Орган Корти. Аппарат слуха.
Добавил пользователь Дмитрий К. Обновлено: 15.01.2026
периферическая часть звуковоспринимающего аппарата (рецептор слухового анализатора (См. Слуховой анализатор)) у млекопитающих животных и человека. Открыт итальянским гистологом А. Корти (A. Corti; 1822—76). В процессе эволюции возникает на основе структур боковых органов (См. Боковые органы). К. о. располагается в спирально завитом костном канале внутреннего уха (См. Внутреннее ухо) — улитковом ходе, заполненном эндолимфой. Верхняя стенка хода прилегает к т. н. лестнице преддверия и называется рейснеровой перепонкой; нижняя стенка, граничащая с т. н. барабанной лестницей, образована основной перепонкой, прикрепляющейся к спиральной костной пластинке. Лестницы преддверия и барабанная заполнены перилимфой. Наружная стенка улиткового хода, или сосудистая полоска, содержит много кровеносных сосудов. К. о. расположен на основной перепонке и состоит из внутренних и наружных волосковых клеток, внутренних и наружных опорных клеток (столбовых, клеток Дейтерса, Клаудиуса, Гензена), между которыми находится туннель, где проходят направляющиеся к основаниям волосковых клеток отростки нервных клеток, лежащих в спиральном нервном ганглии. Воспринимающие звук волосковые клетки располагаются в нишах, образуемых телами опорных клеток, и имеют на поверхности, обращенной к покровной перепонке, по 30—60 коротких волосков. Опорные клетки выполняют также трофическую функцию, направляя поток питательных веществ к волосковым клеткам. Функция К. о. — преобразование (трансформация) энергии звуковых колебаний в процесс нервного возбуждения. Звуковые колебания воспринимаются барабанной перепонкой и через систему косточек среднего уха передаются жидким средам внутреннего уха — перилимфе и эндолимфе. Колебания последних приводят к изменению взаиморасположения волосковых клеток и покровной перепонки К. о., что вызывает сгибание волосков и возникновение биоэлектрических потенциалов (См. Биоэлектрические потенциалы), улавливаемых и передаваемых в центральную нервную систему отростками нейронов спирального ганглия, подходящими к основанию каждой волосковой клетки. По др. представлениям, волоски звуковоспринимающих клеток — лишь чувствительные антенны, деполяризующиеся под действием приходящих волн за счёт перераспределения ацетилхолина эндолимфы. Деполяризация вызывает цепь химических превращений в цитоплазме волосковых клеток и возникновение нервного импульса в контактирующих с ними нервных окончаниях. Различающиеся по высоте звуковые колебания воспринимаются различными отделами К. о.: высокие частоты вызывают колебания в нижних отделах улитки, низкие — в верхних, что связано с особенностями гидродинамических явлений в ходе улитки.
Лит.: Винников Я. А., Титова Л. К., Кортиев орган. Гистофизиология и гистохимия, М.— Л., 1961; Wandsronk С., On the mechanism of hearing, [Leiden], 1961; Vinnikov Ya. A., The ultrastructural and cytochemical bases of the mechanism of function of the sense organ receptors, в кн.: The structure and function of nervous tissue, v. 2, N. Y.— L., 1969, p. 265—392.
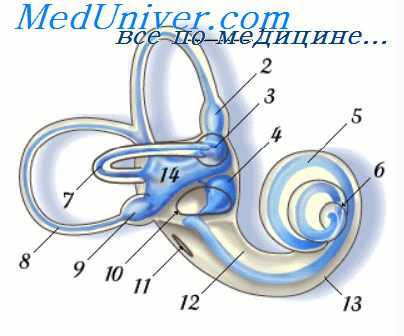
Разрез улитки и кортиева органа (схема): 1 — лестница преддверия; 2 — барабанная лестница; 3 — улитковый ход; 4 — рейснерова перепонка; 5 — сосудистая полоска; 6 — спиральная пластинка; 7 — основная перепонка; 8 — внутренние и 9 — наружные волосковые клетки; 10 — внутренние и 11 — наружные опорные клетки; 12 — клетки Дейтерса; 13 — клетки Клаудиуса; 14 — клетки Гензена; 15 — туннель; 16 — спиральный ганглий; 17 — покровная перепонка.
