смешанные железы располагаются на слизистой оболочке щек, языка, губ. Слизистые железы мелкие, располагаются на слизистой корня языка, глотки, твердого и мягкого неба. Крупные железы выделяют слюну во время подготовки к приему пищи и во время приема. Мелкие железы выделяют слюну постоянно, что имеет важное значение – смачивание ротовой полости, обеспечение разговорной речью. За сутки выделяется 1-1,5 литра слюны.
В слюне имеются органические (альбумины, глобулины, аминокислоты, мочевина, мочевая кислота, лизоцим) и неорганические (соли Nа, Са, К, сернокислые и азотнокислые соли, йодиды, бромиды, хлористые и роданистые соединения) вещества. Реакция слюны – слабощелочная, рН = 7,36. В слюне содержится 98 – 99 % воды и 1 – 2 % плотного остатка. Слюна содержит ферменты – амилаза (птеонин), мальтаза (действуют на крахмал, расщепляя его до амилодекстринов, а затем до моносахаридов). Также в слюне содержатся: протеиназа, липаза, фосфатаза, лизоцим (бактерицидное действие), калликреин. Железы также секретируют паратин, который контролирует синтез белков, уровень Са в крови, усиливает сперматогенез, гематопоэз, повышает проницаемость гистогематических барьеров. Слюнные железы секретируют фактор роста нервов, эпидермальный фактор, фактор роста эпителия сосудов, кожи, мышц, почек. Т.о. слюнные железы выполняют экзо - и эндокринные функции.
Физиологическим раздражителем слюнных желез является пища. В зависимости от органического состава пища может изменять количественный и качественный состав слюны. Опытами установлено, что на сухари выделяется 4 мл слюны, на мясные сухари – 6 мл, на хлеб – 3,2 мл, на мясо – 2 мл, на молоко – 1-2 мл. Этот количественный состав слюны объясняется тем, как эти вещества воздействуют на рецепторы в слизистой ротовой полости. Сухая пища раздражает сильнее, чем жидкая. Слюна выделяется на отвергаемые физические и химические вещества. Опыты показали, что введение песка в ротовую полость собаки вызывает обильное слюноотделение. Введение HCl также стимулирует слюнообразование и слюноотделение.
При выделении слюны выделяют 3 фазы (периода):
латентный период – это время от начала дачи пищи до начала выделения слюны. Он зависит от состава пищи. При сильном раздражении он равен 2-3 секундам, при слабом – 20-30 секунд.
период слюноотделения – время, в течение которого пища находится в ротовой полости.
период последействия – пища промочена, а слюна еще выделяется.
Механизм регуляции слюноотделения:
Попадая в рот, пища подвергается механической и химической обработке и своими физико-химическими свойствами раздражает рецепторы ротовой полости. В рецепторах генерируются потенциалы действия, которые в виде нервных импульсов, по афферентным нервам (n. lingualis, n. glossopharyngeus и верхний гортанный нерв), поступает в продолговатый мозг, в центр слюноотделения. Этот центр возбуждается и посылает импульсы по парасимпатическим и симпатическим нервам к слюнным железам. Парасимпатическая иннервация осуществляется n. glossopharyngeus и n. facialis. Околоушная железа иннервируется n. glossopharyngeus, от него к железе подходит веточка auriculartemporalis. Поднижнечелюстная и подъязычная железы иннервируются n. facialis, от него к железе подходит веточка chordatympani. Центр симпатической иннервации располагается в боковых рогах II- VI грудных сегментов. От них начинаются преганглионарные волокна, которые идут к верхнему шейному симпатическому ганглию, прерываются, и от этого ганглия начинаются постганглионарные нервные волокна, иннервирующие эти слюнные железы. Раздражение (возбуждение) парасимпатических нервов вызывает обильную секрецию слюны, бедной органическими веществами. Раздражение симпатических нервов вызывает секрецию небольшого количества слюны, но она богата органическими веществами.
Кроме этого, в регуляции образования и выделения слюны принимают участие и условно-рефлекторные механизмы. Слюна может выделяться на звуки, приготовление пищи, вид пищи, запах и т.д.
Основные функции слюны:
Смачивание, необходимое для:
вкусовой оценки (на вкусовые рецепторы действуют только растворенные вещества);
химической обработки пищи;
проглатывания (сухая пища не проглатывается);
начала образования химуса;
вымывания остатков пищи из ротовой полости;
действия защитных компонентов слюны.
Защитная функция(гигиена полости рта), обусловленная:
бактерицидным действием лизоцима и иммуноглобулинов;
вымыванием остатков пищи и бактерий из ротовой полости;
обволакиванием слизистой полости рта муцином.
Начало химической обработки пищи - начало переваривания углеводов под действием α-амилазы.
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
Регуляция секреции желудочного сока. Принципы секреции желудочного сока.
