Репродукция -РНК-вирусов. Репродукция вирусов с двухнитевыми РНК.
Добавил пользователь Владимир З. Обновлено: 27.01.2026
Характерными свойствами вирусов являются: а) наличие одного типа нуклеиновой кислоты; б) способность синтезировать экзотоксины; в) абсолютный паразитизм;
Какие варианты нуклеиновых кислот могут присутствовать в вирусном геноме:
несегментированная двухнитевая ДНК
несегментированный двухнитевый гибрид ДНК –РНК
сегментированная двухнитевая РНК
сегментированная двухнитевая ДНК
сегментированный двухнитевый гибрид ДНК - РНК.
Укажите типы симметрии, по которым организованы нуклеокапсиды вирусных частиц:
асимметричная организация нуклеокапсида
В состав сложных вирусов входят: а) капсид; б) суперкапсид; в) нуклеиновая кислота;
К сложным вирусам относятся: а) вирус полиомиелита; б) вирус кори; в) вирус гриппа;
Какая из перечисленных ниже стадий репликации вирусов не является температурозависимым процессом:
высвобождение нуклеиновой кислоты
интеграция вирусной ДНК в геном клетки
сборка дочерних популяций
высвобождение дочерних популяций
Взаимодействие между какими вирусами может привести к фенотипическому смешиванию:
между близкородственными вирусами с «голыми» капсидами
между «голым» и «одетым» вирусами
между близкородственными ДНК-содержащими вирусами
между любыми вирусами, инфицирующими одну и ту же клетку.
только между ДНК-содержащими вирусами
только между РНК-содержащими вирусами
Какие вирусы могут вызвать персистирующие инфекции:
значительно изменяющие метаболизм клетки
инфицирующие нечувствительные клетки
вирусы, высвобождающиеся почкованием
обусловливает продолжительную защиту
положена в основу применения живых вирусных вакцин
приводит к проникновению в клетку вирусов различных видов
исключает проникновение в клетку вирусов различных видов
Противовирусными препаратами являются: а) антибиотики; б) интерфероны;
В диагностике вирусных инфекций применяют методы: а) вирусологический;
Каким образом можно выявить наличие вируса в заражённой культуре клеток:
по цитопатическим изменениям клеток
по способности эритроцитов адсорбироваться на ЦПМ инфицированных клеток
обнаружением вирусных белков в монослое инфицированных клеток
выявлением изменения питательных потребностей инфицированных клеток
по способности вирусов агглютинировать эритроциты
при электронной микроскопии
Какие из указанных лабораторных методов исследования применяют на практике для диагностики вирусных инфекций:
определение титра противовирусных АТ в парных сыворотках
выделение и идентификация возбудителя в клеточных культурах
выявление Аг и нуклеиновых кислот возбудителей в клинических образцах
оценка иммунного статуса больного
заражение лабораторных животных
гистологическое изучение биоптатов поражённых органов
Присутствие вирусов в культуре ткани выявляют:
по цитопатическому действию на культуру клеток
в реакции гемадсорбции
в реакции Асколи
методом цветной пробы
Укажите наиболее важный фактор, обеспечивающий невосприимчивость к инфицированию патогенными вирусами в популяции:
Общая вирусология с основами таксономии вирусов позвоночных
Основными химическими соединениями, которые входят в состав всех вирусов, являются белки и нуклеиновые кислоты. В состав ряда вирусов входят липиды и углеводы.
Белки. Локализация вирусных белков. Белки, связанные с жизненным циклом вируса, разделяют на белки, детерминируемые геномом вируса и белки, имеющие клеточное происхождение. В качестве примера клеточных белков, которые обнаружены в составе некоторых вирионов, могут быть приведены белок цитоскелета – актин, и ядерные белки – гистоны. Белки клеточного происхождения, участвующие в процессе репликации вируса, будут рассмотрены в разделе взаимодействия вируса с клеткой.
По месту локализации белки, детерминируемые вирусным геномом, разделяют на две группы: 1) структурные белки – это белки, входящие в состав ВЧ, их обозначают как VP; 2) неструктурные белки – это предшественники структурных белков, регуляторные белки и ферменты, обслуживающие процесс внутриклеточной репродукции вируса и не входящие в состав ВЧ. Их обозначают как NS-белки.