Большая советская энциклопедия. — М.: Советская энциклопедия . 1969—1978 .
Орган Корти. Аппарат слуха.
Аппарат слуха (орган Корти) располагается на баэилярной мембране. У внутренней стороны Кортиева органа спиральная пластинка утолщается, образуя лимб, который вдается в просвет улиткового протока.
От верхней губы лимба протянута гелеобразная покровная мембрана (текториальная мембрана), которая своим нижним основанием лежит на волосковых клетках органа Корти. Покровная мембрана представляет собой бесклеточный матрикс, состоящий из фибрилл и гомогенной субстанции. От нижней губы лимба протянута базилярная мембрана.
Орган Корти состоит из одного ряда внутренних волосковых клеток и трех рядов наружных волосковых. опорных и столбовых клеток. Между наружными и внутренними клетками расположены опорные клетки Дейтерса, кнаружн от них — высокие опорные клетки Гензена. а далее, кнаружи, - цилиндрические клетки Клаудиуса.
Столбовые клетки формируют туннель органа Корти. Сам перепончатый проток улитки заполнен эндолимфой. Волосковые клетки омываются кортилимфой, которая, вероятно, образуется капиллярами, расположенными под базилярной мембраной.
В поддержании гомеостаза в органе Корти участвуют апудоциты, которые продуцируют биогенные амины, адреналин и норадреналин. Сами апудоциты обнаружены в сосудистой полоске.
Базилярная мембрана в основном состоит из радиальных волокон диаметром до 18—25 мм, которые формируют компактный слой. Структура базилярной мембраны отличается по мере ее удаления от основания к верхушке улитки. У основания улитки волокна мембраны более плотные, чем на верхушке. Кроме того, базилярная мембрана расширяется по направлению к верхушке улитки. Если у основания улитки ширина мембраны до 0,16 мм, то у геликотремы ее ширина достигает 0,52 мм.
Рецепторная клетка органа слуха устроена по общему принципу с чувствительной волосковой клеткой аппарата равновесия. Однако форма наружных и внутренних волосковых клеток отличается. Верхняя поверхность каждой внутренней волосковой клетки покрыта кутикулярной мембраной. в которую погружены стереоцилии. Каждая из этих клеток имеет до 40 волосков, выстроенных в два и более ряда. Свободными от кутикулы остаются участки, удаленные от модиолюса (у наружного края внутренней волосковой клетки). В этих участках располагается базальное тело или измененная киноцилия.
Верхняя поверхность наружных волосковых клеток содержит до 150 стереоцилии, расположенных в три ряда. Верхушки самых длинных волосков контактируют с покровной мембраной.
Слуховой анализатор включает элементы звукопроведеиия - это наружный слуховой проход, среднее ухо и перилимфа лабиринта, а также элементы звуковосприятия, состоящие из периферического рецептора и центрального анализатора. Периферический рецептор представлен перепончатым улитковым протоком, эндолимфой, органом Корти и спиральным ганглием. Центральный слуховой анализатор образован слуховыми ядрами, проводящими путями и височной долей коры головного мозга.
- Вернуться в оглавление раздела "Анатомия человека."
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Орган Корти. Физиология и функция органа Корти
а) Функция органа Корти. Орган Корти, показанный на рисунках ниже, является рецепторным органом, который генерирует нервные импульсы в ответ на вибрацию основной мембраны.
Улитка Разрез через один из витков улитки Орган Корти; особо выделены волосковые клетки и покровная мембрана, под давлением которой находятся погруженные в нее волоски
Обратите внимание, что орган Корти лежит на поверхности базальных волокон и основной мембраны. Истинными сенсорными рецепторами в органе Корти являются два специализированных типа нервных клеток, называемых волосковыми клетками: один ряд внутренних волосковых клеток, число которых равно примерно 3500, а диаметр каждой — около 12 мкм, и три или четыре ряда внешних (или наружных) волосковых клеток, числом около 12000 и диаметром каждой около 8 мкм.
Базолатеральные поверхности волосковых клеток формируют синапсы с сетью окончаний нерва улитки. От 90 до 95% этих синапсов локализуются на внутренних волосковых клетках, указывая на их особое значение для восприятия звука.
Нервные волокна, стимулируемые волосковыми клетками, ведут к спиральному ганглию Корти, который лежит в костном стержне (центре) улитки. Нервные клетки спирального ганглия посылают примерно 30000 аксонов в нерв улитки и затем в центральную нервную систему на уровне верхнего отдела продолговатого мозга. Связь органа Корти со спиральным ганглием и улитковым нервом показана на первом рисунке выше.
а) Возбуждение волосковых клеток. На последнем рисунке выше видно, что от волосковых клеток вверх выступают тончайшие волоски, или стереоцилииу которые касаются или погружены в поверхностное желеобразное покрытие покровной мембраны, лежащей над стереоцилиями в средней лестнице. Эти волосковые клетки подобны волосковым клеткам в макулах и кристах ампул вестибулярного аппарата (просим вас пользоваться формой поиска выше). Сгибание волосков в одном направлении деполяризует волосковые клетки, а сгибание в противоположном направлении гиперполяризует их.
Это, в свою очередь, возбуждает волокна слухового нерва, синаптически связанные с волосковыми клетками.
На рисунке ниже показан механизм, с помощью которого вибрация базальной мембраны возбуждает окончания волосков.
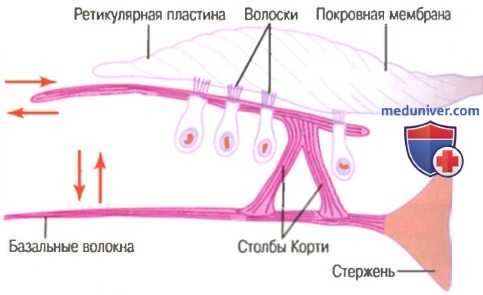
Стимуляция волосковых клеток при движениях вперед-назад их волосков, погруженных в гель, покрывающий покровную мембрану
Наружные концы волосковых клеток плотно фиксированы в ригидной структуре, состоящей из плоской пластины, или ретикулярной мембраны, которая поддерживается треугольными столбами Корти, плотно прикрепленными к базальным волокнам. Базальные волокна, столбы Корти и ретикулярная мембрана двигаются, как единая ригидная структура.
При движении основной мембраны вверх ретикулярная мембрана сдвигается вверх и внутрь по направлению к центральному костному стержню улитки. Затем, когда основная мембрана отклоняется вниз, ретикулярная мембрана колеблется вниз и наружу. Движение внутрь и наружу ведет к тому, что волоски волосковых клеток сдвигаются вперед-назад относительно покровной мембраны. Таким образом, волосковые клетки возбуждаются каждый раз, когда колеблется основная мембрана.
б) Слуховые сигналы передаются в основном внутренними волосковыми клетками. Хотя наружных волосковых клеток в 3-4 раза больше, чем внутренних, примерно 90% волокон слухового нерва стимулируются внутренними, а не наружными клетками. Но несмотря на это, если наружные клетки поражаются, а внутренние функционально остаются совершенно нормальными, происходит значительная потеря слуха. Следовательно, как полагают, наружные волосковые клетки каким-то образом регулируют чувствительность внутренних волосковых клеток к звукам разной высоты — феномен, называемый «настройкой» рецепторной системы.
Эта концепция поддерживается тем фактом, что большое количество ретроградных нервных волокон идет из ствола мозга к месту расположения наружных волосковых клеток. Стимуляция этих нервных волокон может вызывать укорочение наружных волосковых клеток и, возможно, изменяет также степень их жесткости. Эти эффекты позволяют предполагать наличие ретроградного нервного механизма регуляции чувствительности уха к различной высоте звука, активируемого через наружные волосковые клетки.
Видео физиология слухового анализатора - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Физиология кортиева органа внутреннего уха
Улитка является органом слуха млекопитающих, заключена в височную кость и представляет собой костную структуру по форме напоминающую раковину улитки («cochlos» — греч. «улитка»). У людей улитка закручена в 22/3 оборота вокруг центральной оси, модиолуса. В костной части улитки (также известной как костный лабиринт), имеются каналы или лестницы, образованные перепончатым лабиринтом: центральная, срединная лестница, также известная как улитковый проток, отделена от вестибулярной лестницы преддверной мембраной (мембраной Рейсснера) и от барабанной лестницы базальная мембраной.
Вестибулярная лестница соединяется со средним ухом через овальное окно, которое прилегает к подножной пластинке стремени. Круглое окно, соединяющее барабанную лестницу и среднее ухо, прикрыто мембраной круглого окна. Вестибулярная и барабанная лестница соединяются на вершине улитке (в области геликотремы); улитковый проток заканчивается слепо. Вестибулярная и барабанная лестницы наполнены перилимфой, внеклеточной жидкостью с высоким содержанием Na + и низким содержанием К + , в то время как улитковый проток наполнен эндолимфой, отличающейся высоким содержанием К + и низким содержанием Na + . Эндолимфа улитки имеет положительный электрический потенциал, который приблизительно равен +85 мВ.
Разница в концентрации ионов между перелимфой и эндолимфой и разность потенциалов обеспечивает энергию, необходимую для работы улитки.
С физиологической точки зрения в улитке можно выделить три функциональных отдела:
(1) кортиев орган, представляющий «датчик» улитки, преобразующий механические колебания в электрические сигналы (механоэлектрическая передача);
(2) сосудистую полоску, улитковую «батарею», генерирующую энергию (эндокохлеарный потенциал), для механоэлектрической передачи и влияющую на гомеостаз жидкостей улитки;
(3) спиральный ганглий, включающий аксоны нейронов («электрический провод»), передающий электрические сигналы от улитки к центральной нервной системе.
Все три отдела необходимы для нормального функционирования улитки и будут детально описаны в отдельной статье на сайте (список представлен в конце статьи, либо просим пользоваться формой поиска на главной странице сайта).
Кортиев орган является рецептором органа слуха млекопитающих. Так он был назван после того, как в XIX веке, итальянский гистолог Альфонсо Корти, впервые обнаружил и описал этот морфологически сложный орган.
Кортиев орган состоит из двух типов чувствительных рецепторов, внутренних и наружных волосковых клеток. Около 3500 бокаловидных внутренних волосковых клеток выстроились в один ряд по всей длине улиткового протока. Латеральнее внутренних клеток обнаруживаются три ряда наружных волосковых клеток, отличающихся специфической цилиндрической формой.
Оба типа волосковых клеток представляют собой пучки из высокоорганизованных, содержащих актин стереоцилий с увеличением высоты к латеральному ряду, таким образом, самые короткие находятся в середине. Волосковые пучки волосковых клеток внутреннего уха образуют изогнутую линию, состоящую из 2-3 рядов стереоцилий. Пучки наружных волосковых клеток имеют вид буквы V. Пучки волосковых клеток являются их механочувствительными органеллами.
Каждая волосковая клетка располагается на верхушке фаланговой клетки, которая у наружных волосковых клеток называется клеткой Дейтерса. Внутренние и наружные столбовые (поддерживающие) клетки условно разделяют пространство между внутренними и наружными волосковыми клетками и образуют туннель кортиева органа. Также имеются и другие поддерживающие клетки кортиева органа. С медиальной стороны расположены внутренние пограничные клетки, латеральнее — клетки Гензена (кнаружи от пограничных клеток), клетки Клаудиуса, и клетки Беттхера. По всей своей длине кортиев орган покрыт покровной мембраной. Эта бесклеточная структура, внутренней частью укрепленная на преддверной губе спиральной пластинки улитки и соединенная с пучками наружных волосковых клеток.
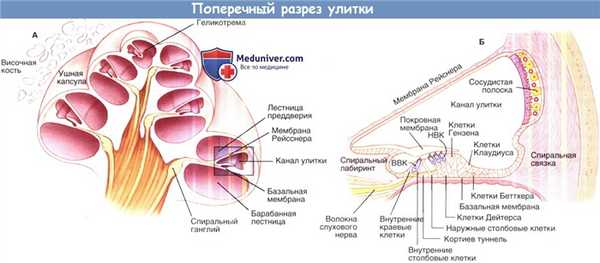
Поперечный разрез улитки. А, схематичное изображение улитки в разрезе.