Железы желудка в состоянии относительного покоя (в условиях отсутствия процесса пищеварения) выделяют небольшое количество сока нейтральной или слабощелочной реакции (фоновая секреция). Под влиянием пищевого раздражения во время приема пищи железы желудка секретируют значительный объем желудочного сока, богатого протеолитическими ферментами. Эта реакция желез является рефлекторным ответом на раздражение пищей рецепторов слизистой оболочки ротовой полости, глотки и желудка (безусловный рефлекс) и воздействие комплекса раздражителей, влияющих на другие рецепторы, предшествующих и сопутствующих приему пищи (условный рефлекс).
Афферентные импульсы с раздражаемых пищей рецепторов (тактильных, температурных и вкусовых) по афферентным волокнам V, VII, IX и X черепно-мозговых нервов передаются в бульбарный, таламический, гипо-таламический и корковый отделы пищевого центра. Нисходящие влияния коркового представительства пищевого центра активируют парасимпатические и симпатические ядра гипоталамуса. При возбуждении парасимпатических ядер их эфферентные влияния активируют клетки бульбарного отдела пищевого центра, импульсы от которых по преганглионарным волокнам блуждающего нерва передаются на постганглионарные нейроны ин-трамуральных ганглиев желудка (клетки Догеля I типа). Выделяемый в окончаниях их аксонов медиатор ацетилхолин через М-холинорецепторы мембраны гландулоцитов стимулирует секреторную деятельность главных, обкладочных и мукоидных клеток желез желудка. Поэтому после перерезки блуждающих нервов у собак не выделяется желудочный сок при раздражении пищей рецепторов полости рта, на ее вид и запах.
Ацетилхолин активирует также эндокринные У-клетки слизистой оболочки антрального отдела желудка, вырабатывающие гастрин, который через кровоток достигает обкладочных клеток желез и через специальные рецепторы их мембраны вызывает выработку соляной кислоты.
Ацетилхолин и гастрин в слизистой оболочке фундального отдела желудка стимулируют активность эндокринных клеток, выделяющих гиста-мин, который, диффундируя по межклеточным пространствам, через Н2-рецепторы мембраны обкладочных клеток стимулирует выработку большого количества кислого желудочного сока, бедного ферментами и мукоида-ми. После перерезки блуждающих нервов (в экспериментах на животных) сокогонный эффект ацетилхолина, гастрина и гистамина резко ослабляется.
Поступление пищи в желудок во время еды усиливает безусловно-рефлекторное отделение желудочного сока, что обусловлено раздражением механорецепторов при растяжении его стенок (особенно антрального отдела) и хеморецепторов пептидами и экстрактивными веществами пищи. При этом рефлекс осуществляется с участием бульбарного отдела пищевого центра, дополняется местной реакцией, реализуемой через холинергиче-ские нейроны интраорганной нервной системы.
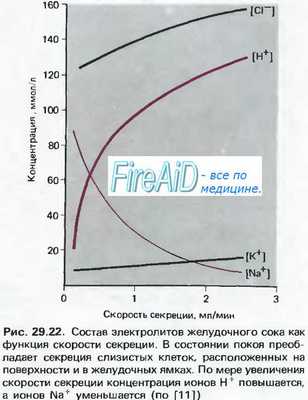
Увеличение концентрации соляной кислоты в желудочном соке приводит к угнетению выработки гастрина У-клетками и последующим уменьшением кислотности сока. Этот механизм саморегуляции предотвращает избыточное выделение соляной кислоты. Подавляют секрецию соляной кислоты соматостатин, ВИП и серотонин (см. табл. 11.1).
При возбуждении симпатических ядер гипоталамуса его нисходящие влияния стимулируют преганглионарные симпатические нейроны боковых рогов торакальных сегментов (IV—X) спинного мозга. Их эфферентные влияния стимулируют постганглионарные нейроны симпатических ганглиев, аксоны которых в составе чревных нервов достигают желез желудка. Выделяющийся в их окончаниях медиатор норадреналин тормозит секрецию желудочного сока, но приводит к накоплению секреторного материала в гландулоцитах, содержащих пепсиногены и мукоиды.
Поступление в двенадцатиперстную кишку не полностью переваренного химуса из желудка вызывает дополнительное возбуждающее и тормозное влияние на железы желудка. Гастрин, вырабатываемый У-клетками слизистой оболочки кишки, бомбезин (гастрин-рилизинг-гормон), выделяемый Р-клетками, продукты гидролиза белков гуморально стимулируют выработку соляной кислоты.
Выделяемые эндокринными клетками двенадцатиперстной кишки холецистокинин, гастроингибирующий пептид, нейротензин и другие пептиды (см. табл. 11.1) под влиянием продуктов гидролиза жиров в двенадцатиперстной кишке оказывают тормозной эффект на выработку соляной кислоты железами желудка.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: