Особенности клеточной оболочки грамположительных бактерий
Добавил пользователь Alex Обновлено: 23.01.2026
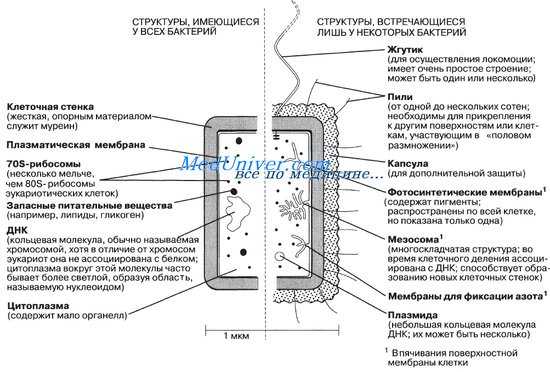
Обязательными органоидами являются: ядерный аппарат, цитоплазма, цитоплазматическая мембрана.
Необязательными (второстепенными) структурными элементами являются: клеточная стенка, капсула, споры, пили, жгутики.
1.В центре бактериальной клетки находится нуклеоид- ядерное образование, представленное чаще всего одной хромосомой кольцевидной формы. Состоит из двухцепочечной нити ДНК. Нуклеоид не отделен от цитоплазмы ядерной мембраной.
2.Цитоплазма- сложная коллоидная система, содержащая различные включения метаболического происхождения (зерна волютина, гликогена, гранулезы и др.), рибосомы и другие элементы белоксинтезирующей системы, плазмиды (вненуклеоидное ДНК), мезосомы (образуются в результате инвагинации цитоплазматической мембраны в цитоплазму, участвуют в энергетическом обмене, спорообразовании, формировании межклеточной перегородки при делении).
3.Цитоплазматическая мембрана ограничивает с наружной стороны цитоплазму, имеет трехслойное строение и выполняет ряд важнейших функций- барьерную (создает и поддерживает осмотическое давление), энергетическую (содержит многие ферментные системы- дыхательные, окислительно- восстановительные, осуществляет перенос электронов), транспортную (перенос различных веществ в клетку и из клетки).
4.Клеточная стенка- присуща большинству бактерий (кроме микоплазм, ахолеплазм и некоторых других не имеющих истинной клеточной стенки микроорганизмов). Она обладает рядом функций, прежде всего обеспечивает механическую защиту и постоянную форму клеток, с ее наличием в значительной степени связаны антигенные свойства бактерий. В составе - два основных слоя, из которых наружный- более пластичный, внутренний- ригидный.
Основное химическое соединение клеточной стенки, которое специфично только для бактерий- пептидогликан (муреиновые кислоты). От структуры и химического состава клеточной стенки бактерий зависит важный для систематики признак бактерий- отношение к окраске по Граму. В соответствии с ним выделяют две большие группы- грамположительные (“грам+”) и грамотрицательные (“грам - “) бактерии. Стенка грамположительных бактерий после окраски по Граму сохраняет комплекс йода с генциановым фиолетовым (окрашены в сине- фиолетовый цвет), грамотрицательные бактерии теряют этот комплекс и соответствующий цвет после обработки и окрашены в розовый цвет за счет докрашивания фуксином.
Особенности клеточной стенки грамположительных бактерий.
Мощная, толстая, несложно организованная клеточная стенка, в составе которой преобладают пептидогликан и тейхоевые кислоты, нет липополисахаридов (ЛПС), часто нет диаминопимелиновой кислоты.
Особенности клеточной стенки грамотрицательных бактерий.
Клеточная стенка значительно тоньше, чем у грамположительных бактерий, содержит ЛПС, липопротеины, фосфолипиды, диаминопимелиновую кислоту. Устроена более сложно- имеется внешняя мембрана, поэтому клеточная стенка трехслойная.
При обработке грамположительных бактерий ферментами, разрушающими пептидогликан, возникают полностью лишенные клеточной стенки структуры- протопласты. Обработка грамотрицательных бактерий лизоцимом разрушает только слой пептидогликана, не разрушая полностью внешней мембраны; такие структуры называют сферопластами. Протопласты и сферопласты имеют сферическую форму (это свойство связано с осмотическим давлением и характерно для всех безклеточных форм бактерий).
L- формы бактерий.