Выделенная область, показанная на рисунке Б, изображает улитковый проток и окружающие структуры, такие как кортиев орган и сосудистую полоску в разрезе.
ВВК — внутренние волосковые клетки; НВК — наружные волосковые клетки.
а) Базиллярная мембрана и тонотопия кортиева органа. Когда звук воздействует на барабанную перепонку, вибрация передается на внутреннее ухо через три слуховые косточки. Движение стремени вызывает смещение жидкости улитки в вестибулярной лестнице. Несжимаемость перелимфы создает градиент давления между вестибулярной и барабанной лестницами, приводя в движение базальную мембрану и кортиев орган. Это смещение может быть представлено в виде бегущей волны, которая движется от основания к верхушке вдоль базальной мембраны. Для стимулов чистых тонов бегущая волна достигает максимума в определенном месте базальной мембраны и затем спадает.
Точное месторасположение этого максимума зависит от частоты стимула, что является основополагающим принципом тонотопической организации в улитке. Специфическое влияние частоты на определенные участки базальной мембраны регулируется свойствами всех пассивных компонентов, таких как внеклеточные, клеточные и молекулярные структуры в этом месте, а также свойствами активных систем, таких как кохлеарный усилитель (описан далее). Основание улитки у людей настроено на частоты до 20 кГц, а в области верхушки на частоты до 20 Гц. Тонотопический градиент анатомически проявляется не только в изменении ширины базальной мембраны, но и в изменениях высоты волосковых клеток и длинны клеточных структур, таких как стереоцилии и собственно пучки волосковых клеток.
Внутренние волосковые клетки улитки служат основой слуха, являясь сенсорными клетками, преобразующими механические стимулы в электрические сигналы и посредством синаптической активности передающими эти сигналы в мозг. В основе этого процесса лежит механоэлектрическая передача, которая возникает на самом конце или около кончика стереоцилии. Упомянутый аппарат механоэлектрической передачи присутствует во всех волосковых клетках и состоит из одного или более механически закрытых катионных каналов, тесно связанных эластичных структур и концевой связью стереоцилии со следующей, более высокой стереоцилией.
Механические отклонения стереоцилий волосковых клеток в сторону более высоких рядов приводит к смещению соседних стереоцилий. Постепенное увеличение механического напряжения в передаточном аппарате ведет к конформационным изменениям белка-передатчика в канале и к увеличению пропускной способности канала, которая в покое у млекопитающих составляет 40-50%. Несмотря на некоторое количество белков-кандидатов, ни один из предполагаемых компонентов передаточного аппарата волосковых клеток не был однозначно функционально связан с биофизическим процессом механической передачи. Наиболее вероятным кандидатом является кадхерин-23 и протокадхерин-15, которые были предложены в качестве компонентов концевой связи, а также миозин-1с, необходимый для адаптационного процесса, контролирующего заданное значение механочувствительности.
Мутация гена, кодирующего либо кадхерин-23, либо протокадхерин-15 у человека, приводит к синдрому Ушера (врожденной глухоте с прогрессирующей потерей зрения в результате пигментного ретинита).
При механической стимуляции в направлении самого высокого ряда стереоцилий ионы К + и Са 2+ входят в волосковые клетки через открытые каналы механоэлектрической передачи, находящиеся около верхушек стереоцилий. Это отклонение ведет к деполяризации клетки. При отклонении стереоцилии, канал передачи закрывается, тем самым гиперполяризируя клетку. После длительного сгибания пучков волосковых клеток происходит адаптация изначально большой пропускной способности канала, проявляясь уменьшением тока в связи с закрытием передаточных каналов. Было сделано предположение, что за адаптацию отвечают два различных процесса: быстрое закрытие каналов передачи и скольжение миозина, связанно с передаточным аппаратом. Быстрое повторное включение каналов или «быстрая адаптация» предположительно связана с Са 2+ во внутриклеточном пространстве около ворот канала. Точный механизм этого процесса пока неясен.