Свойства вирусных белков. В состав вирионов входят белки с различной молекулярной массой (от 4 до 100 кД), состоящие из одной или нескольких полипептидных цепей. Количество этих белков также различно у разных вирусов. В состав нуклеокапсида ВТМ входит один белок. У других вирусов в состав вириона может входить несколько десятков белков, имеющих различные физико-химические свойства. Белки, формирующие капсид, нуклеокапсид и коровую оболочку, обладают одним общим свойством – способностью к самосборке.
В состав ВЧ могут входить низкомолекулярные белки, не участвующие в формировании капсида. Например, геномные белки пикорнавирусов и аденовирусов. Геномный белок ковалентно связан с нуклеиновой кислотой и участвует в ее репликации.
Сложные белки представлены гликопротеинами (обозначают как gp) и липопротеинами. Наличие гликопротеина определяет присутствие в вирионе углеводного компонента, который может быть представлен олигосахаридами маннозного типа, галактозой, N-ацетилглюкозамином или нейраминовой кислотой. Вирусные гликопротеины, как правило, экспонированы на наружной поверхности ВЧ и выполняют три основные функции: обеспечивают связывание вириона с клеточным рецептором (функция прикрепительного белка), обладают фузионной активностью (обеспечивают слияние мембран) и определяют антигенные свойства вирусов. В то же время, вирусные гликопротеины могут быть и неструктурными белками и, оставаясь в интегральной форме в мембране шероховатого эндоплазматического ретикулюма (ШЭР), выполнять функции транслоказ, обеспечивая транспорт вирусных компонентов в его просвет.
Вирусные липопротеины представлены белками, ацилированными, как правило, миристиновой (C14) кислотой. Остатки жирных кислот, соединенные с молекулой белка, выполняют функцию липофильного якоря.
Вирусные белки-ферменты могут входить в состав вирусной частицы или являться неструктурными белками и появляться в клетке после экспрессии вирусного генома. Наиболее оснащенным ферментами является вирион вируса оспы, который имеет практически полный набор энзимов, необходимых для независимой внутриклеточной репликации вируса. В то же время, мелкие просто организованные изометрические вирусы с позитивным РНК-геномом могут не иметь никаких ферментов в составе вириона.
Функционально активные белки вирусов представлены, в первую очередь, ферментами нуклеинового обмена, обеспечивающими сложные механизмы репликации/транскрипции вирусного генома; ферментами, осуществляющими посттрансляционный процессинг и модификацию белков, и ферментами, участвующими в проникновении вирионов в клетку хозяина.
Первая группа ферментов наиболее многочисленна и включает как аналоги клеточных ферментов, так и вирус-специфические ферменты.
ДНК-зависимая ДНК-полимераза – осуществляет синтез ДНК на матрице ДНК (вирус оспы).
ДНК-зависимая РНК-полимераза – осуществляет синтез мРНК на матрице ДНК (вирус оспы).
Обратная транскриптаза или ревертаза или РНК-зависимая ДНК-полимераза осуществляет синтез ДНК на матрице РНК. Впервые открыта в 1970 г. у ретровирусов Темином и Мизутани.
Хеликаза – осуществляет расплетете двухнитевой структуры ДНК. Кроме этого хеликазы обладают нуклеотидтрифосфат-зависимой РНК-хеликазной активностью, которая включает три процесса: связывание дезоксинуклеотидтрифосфата, его гидролиз и за счет этой энергии расплетение двухнитевой РНК.
мРНК-модифицирующие ферменты : поли-А-полимераза – аденилирует 3'конец РНК за счет энергии АТФ; Кэп-энзим и метилтрансферазный комплекс – катализирует образование на 5'-конце кэп-структуры.
АТФ-аза, ГТФ-аза – осуществляют гидролиз соответствующих энергетических субстратов.
Рибонуклеаза Н – разрушает РНК, находящуюся в дуплексе с ДНК. Вторая группа вирусных ферментов – ферменты белкового обмена.
Здесь мы приведем лишь некоторые из них:
Протеиназы – ферменты, участвующие в посттрансляционном процессинге полипротеинов. Являются NS-белками РНК-содержащих вирусов;
Протеинкиназы – ферменты, фосфорилирующие структурные белки вирионов. Обнаружены в составе вируса везикулярного стоматита, вируса бешенства, альфавирусов и ретровирусов. Примерами ферментов, участвующих в проникновении вирусов в клетку, являются лизоцим бактериофагов и нейраминидаза вируса гриппа.