Под действием ряда факторов, неблагоприятно действующих на бактериальную клетку (антибиотики, ферменты, антитела и др.), происходит L- трансформация бактерий, приводящая к постоянной или временной утрате клеточной стенки. L- трансформация является не только формой изменчивости, но и приспособления бактерий к неблагоприятным условиям существования. В результате изменения антигенных свойств (утрата О- и К- антигенов), снижения вирулентности и других факторов L- формы приобретают способность длительно находиться (персистировать) в организме хозяина, поддерживая вяло текущий инфекционный процесс. Утрата клеточной стенки делает L- формы нечувствительными к антибиотикам, антителам и различным химиопрепаратам, точкой приложения которых является бактериальная клеточная стенка. Нестабильные L- формы способны реверсировать в классические (исходные) формы бактерий, имеющие клеточную стенку. Имеются также стабильные L- формы бактерий, отсутствие клеточной стенки и неспособность реверстровать которых в классические формы бактерий закреплены генетически. Они по ряду признаков очень напоминают микоплазмы и другие молликуты- бактерии, у которых клеточная стенка отсутствует как таксономический признак. Микроорганизмы, относящиеся к микоплазмам- самые мелкие прокариоты, не имеют клеточной стенки и как все бактериальные бесстеночные структуры имеют сферическую форму.
К поверхностным структурам бактерий (необязательным, как и клеточная стенка), относятся капсула, жгутики, микроворсинки.
Капсула или слизистый слой окружает оболочку ряда бактерий. Выделяют микрокапсулу, выявляемую при электронной микроскопии в виде слоя микрофибрилл, и макрокапсулу, обнаруживаемую при световой микроскопии. Капсула является защитной структурой (прежде всего от высыхания), у ряда микробов- фактором патогенности, препятствует фагоцитозу, ингибирует первые этапы защитных реакций- распознавание и поглощение. У сапрофитов капсулы образуются во внешней среде, у патогенов- чаще в организме хозяина. Существут ряд методов окраски капсул в зависимости от их химического состава. Капсула чаще состоит из полисахаридов (наиболее распространенная окраска- по Гинсу), реже- из полипептидов.
Жгутики. Подвижные бактерии могут быть скользящие (передвигаются по твердой поверхности в результате волнообразных сокращений) или плавающие, передвигающиеся за счет нитевидных спирально изогнутых белковых (флагеллиновых по химическому составу) образований- жгутиков.
По расположению и количеству жгутиков выделяют ряд форм бактерий.
1.Монотрихи- имеют один полярный жгутик.
2.Лофотрихи- имеют полярно расположенный пучок жгутиков.
3.Амфитрихи- имеют жгутики по диаметрально противоположным полюсам.
4.Перитрихи- имеют жгутики по всему периметру бактериальной клетки.
Способность к целенаправленному движению (хемотаксис, аэротаксис, фототаксис) у бактерий генетически детерминирована.
Фимбрии или реснички - короткие нити, в большом количестве окружающую бактериальную клетку, с помощью которых бактерии прокрепляются к субстратам (например, к поверхности слизистых оболочек). Таким образом, фимбрии являются факторами адгезии и колонизации.
F- пили (фактор фертильности) - аппарат конъюгации бактерий, встречаются в небольшом количестве в виде тонких белковых ворсинок.
Эндоспоры и спорообразование.
Спорообразование- способ сохранения определенных видов бактерий в неблагоприятных условиях среды. Эндоспоры образуются в цитоплазме, представляют собой клетки с низкой метаболической активностью и высокой устойчивостью (резистентностью) к высушиванию, действию химических факторов, высокой температуры и других неблагоплиятных факторов окружающей среды. При световой микроскопии часто используют метод выявления спор по Ожешко. Высокая резистентность связана с большим содержанием кальциевой соли дипиколиновой кислоты в оболочке спор. Расположение и размеры спор у различных микроорганизмов отличается, что имеет дифференциально- диагностическое (таксономическое) значение. Основные фазы “жизненного цикла” спор- споруляция (включает подготовительную стадию, стадию предспоры, образования оболочки, созревания и покоя) и прорастание, заканчивающееся образованием вегетативной формы. Процесс спорообразования генетически обусловлен.
Некультивируемые формы бактерий.
У многих видов грамотрицательных бактерий, не образующих спор, существует особое приспособительное состояние- некультивируемые формы. Они обладают низкой метаболической активностью и активно не размножаются, т.е. не образуют колоний на плотных питательных средах, при посевах не выявляются. Обладают высокой устойчивостью и могут сохранять жизнеспособность в течение нескольких лет. Не выявляются классическими бактериологическими методами, обнаруживаются только при помощи генетических методов ( полимеразной цепной реакции- ПЦР).