Второй процесс, «медленная адаптация», происходит примерно в 10 раз медленнее, чем быстрое открытие канала, и возникает при соскальзывании концевого соединения стреоцилии вниз. Во время стимула адаптация ведет к переустановке точки покоя, тем самым позволяя передаточному аппарату непрерывно функционировать в области наивысшей чувствительности. Предполагается, что поступление Са 2+ через открытые каналы ведет к проскальзыванию входящего в адаптационный аппарат миозина, который смещается по направлению к вершине стереоцилии вдоль актиновых нитей. Проскальзывание миозина уменьшает напряженность на концевом соединении и снижает пропускную способность передаточного канала, которая в свою очередь, выключает местный приток Са 2+ .
При низких концентрациях Са 2+ миозин адаптационного аппарата будет эффективно двигаться вверх, тем самым восстанавливая напряженность концевой связи до точки, когда пропускная способность передаточного канала будет близка к пропускной способности в покое. Миозин 1с выдвигается в качестве важного компонента адаптационного механизма, что не исключает участия других типов миозина в этом процессе.
Стереоцилии улитки. Показан апикальный участок кортиева органа, длиной 150 мкм.
Пучки стереоцилий окрашены фаллоидином, меченым флюоресцеином, который связывается с филаментами актина.
Изогнутые пучки внутренних волосковых клеток находятся внизу, наверху видны три ряда V-образных пучков наружных волосковых клеток. Тонотопическая организация кортиева органа.
Схематическое изображение анатомических изменений на протяжении улитки от основания к верхушке,
включающее увеличение ширины базальной мембраны и размера наружных волосковых клеток.
Эти изменения обеспечивают перестройку частоты кортиева органа.
Подобным образом чувствительность к ототоксическому повреждению, такому как шум или действие аминогликозидов, выше в основании улитки и уменьшается по направлению к верхушке.
IHC — внутренние волосковые клетки; ОНС — наружные волосковые клетки.
б) Наружные волосковые клетки и амплификация. Наружные волосковые клетки играют ключевую роль в амплификации движений базальной мембраны. Амплификация (усиление) необходима для улавливания звуков с низким уровнем звукового давления. Важная роль наружных волосковых клеток была продемонстрирована в случаях, когда канамицин (ототоксичный антибиотик) использовался для селективного повреждения наружных волосковых клеток, в то время как внутренние волосковые клетки оставались интактными. Повреждение наружных волосковых клеток приводит к исключению низкого порога чувствительности слухового нерва и его точной настройки, но также влияет и на характеристики высокого порога. Это наблюдение привело к гипотезе, что наружные волосковые клетки являются несущими главную ответственность за амплификацию и точную настройку слуховой системы.
Одним из механизмов усиления является соматическая электроподвижность. Длина изолированных наружных волосковых клеток в ответ на электростимуляцию изменялась на 3-5%. Во время деполяризации наружные волосковые клетки сокращались, в то время как при гиперполяризации возникало их удлинение. В результате наружные волосковые клетки оказывают механическое воздействие, которое приводит базальную мембрану в движение при стимулах до нескольких кГц.
Усиление стимуляции базальной мембраны, в частности, движение стереоцилий в направлении возбуждения и деполяризация наружных волосковых клеток, которые в свою очередь сокращаются, подтягивает базальную мембрану кверху. Таким образом электроподвижность наружных волосковых клеток усиливает подвижность базальной мембраны, вызванную бегущей волной.
Полагают, что престин является белком, ответственным за соматическую электроподвижность в наружных волосковых клетках. Это подтверждается рядом доказательств. Во-первых, при трансфекции престином клетки демонстрируют электроподвижность с амплитудой (размером) до 0,2 мкм, что говорит о необходимости перстина для движения. Во-вторых, престин обнаруживается в нужном месте, а именно в латеральной мембране волосковых клеток. И, наконец, прецизионное удаление или модификация престина у мышиной модели влияло на чувствительность улитки и указывало на необходимость престина для электроподвижности наружных волосковых клеток. Престин относится к суперсемейству передачи анионов SCL26, которые могут быть посредниками электронейтрального обмена хлорида и карбоната через плазматическую мембрану.