Липиды. Все оболочечные РНК-содержащие почкующиеся вирусы имеют липиды клеточного происхождения, входящие в состав суперкапсида (от 15 % до 30 % от сухого веса). От 50 % до 60 % липидов представлены фосфолипидами, от 20 % до 30 % составляет холестерин.
У ДНК-геномных вирусов липиды содержат вирусы оспы, герпеса, гепатита B. Это непочкующиеся вирусы. У вируса оспы липиды не образуют дифференцированной оболочки, которая формируется в цитоплазме в процессе морфогенеза поксвириона. Липиды вируса гепатита B образуются путем инвагинации мембран эндоплазматического ретикулюма (ЭПР). Липидсодержащая оболочка вируса герпеса формируется при прохождении внутреннего компонента вириона через ядерную мембрану. Следовательно, в состав вирусной оболочки герпесвирусов входят липиды ядерной мембраны.
Нуклеиновые кислоты. Клетки всех живых организмов содержат два вида нуклеиновой кислоты – ДНК (двухнитевая ДНК клеточного генома) и РНК (мРНК, тРНК, рРНК). В отличие от клеток, вирионы содержат только один вид нуклеиновой кислоты – ДНК или РНК. И та и другая являются хранителями наследственной информации и выполняют функции генома. Однако следует учитывать, что наличие одного вида нуклеиновой кислоты является характеристикой вириона, но не вируса. В жизненном цикле вируса его геномная нуклеиновая кислота транскрибируется, то есть ДНК-содержащие вирусы образуют РНК. Ряд РНК-содержащих вирусов имеют в цикле репродукции стадию обратной транскрипции и синтезируют ДНК на матрице РНК. Примерно 20 % всех вирусов имеют ДНК-геном, 80 % – РНК-геном. Способность РНК хранить наследственную информацию – уникальное свойство вирусов. Размеры вирусных геномов (длина нуклеотидных последовательностей, выраженная в нуклеотидах) варьируют в широких пределах – от 1,7 тысяч нуклеотидов (т.н.) у цирковируса свиней до 300 т.н. у фикоднавирусов архибактерий.
Кроме того, что геном вирусов может быть представлен или ДНК или РНК, он может находиться в разных видах – в виде двухнитевой (дн) или однонитевой (он) формы, в виде линейной или кольцевой, в виде непрерывной или сегментированной формы.
Многообразие видов РНК геномов расширяется за счет существования последовательностей, отличающихся направлением связей сахаро-фосфатного остова.
Однонитевые РНК могут иметь позитивную полярность – (+) РНК, негативную полярность – (-) РНК или могут быть представлены обоюдозначащей цепью – (+, – ) РНК (амбисенс стратегия кодирования). В свою очередь, РНК позитивной полярности могут иметь разную структурную организацию: могут, являясь матричной РНК, иметь на 5'-конце кэп (7-метилгуанозин, Сар), а на 3'-конце – поли-А (poly-A) последовательность; могут не иметь кэпа или поли-А; могут иметь на 5'-конце геномный белок; могут иметь на 3'-конце тРНК-подобную или шпильковую структуру.
Виды геномов вирусов легли в основу их классификации. Однако следует учитывать, что вид генома в настоящее время не является формальным таксоном и используется для удобства ориентации в многообразии вирусов.
Углеводы. Углеводный компонент вирусов находится в составе гликопротеидов. Количество сахаров в составе гликопротеидов может быть достаточно большим, достигая от 10 % до 13 % от массы вириона. Химическая специфичность их полностью определяется клеточными ферментами, обеспечивающими перенос и присоединение соответствующих сахарных остатков. Обычными сахарными остатками, обнаруживаемыми в вирусных белках, являются фруктоза, сахароза, манноза, галактоза, нейраминовая кислота, глюкозамин. Таким образом, подобно липидам, углеводный компонент определяется клеткой-хозяином, благодаря чему один и тот же вирус, выращенный в клетках разных видов, может значительно отличаться по составу сахаров в зависимости от специфичности клеточных гликозилтрансфераз.
Углеводный компонент гликопротеидов играет существенную роль в структуре и функции белка. Он является каркасом для локальных участков гликопротеида, обеспечивая сохранение конформации белковой молекулы, и обусловливает защиту молекулы от протеаз. Возможны и другие функции углеводов, пока достоверно не установленные.