Особенности клеточной оболочки грамположительных бактерий
• Грамположительные бактерии обладают толстой клеточной стенкой, содержащей много слоев пептидогликана
• Существенным компонентом клеточной стенки грамположительных бактерий являются тейхоевые кислоты, однако точная их функция неизвестна
• Многие белки клеточной поверхности грамположительных бактерий ковалентно связаны с мембранными липидами или с пептидогликаном
• Mycobacteria характеризуются наличием специфических компонентов клеточной оболочки, богатых липидами
Как было отмечено ранее, клеточные оболочки грамотрицательных и грамположительных бактерий представляют собой различные структуры. Основное различие в составе их клеточных стенок состоит в том, что по сравнению с грамотрицательными бактериями грамположительные характеризуются более толстым слоем пептидогликана. Кроме того, наряду с пептидогликаном, в клеточной стенке грамположительных бактерий содержится еще один важный класс макромолекулярных полимеров.
Как правило, эти полимеры являются полианионами. К числу хорошо известных и наиболее изученных из них принадлежат тейхоевые кислоты. Рисунок ниже иллюстрирует, что тейхоевые кислоты имеют простую повторяющуюся структуру, состоящую из полимера, содержащего фосфорилированные формы сахаров или остатков глицерина. Связывание полимера с клеткой происходит одним из двух способов. Липотейхоевые кислоты связываются с остатками жирных кислот цитоплазматической мембраны, а тейхоевые кислоты клеточной стенки связываются с пептидогликаном посредством линкерных групп.
В разных бактериях и даже в родственных штаммах одного вида обнаружено огромное количество различных повторяющихся единиц. Полимеры имеют общее свойство — все они являются полианионами, что обусловливается наличием в их составе фосфатных групп. Однако фосфат не играет критической роли, и его отсутствие может заменяться другой группой, например глюкоуронатом. При этом образуется тейхоуроновая кислота. Так же как и пептидогликаны, тейхоевые кислоты синтезируются в цитоплазме, через предшественник, связанный с УДФ, и затем транспортируются вне клетки для сборки.
Тейхоевые кислоты необходимы для жизнедеятельности клетки. Об этом свидетельствует тот факт, что делеция генов, участвующих в их синтезе, приводит к гибели бактерий. Полностью их функции не выяснены, хотя не исключено, что они каким-то образом связаны с поддержанием на клеточной стенке электрического заряда. Также предполагается, что они участвуют в захвате клеткой двухвалентных катионов, контролируют активность ферментов автолиза, действующих на пептидогликан, или обеспечивают общую проницаемость клеточной стенки.
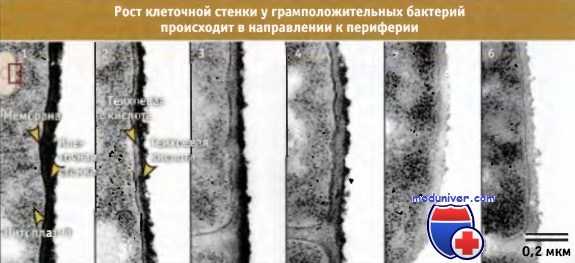
Эксперимент, демонстрирующий постепенное построение новой клеточной стенки у В. subtilis при переходе от среды, дефицитной по магнию, к среде с недостатком фосфата.
Предсуществующая клеточная стенка (содержащая тейхоевую кислоту) постепенно,
на всем протяжении равномерно замещается новосинтезированным материалом (содержащим теихоуроновую кислоту).
У грамположительных бактерий с отсутствующей наружной мембраной существует два механизма удержания поверхностных белков, с тем чтобы не допустить их потери. Один механизм представлен на рисунке ниже.
Набор белков, секретируемых по общему секреторному пути (Sec), подвергается расщеплению специфической сигнальной пептидазой «типа II». До их расщепления фермент фосфатидилг-лицерин: пролипопротеин диацилглицерил трансфераза присоединяет белок к фосфофипидам на наружной стороне цитоплазматической мембраны. Белки, предназначенные для этой «липомодификации», узнаются по наличию сигнала сортинга, который содержит цистеиновый остаток сразу же после сайта расщепления сигнальной пептидазой. Ковалентное связывание белка с наружной поверхностью цитоплазматической мембраны предотвращает его отщепление.