Точный механизм, благодаря которому работает этот насос, по-прежнему неизвестен, однако можно предположить, что белковый насос, работающий на принципах изменения напряжения, способен работать гораздо быстрее, чем классическая АТФ-зависимая помпа. В настоящее время, согласно рабочей гипотезе, внутриклеточные анионы действуют как датчики напряжения, связанные с престином и запускающие конформационные изменения. Гиперполяризация ведет к связыванию анионов с престином, что вызывает увеличение поверхности престина, и приводит к увеличению клетки. С другой стороны, деполяризация ведет к отсоединению анионов, уменьшению поверхности престина и к сокращению клетки. В покое анионы обычно отделены от престина, поэтому белок может трансформироваться.
Другой вероятный источник амплификации связан с активным движением пучка волосковых клеток, вызванного воздействием механической передачи и адаптации. В улитке не относящихся к млекопитающим позвоночных пучки волосковых клеток способны генерировать устойчивые колебательные движения, подобная сеть усиления присутствует в наружных волосковых клетках млекопитающих. Активные движения стереоцилий являются важным механизмом амплификации у немлекопитающих позвоночных, и вполне вероятно, что подобный процесс также используется для усиления или настройки кортиева органа у млекопитающих в тесной связи с электроподвижностью наружных волосковых клеток, управляемой престином.
в) Покровная мембрана кортиева органа. Покровная мембрана является внеклеточной структурой, которая лежит на внутренних и наружных волосковых клетках. Однако, только наиболее высокие стереоцилии волосковых клеток непосредственно вплетены в ее нижнюю часть. Покровная мембрана прикрепляется к внутреннему краю лимба и подвижно связана с поддерживающими клетками, такими как клетка Гензена, посредством микроскопических тяжей, названных трабекулами. Важность покровной мембраны подтверждается тем, что мутация гена, кодирующего покровную мембрану (альфа- и бета-текторин), является причиной выраженной потери слуха как у человека, так и у животных.
Исходя из анатомических наблюдений, вначале полагали, что покровная мембрана действует как простой рычаг, который двигает пучки волосковых клеток, перемещаясь вверх и вниз. Другие модели улитки рассматривали мембрану как простой механический груз или резонансную систему, обладающую массой и упругостью. Недавние исследования показали, что покровная мембрана больше похожа на резонансный гель и участвует в увеличении частотной чувствительности улитки. Вполне вероятно, что все предложенные функции покровной мембраны являются актуальными и так как эта структура, как и большинство структур кортиева органа, изменяет свой размер и форму от основания к вершине, покровная мембрана может также способствовать общей тонотопической организации улитки.
Модель аппарата механоэлектрической передачи стереоцилии.
Изображены известные и предполагаемые компоненты аппарата механоэлектрической передачи. Механоэлектрическая передача.
А. В покое приблизительно 90% каналов передачи закрыты. Молекулярные насосы, основанные на миозине,
поднимаются к вершине стереоцилии и регулируют натяжение концевой связи и связанные с ней структуры для обеспечения более точной чувствительности передаточного аппарата.
Б. увеличение механического натяжения в области концевой связи и связанных с ней структур ведет к открытию каналов передачи и входу катионов, деполяризующих клетку.
Местное увеличение концентрации ионов Са 2+ активирует миозиновые насосы и приводит к скольжению передаточного аппарата, тем самым уменьшается механическое натяжение и открываются каналы передачи.
В. деполяризация наружных волосковых клеток крысы в ответ на умеренное механическое отклонение в 50 нм.
Показан быстрый рост рецепторного потенциала, который позволяет достигнуть максимума в 550 пА в отдельно взятой клетке при максимальной стимуляции.
Представленная схема отображает время быстрой (τ1) и медленной (τ2) адаптации.
Кортиев орган
Кортиев орган — рецепторная часть слухового анализатора, расположенная внутри перепончатого лабиринта. В процессе эволюции возникает на основе структур органов боковой линии.
Воспринимает колебания волокон, расположенных в канале внутреннего уха, и передаёт в слуховую зону коры больших полушарий, где и формируются звуковые сигналы. В кортиевом органе начинается первичное формирование анализа звуковых сигналов.