Компоненты клетки-хозяина. В составе вирионов могут находиться компоненты клетки-хозяина. К таким компонентам могут относиться белки, и даже целые клеточные структуры. Так, например, в составе ряда оболочечных вирусов может находиться белок цитоскелета актин, в составе паповавирусов содержатся клеточные гистоны. Ряд вирусов содержит клеточные ферменты, например, протеинкиназы. В составе аренавирусов обнаружены рибосомы.
Клеточные компоненты могут включаться в вирион случайно или закономерно, В некоторых случаях они играют существенную роль в репродукции вируса, как, например, гистоны в репродукции паповавирусов.
Назовите стадию репликации вирусов, которая не является температурозависимым процессом.
+ Между близкородственными вирусами с "голыми" капсидами.
Между "голым" и "одетым" вирусами.
Между близкородственными ДНК-геномными вирусами.
Между любыми вирусами, инфицирующими одну и ту же клетку.
Только между ДНК- геномными вирусами.
Только между РНК- геномными вирусами.
Назовите типы симметрии, по которым организованы нуклеокапсиды вирусных частиц.
Ассиметричная организация нуклеокапсида.
Какие вирусы могут вызвать персистирующие инфекции?
+ Вирусы, высвобождающиеся почкованием.
Високовирулентные литические вирусы.
Вирусы, которые значительно изменяют метаболизм клетки.
Вирусы, инфицирующие нечувствительные клетки.
18. Какие варианты нуклеиновых кислот могут присутствовать в вирусном геноме?
+ Двухнитевая нефрагментированная ДНК.
+ Двухнитевая фрагментированная РНК (Семейство Reoviridae).
+ Однонитевая нефрагментированная +РНК (Семейство Picornaviridae).
+ Однонитевая нефрагментированная –РНК (Семейство Paramyxoviridae).
+ Однонитевая фрагментированная –РНК (Семейство Orthomyxoviridae).
+ Две идентичные нити +РНК (диплоидный геном – Семейство Retroviridae).
+ Однонитевая кольцевая РНК (дефектный вирус – вирус гепатита-дельта).
Двухнитевой нефрагментованный гибрид ДНК-РНК.
Двухнитевая фрагментированная ДНК.
Двухнитевой фрагментированный гибрид ДНК-РНК.
Каким образом можно выявить наличие вируса в зараженной культуре клеток?
+ По цитопатическому действию вируса на культуру клеток.
+ По способности эритроцитов адсорбироваться на цитоплазматической мембране инфицированных клеток.
+ Обнаружением вирусных белков в монослое инфицированных клеток.
+ При электронной микроскопии.
Правильного ответа нет.
Какие важнейшие факторы обеспечивают резистентность популяции к инфицированию вирусами, имеющими патогенное действие?
+ Формирование иммунной прослойки.
+ Проведение активной иммунопрофилактики.
Невосприимчивость к повторному инфицированию.
Проведение пассивной иммунопрофилактики.
Применение противовирусных химиопрепаратов с профилактической целью.
Какие Вам известны методы лабораторной диагностики вирусных инфекций?
+ Серологический: определение титра противорусных антител в парных сыворотках.
+ Вирусологический: выявление возбудителя в культуре клеток и его идентификация.
Выявление возбудителя и нуклеиновых кислот возбудителя в клиническом материале.
Оценка иммунного статуса больного.
Биологический: инфицирование лабораторных животных.
Гистологическое изучение биоптатов инфицированных органов.
22. Что Вы понимаете под понятием "вирус"?
+ Облигатный внутриклеточный паразит, репродукция которого происходит только в клетке.
Облигатный внутриклеточный паразит, которому характерен бинарный способ деления.
Облигатный внутриклеточный паразит, имеющий клеточное строение.
23. Что Вы понимаете под понятием "вирион"?
+ Элементарная вирусная частица, состоящая из нуклеоида и капсида.
+ Конечная фаза развития вируса.
+ Внеклеточная стадия вируса в состоянии покоя.
Понятие "вирион" идентично понятию "вирус".
Кому из ученых принадлежит приоритет открытия вирусов и кто из ученых является основателем вирусологии?
Репродукция -РНК-вирусов. Репродукция вирусов с двухнитевыми РНК.
Вирусы. Вирион. Морфология вирусов. Размеры вирусов. Нуклеиновые кислоты вирусов.