У грамположительных бактерий большое количество белков подвергается модификации с участием такого механизма. Например, считается, что у В. subtilis существует более 100 подобных белков. (Хотя у грамположительных бактерий этот механизм чаще используется для поверхностно-связанных белков, липомодифицированные белки также присутствуют у грамотрицательных бактерий.)
Второй механизм удержания белков на клеточной поверхности у грамположительных бактерий состоит в связывании белка с пептидогликаном клеточной стенки. Вообще говоря, многие белки, связанные с клеточной стенкой, по-видимому, участвуют во взаимоотношениях между патогеном и организмом хозяина (хотя это предположение может являться следствием систематической ошибки секвенирования генома патогенного микроорганизма). Ключевым ферментом, принимающим участие в этом процессе, является сортаза. Известно большое количество субстратов этого фермента.
Наиболее хорошо изучен белок А из Staphylococcus aureus. У белков, прикрепленных к клеточной стенке, на N- и С-концевых участках находятся узнаваемые последовательности. На N-концевом участке присутствует классический расщепляемый сигнальный пептид, необходимый для экспорта белка через цитоплазматическую мембрану. На С-конце, за вторым гидрофобным доменом расположены несколько в основном положительно заряженных аминокислотных остатков. По-видимому, во время работы сортазы этот домен поддерживает С-концевой участок в непосредственной близости от цитоплазматической мембраны.
Сайт узнавания сортазой расположен непосредственно перед С-концевым гидрофобным участком и представляет собой короткую последовательность, обычно Leu-Pro-X-Thr-Gly, где X может быть любая аминокислота. В белке расщепляется связь между четвертым и пятым остатками этой последовательности аминокислот, и вероятно, при действии сортазы, он связывается с пептидогликаном амидной связью, хотя пока такой механизм однозначно не продемонстрирован.
Как иллюстрирует рисунок ниже, синтез клеточной стенки у грамположительных бактерий сопровождается наращиванием нового материала, которое происходит в направлении изнутри-кнаружи. По мере созревания пептидогликана, он мигрирует к внешней стороне, пока не достигнет поверхности, где высвобождается за счет автолиза. Согласно современным представлениям, пептидогликан в релаксированном состоянии включается в состав стенки и, по мере миграции кнаружи, за счет роста клетки, постепенно растягивается. В конце концов, по мере увеличения растяжения, материал оказывается под нагрузкой, и в этот момент наступает гидролиз, после чего продолжается дальнейшее построение клеточной стенки.
Остается неразрешенным важный вопрос, каким образом ферменты автолиза регулируют созревание пептидогликана и рост клетки? Также не ясно, насколько модель синтеза пептидогликана «3 вместо 1», объясняющая наращивание клеточной стенки у грамотрицательных бактерий, приложима к многослойной клеточной стенке грамположительных бактерий.
Даже у таких хорошо известных микроорганизмов, как B.subtilis и E.coli, детали строения клеточной стенки исследованы недостаточно. Причем это несмотря на то, что в основе действия наиболее распространенных антибиотиков (особенно р-лактамов и гликопептидов) лежит ингибирование синтеза клеточной стенки бактерий. В основном это связано с тем, что внимание исследователей было сосредоточено на изучении биохимии взаимодействия ингибиторов с препаратами очищенных ферментов. К числу неизученных относится важнейший вопрос, касающийся механизма синтеза пептидо-гликана с участием РВР, учитывая трехмерную структуру клеточной стенки растущей бактерии.
Для разных микроорганизмов структура клеточной стенки существенно варьирует. Два крайних случая подобной специализации можно проиллюстрировать микоплазмой, у которой вообще отсутствует клеточная стенка, и микобактериями, для которых характерно присутствие сложных жирных кислот, ковалентно связанных с пептидогликаном.
Тейхоевые кислоты являются компонентами клеточной стенки грамположительных бактерий.
Они закрепляются на поверхности клетки либо через цитоплазматическую мембрану, либо через пептидогликан. У грамположительных бактерий один из механизмов привязки белков
с клеточной поверхностью заключается в присоединении их к фосфолипидам плазматической мембраны.
Жирные длинноцепочечные кислоты обозначены буквами R1, R2 и R3. У грамположительных бактерий некоторые белки клеточной поверхности связываются с пептидогликаном клеточной стенки.