Содержание
История изучения
Открыт итальянским гистологом Альфонсо Корти (Alfonso Corti; 1822—1876).
Анатомия
Расположение
Кортиев орган располагается в спирально завитом костном канале внутреннего уха — улитковом ходе, заполненном эндолимфой и перилимфой. Верхняя стенка хода прилегает к т. н. лестнице преддверия и называется рейснеровой перепонкой; нижняя стенка, граничащая с т. н. барабанной лестницей, образована основной перепонкой, прикрепляющейся к спиральной костной пластинке.
Структура и функции
К. о. расположен на основной перепонке и состоит из внутренних и наружных волосковых клеток, внутренних и наружных опорных клеток (столбовых, клеток Дейтерса, Клаудиуса, Гензена), между которыми находится туннель, где проходят направляющиеся к основаниям волосковых клеток отростки нервных клеток, лежащих в спиральном нервном ганглии. Воспринимающие звук волосковые клетки располагаются в нишах, образуемых телами опорных клеток, и имеют на поверхности, обращенной к покровной перепонке, по 30—60 коротких волосков. Опорные клетки выполняют также трофическую функцию, направляя поток питательных веществ к волосковым клеткам.
Функция Кортиева органа — преобразование энергии звуковых колебаний в процесс нервного возбуждения.
Физиология
Звуковые колебания воспринимаются барабанной перепонкой и через систему косточек среднего уха передаются жидким средам внутреннего уха — перилимфе и эндолимфе. Колебания последних приводят к изменению взаиморасположения волосковых клеток и покровной перепонки Кортиева органа, что вызывает сгибание волосков и возникновение биоэлектрических потенциалов, улавливаемых и передаваемых в центральную нервную систему отростками нейронов спирального ганглия, подходящими к основанию каждой волосковой клетки.
По другим представлениям, волоски звуковоспринимающих клеток — лишь чувствительные антенны, деполяризующиеся под действием приходящих волн за счёт перераспределения ацетилхолина эндолимфы. Деполяризация вызывает цепь химических превращений в цитоплазме волосковых клеток и возникновение нервного импульса в контактирующих с ними нервных окончаниях. Различающиеся по высоте звуковые колебания воспринимаются различными отделами Кортиевого органа: высокие частоты вызывают колебания в нижних отделах улитки, низкие — в верхних, что связано с особенностями гидродинамических явлений в ходе улитки.
Добавьте ссылки на источники, в противном случае она может быть выставлена на удаление.
Дополнительные сведения могут быть на странице обсуждения.
Таким образом улитка является механическим измерителем АЧХ, и по действию схожа с АЧХ-метром, а не с микрофоном. Это позволяет мозгу сразу реагировать на конкретный звук, а не производить преобразование Фурье математически (на что, впрочем, у него не хватит вычислительных способностей), с целью разложения воспринимаемого звука на отдельные источники.
По поляризации звуковых гармоник можно судить о направлении(угловом) источника звука. Таким образом ухо позволяет получить информацию о амплитуде и поляризации каждой гармоники звуковых колебаний. Для низких частот (десятки герц) ухо и мозг успевают также извлечь информацию о фазе гармоник, что позволяет определить направление (как расстояние от головы по оси, проходящей через уши) низкочастотного колебания, если вычислить разность фаз сигнала от правого и левого уха.
Особенность дополнительного сжатия акустической информации позволяет значительно сократить время на анализ полученных данных. Закрученность улитки позволяет снимать спектр, совмещая октавы, то есть ось частоты в АЧХ звуковых колебаний закручивается, амплитуды октав совмещаются, что даёт возможность значительно сократить количество необходимых информационных каналов. Эта физическая основа слуха служит причиной восприятия музыки человеком.
Читайте также:
- Продольная меланонихия и подногтевая меланома. Диагностика и лечение
- Диагностика субэпендимальной гигантоклеточной астроцитомы по КТ, МРТ
- Операция Гохмана при эпикондилите плеча и ее эффективность
- Восточно-африканский лейшманиоз. Кожный лейшманиоз
- Взаимодействие белков с гистонами при образовании гетерохроматина