Предполагается, что фермент сортаза расщепляет связь Thr-GLy и ковалентно закрепляет белок на клеточной стенке.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Строение клетки бактерий. Структура
Жёсткая бактериальная клеточная стенка придаёт микроорганизмам определённую форму, а также служит механическим барьером, предохраняющим клетку от воздействия факторов окружающей среды. Клеточная стенка грамположительных бактерий состоит из толстого слоя пептидогликана и клеточной мембраны, в то время как у грамотрицательных микроорганизмов — из трёх слоев: внутренней, внешней мембраны и тонкого слоя пептидогликана.
Клеточная стенка микобактерий также содержит большое количество липидных веществ, некоторые из которых обладают иммунореактивностью.
По форме бактерии подразделяют на кокки (округлой формы), бациллы (палочковидной формы) и коккобациллы (промежуточная форма). Кроме того, различают изогнутые и спиралевидные бактерии. Наиболее важные структурные компоненты бактериальной клетки:
• капсула — слизистое аморфное образование, состоящее из полисахаридов и защищающее клетку от фагоцитоза и высыхания;
• липополисахариды — мощные стимуляторы выброса цитокинов, защищающие грамотрицательные бактерии от цитолитических компонентов комплемента;
• фимбрии (пили) — тонкие нитевидные белковые органеллы, участвующие в адгезии (прикрепление к клеткам организма хозяина) и расселении микроорганизмов (факторы колонизации). Например, фимбрии (Р-фимбрии) уропатогенных штаммов Escherichia coli избирательно связываются с маннозными рецепторами эпителия мочеточников.
Антигены фимбрии часто обладают иммуногенностью, индивидуальной для каждого штамма (например, Neisseria gonorrhoeae), чем можно объяснить возникновение рецидивирующих инфекций;
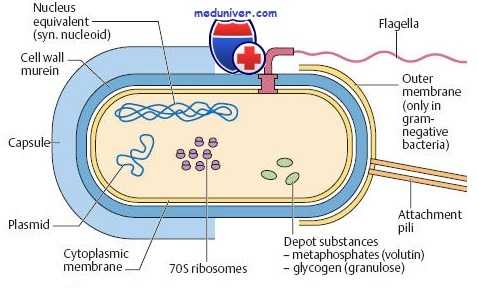
• жгутики — органы движения бактерий, позволяющие микроорганизмам передвигаться в поисках источников питания и проникать через слизистые оболочки организма хозяина. Жгутики (один или несколько) могут располагаться на полюсах (полярное расположение) или по всей поверхности (перитрихи) бактериальной клетки. У некоторых видов (например, у определённых штаммов Treponema) жгутики закреплены внутри клеточной стенки;
• слизь — полисахаридная субстанция, секретируемая некоторыми бактериями и предохраняющая их от воздействия антибиотиков и иммунной системы хозяина;
• споры — метаболически неактивные формы бактерий, образующиеся в неблагоприятных условиях и позволяющие микроорганизмам долгое время выдерживать воздействие механических, температурных и химических факторов окружающей среды (до наступления благоприятных условий).
Бактерий относят к прокариотам, так как они не имеют ядра и содержат только одну хромосому. Процессы спирализации и суперспирализации ДНК микроорганизмов катализирует ДНК-гираза, что позволяет хранить большой объём хромосомного материала.
Рибосомы микроорганизмов отличаются от рибосом эукариотов, что делает их мишенями для антибактериальных препаратов. Более того, бактерии имеют дополнительную ДНК, содержащуюся в плазмидах. Кроме того, в них часто заключена генетическая информация о факторах патогенности.
Видео строение бактериальной клетки - анатомия бактерии
Особенности клеточной стенки грамположительных и грамотрицательных бактерий
Цитоплазматическая мембрана является полифункциональной структурой. Выполняет следующие функции:
· воспринимает всю химическую информацию, поступающую в клетку из внешней среды;
· является основным осмотическим барьером, благодаря которому внутри клетки поддерживается определенное осмотическое давление;
· совместно с клеточной стенкой участвует в регуляции роста и клеточного деления бактерий;
· участвует в регуляции процессов репликации и сегрегации хромосом и плазмид;
· в цитоплазматической мембране содержится значительное количество ферментов, в том числе системы переноса электронов;
· участвует в процессах транспорта питательных веществ в клетку и продуктов жизнедеятельности из клетки в окружающую среду;
· участвует в синтезе компонентов клеточной стенки и образовании мезосом;
· с цитоплазматической мембраной связаны жгутики и аппарат регуляции их движения.
Цитоплазма бактерий представляет собой сложную коллоидную систему. Она неподвижна. В цитоплазме располагаются ядерный аппарат – нуклеоид, плазмиды, рибосомы, мезосомы, различные включения.
Нуклеоид состоит из одной замкнутой спиральной нити ДНК. Длина нити – около 1 мм. Это одиночная бактериальная хромосома. В нуклеоиде также находятся РНК – полимераза, основные белки (но не гистоны). Бактериальная хромосома имеет вид бус. Один конец её часто связан с мезосомой.
Плазмиды – внехромосомные генетические структуры бактерий. Представляют собой замкнутую молекулу двухнитчатой ДНК, связанную с белком. Локализуются в цитоплазме клетки или в состоянии интеграции с хромосомой. Приобретение или утрата плазмид приводит к приобретению или утрате одного или нескольких признаков, хотя существуют «немые» плазмиды. Наиболее известны F – плазмида, R – фактор, Col – плазмида.
Мезосомы – мембранные структуры бактерий, выполняющие функцию генерации энергии, аналоги митохондрий эукариот. Способны ассоциироваться с рибосомами. Представляют собой инвагинации цитоплазматической мембраны, на которой локализованы ферменты дыхания.
Рибосомы – внутриклеточные органоиды, осуществляющие синтез белка. Состоят из белка и трех типов РНК, построены из 2 субъединиц. Бактериальные рибосомы не связаны с мембранным аппаратом, имеют константу седиментации 70S.
Для изучения морфологии бактерий из них готовят нативные (прижизненные) препараты и фиксированные мазки. Фиксированные окрашенные препараты позволяют разграничить бактерии, выявлять детали строения микробной клетки, они удобны в практической работе, особенно при изучении патогенных микроорганизмов. Подготовка фиксированных препаратов включает приготовление мазка, высушивание его, фиксацию и окрашивание мазка.
Для приготовления мазка на чистое обезжиренное предметное стекло наносят каплю физиологического раствора, в которую петлей вносят исследуемый материал с плотной питательной среды и распределяют так, чтобы получить тонкий и равномерный мазок диаметром около 1 – 1,5 см. При приготовлении мазков из жидких культур исследуемый материал непосредственно наносится на предметное стекло.
Высушивание мазка проводится на воздухе или в струе тёплого воздуха.
Фиксация мазка производится с целью прикрепления микроорганизмов к стеклу и обеззараживания препарата. Кроме того, фиксированные бактерии лучше воспринимают красители. Для фиксации мазков высушенный препарат проводят 3 – 4 раза через пламя горелки, причём в пламени препарат следует выдерживать не более 2 секунд. Если фиксация проведена правильно, стекло при прикосновении к тыльной поверхности руки тотчас после окончания процесса фиксации слегка обжигает кожу. В некоторых случаях мазки можно фиксировать путем погружения в этиловый спирт на 15 – 20 минут, в смесь равных объёмов этилового спирта и эфира (смесь Никифорова) на 15 – 20 минут, в ацетон на 5 минут.
Окрашивание мазков осуществляется растворами анилиновых красителей. В основе окраски лежат сложные химические и физико – химические процессы взаимодействия компонентов микробной клетки с анилиновыми красителями. Красящая способность последних зависит от наличия у них карбоксильных, серосодержащих, аминогрупп, от заряда микробной клетки. Для окраски микроорганизмов используют, главным образом, основные красители - метиленовый синий, кристаллический фиолетовый, генциан фиолетовый, фуксин, гематоксилин, везувин и др.. Для выявления различных структурных элементов бактериальной клетки применяют нейтральные и кислые красители (нейтральный красный, кислый фуксин, Конго, пикриновая кислота и др.). Отношение микроорганизмов к красителям определяет их тинкториальные свойства.
Различают простые и сложные методы окраски. Первые состоят в окрашивании мазков одним красителем и дают возможность ознакомиться с общей морфологией микробов. Сложные, или дифференцированные, способы окраски производятся различными красителями и применяются для детального изучения структуры клетки, а также для характеристики и дифференциации данного микроба от других.
1. Приготовить мазки из жидкой культуры бактерий и с плотной питательной среды.
2. Окрасить мазок простым способом, для чего на фиксированный препарат нанести водный раствор фуксина (фуксин Пфейффера) на 1 – 2 мин или раствор метиленового синего на 3 – 5 мин, затем краску слить, препарат ополоснуть водой и просушить с помощью фильтровальной бумаги. Микроскопировать с иммерсионной системой и зарисовать.
3. Окрасить мазок дифференцированным методом.
Техника окраски по Граму
а) на фиксированный препарат поместить полоску фильтровальной бумаги и на неё нанести раствор карболового генцианвиолета на 1 – 2 минуты;
б) краситель слить, снять фильтровальную бумагу и обработать препарат в течение 1 – 2 минут раствором Люголя до почернения препарата;
в) слить раствор Люголя и на препарат нанести несколько капель этилового спирта на 30 – 60 сек;
г) промыть водой и докрасить препарат водным фуксином в течение 1 – 2 минут;
д) слить краску, промыть водой, высушить и микроскопировать препарат с иммерсионной системой.
При данном методе одни бактерии окрашиваются в тёмно – фиолетовый цвет (грамположительные), другие – в красный или розовый (грамотрицательные). Сущность метода состоит в том, что клеточная стенка грамположительных бактерий прочно фиксируют комплекс генцианвиолет – раствор Люголя, не обесцвечивается этанолом и потому не воспринимает дополнительный краситель (фуксин). У грамотрицательных микробов комплекс легко вымывается из клетки этанолом, и они окрашиваются дополнительным красителем.
Зарисовать грамположительные и грамотрицательные бактерии.
4. Изучить в готовых препаратах различные формы микроорганизмов: шаровидные, палочковидные, извитые, нитевидные.
Контрольные вопросы
1. Каковы морфологические и структурные особенности строения бактериальной клетки?
2. Какова структура, химический состав и функции клеточной стенки бактерий?
3. Какими отличительными особенностями характеризуются клеточные стенки грамположительных и грамотрицательных бактерий?
4. Какую роль играет цитоплазматическая мембрана?
5. Каковы особенности организации ядерного аппарата бактерий?
6. Чем характеризуются плазмиды?
7. Что собой представляют цитоплазма, мезосомы и рибосомы бактерий?
8. Как и для чего производится фиксация мазков?
9. В чём состоит механизм окраски бактерий и от чего он зависит?
10. С какими целями применяются простые и сложные методы окраски?
11. В чём заключается принцип окраски по Граму?
12. Каковы преимущества исследования бактерий в живом состоянии и в фиксированных окрашенных препаратах?
Клеточная стенка
Клеточная стенка – структурный компонент, присущий только бактериям (кроме микоплазм). Клеточная стенка выполняет следующие функции:
1. Определяет и сохраняет постоянную форму клетки.
2. Защищает внутреннюю часть клетки от действия механических и осмотических сил внешней среды.
3. Участвует в регуляции роста и деления клеток.
4. Обеспечивает коммуникацию с внешней средой через каналы и поры.
5. Несет на себе специфические рецепторы для бактериофагов.
6. Определяет во многом антигенную характеристику бактерий (природу и специфичность О– и К-антигенов).
7. Содержащийся в ее составе пептидогликан наделяет клетку важными иммунобиологическими свойствами (см. ниже).
8. Нарушение синтеза клеточной стенки бактерий является главной причиной их L-трансформации.
Строение клеточной стенки. В ее составе имеется два слоя: наружный – пластичный и внутренний – ригидный. Основу клеточной стенки составляет пептидогликан, который ранее называли муреином (лат. mureus – стенка). Он имеется только у эубактерий (кроме микоплазм). Пептидогликан (см. цв. вкл., рис. 4) включает в себя остов и два набора пептидных цепочек – боковых и поперечных. Остов пептидогликана одинаков у всех бактерий и состоит из чередующихся молекул аминосахаров – N-ацетилглюкозамина (N-АцГлю) и N-ацетилмураминовой кислоты (N-АцМур), связанных между собой ?-гликозидными связями (рис. 5). Боковые цепочки в каждой молекуле пептидогликана представлены набором идентичных тетрапептидов. Поперечные цепочки также представлены набором из идентичных для данной молекулы пептидогликана пентапептидов, содержащих глицин, – пентаглицинов, однако у разных видов бактерий боковые и поперечные пептиды различны. В тетрапептидной боковой цепочке у большинства грамотрицательных бактерий имеется диаминопимелиновая (диаминопимеловая) кислота (ДАП) – уникальный компонент клеточной стенки, обнаруженный только у прокариот. Кроме того, в составе боковых цепочек пептидогликана обнаружены D-аминокислоты (D-аланин, D-глутамин). Боковые тетрапептиды связаны с N-ацетилмураминовой кислотой остова. Связывание боковых тетрапептидов между собой происходит путем образования поперечных пентаглициновых мостиков между D-аланином одной цепи и диаминопимелиновой кислотой (или иной аминокислотой) другого бокового пептида. Наличие двух типов связей (гликозидные и пептидные), которые соединяют субъединицы пептидогликанов, придает этому гетерополимеру структуру молекулярной сети (см. цв. вкл., рис. 4). Благодаря этим связям пептидогликановый слой клеточной стенки образует огромного размера ригидную мешковидную макромолекулу, которая окружает протопласт, уравновешивает его тургорное давление (у E. coli – до 15 атм.) и придает ему определенную постоянную форму. Пептидогликан может разрушаться под действием различных ферментов, а его синтез блокируют бета-лактамные антибиотики.
Связь между N-ацетилмураминовой кислотой и N-ацетилглюкозамином разрушается лизоцимом, связь между N-ацетилмураминовой кислотой и боковым пептидом (его L-аланином) расщепляют амидазы, а связи межпептидные – эндопептидазы. Пентаглициновый мостик стафилококкового пептидогликана разрушается лизостафином. Образование поперечных сшивок между боковыми цепочками тетрапептидов блокируется пенициллинами (бета-лактамными антибиотиками). Это приводит к разрыхлению пептидогликановой сети, следствием чего является осмотический лизис растущих клеток. Пептидогликан, помимо того что он определяет постоянную форму бактерий, обладает следующими важнейшими иммунобиологическими свойствами.
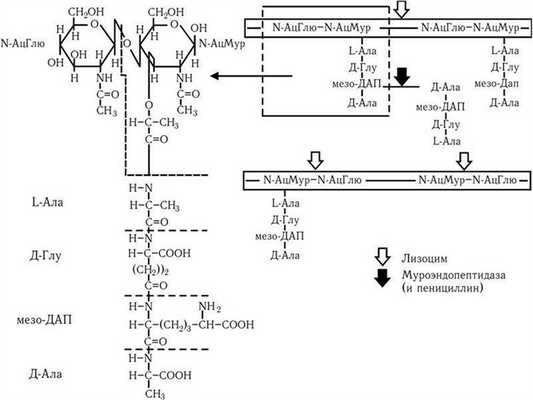
Рис. 5
Стрелками указаны участки молекулы, атакуемые лизоцимом, а также муроэндопептидазой и пенициллином. Объяснение в тексте
1. В его составе обнаружены родоспецифические антигенные детерминанты. Они содержатся в гликановом остове и в тетрапептидах. В межпептидных мостиках имеются видоспецифические антигенные детерминанты.
2. Пептидогликан запускает классический и альтернативный пути активации системы комплемента.
3. Он тормозит фагоцитарную активность макрофагов, т. е. защищает бактерии, особенно грамположительные, от фагоцитоза.
4. Угнетает миграцию макрофагов.
5. Способен индуцировать развитие гиперчувствительности замедленного действия.
6. Обладает противоопухолевым действием.
7. Оказывает пирогенное действие на организм человека и животных.
Таким образом, клеточная стенка является чрезвычайно важной биологической структурой бактерий, определяющей многие их специфические свойства. Как отмечалось выше, все бактерии, в зависимости от их отношения к окраске по Граму, делятся на грамположительные и грамотрицательные. Суть окраски по Граму заключается в том, что вначале бактерии окрашивают кристаллическим или генциановым фиолетовым, а затем – раствором Люголя, после чего мазок обрабатывают спиртом и докрашивают водным фуксином. Грамотрицательные бактерии обесцвечиваются спиртом и поэтому окрашиваются в красный цвет, а грамположительные не обесцвечиваются и сохраняют фиолетовую окраску. Это свойство грамположительных бактерий зависит исключительно от особенностей химического состава и структуры их клеточных стенок, так как при разрушении клеточных стенок или утрате их (в случае L-трансформации) они становятся грамотрицательными.
Причину различного отношения бактерий к окраске по Граму объясняют тем, что после обработки раствором Люголя образуется не растворимый в спирте комплекс йода с генциановым фиолетовым, который у грамположительных бактерий в связи со слабой проницаемостью их стенки не может диффундировать из клетки, в то время как у грамотрицательных легко удаляется при промывании их этанолом, а затем водой.
Читайте также:
- Генетика сложных талассемий. Наследование
- Вытяжение шейного отдела позвоночника. Принципы
- Пластиковые протезы. Непереносимость пластиковых протезов
- Лечение острой левожелудочковой недостаточности. Терапия левожелудочковой недостаточности
- Исходы и осложнения тератомы шеи. Акушерская тактика при тератоме шеи у плода