Вирусы [от лат. virus, яд] — наименьшие по размерам агенты, имеющие геном, окружённый белковой оболочкой. Вирусы не воспроизводятся самостоятельно, они — облигатные внутриклеточные паразиты, репродуцирующиеся только в живых клетках. Все вирусы существуют в двух формах. В настоящее время известны вирусы бактерий (бактериофаги), грибов, растений и животных.
Внеклеточная форма — вирион — включает в себя все составные элементы (капсид, нуклеиновую кислоту, структурные белки, ферменты и др.). Внутриклеточная форма — вирус — может быть представлена лишь одной молекулой нуклеиновой кислоты, так как, попадая в клетку, вирион распадается на составные элементы.
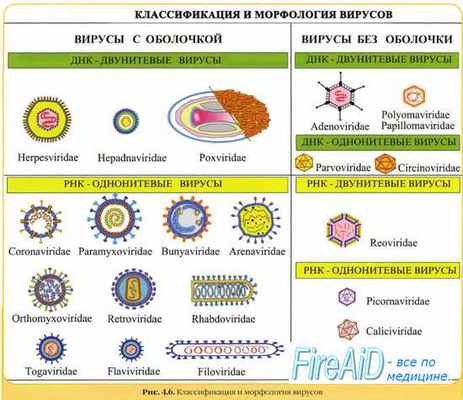
Морфология вирусов. Размеры вирусов.
Несмотря на внутриклеточный паразитизм, среди вирусов имеются крупные виды, соизмеримые по размерам с микоплазмами и хламидиями. Например, вирус натуральной оспы достигает 400 нм и вполне сравним с риккетсиями (300-500 нм) и хламидиями (300-400 нм). По морфологии выделяют вирусы палочковидные (например, возбудитель лихорадки Эбола), пуле-видные (вирус бешенства), сферические (герпесвирусы), овальные (вирус оспы), а также бактериофаги, имеющие сложную форму (рис. 2-1). При всём разнообразии конфигураций, размеров и функциональных характеристик вирусам присущи некоторые общие признаки. В общем виде зрелая вирусная частица (вирион) состоит из нуклеиновой кислоты, белков и липидов, либо в его состав входят только нуклеиновые кислоты и белки.
Нуклеиновые кислоты вирусов
Вирусы содержат только один тип нуклеиновой кислоты, ДИК или РНК, но не оба типа одновременно. Например, вирусы оспы, простого герпеса, Эпстайна-Барр — ДНК-содержащие, а тогавирусы, пикорнавирусы — РНК-содержащие. Геном вирусной частицы гаплоидный. Наиболее простой вирусный геном кодирует 3-4 белка, наиболее сложный — более 50 полипептидов. Нуклеиновые кислоты представлены однонитевыми молекулами РНК (исключая реовиру-сы, у которых геном образован двумя нитями РНК) или двухнитевыми молекулами ДНК (исключая парвовирусы, у которых геном образован одной нитью ДНК). У вируса гепатита В нити двухнитевой молекулы ДНК неодинаковы по длине.
Вирусные ДНК образуют циркулярные, ковалентно-сцёпленные суперспирализованные (например, у паповавирусов) или линейные двухнитевые структуры (например, у герпес- и аденовирусов). Их молекулярная масса в 10-100 раз меньше массы бактериальных ДНК. Транскрипция вирусной ДНК (синтез мРНК) осуществляется в ядре заражённой вирусом клетки. В вирусной ДНК на концах молекулы имеются прямые или инвертированные (развёрнутые на 180") повторяющиеся нуклеотидные последовательности. Их наличие обеспечивает способность молекулы ДНК замыкаться в кольцо. Эти последовательности, присутствующие в одно- и двух-нитевых молекулах ДНК, — своеобразные маркёры вирусной ДНК.
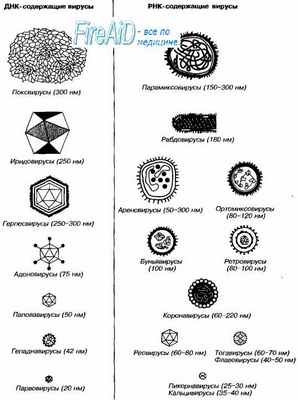
Рис. 2-1. Размеры и морфология основных возбудителей вирусных инфекций человека.
Вирусные РНК представлены одно- или двухнитевыми молекулами. Однонитевые молекулы могут быть сегментированными — от 2 сегментов у ареновирусов до 11 — у ротавирусов. Наличие сегментов ведёт к увеличению кодирующей ёмкости генома. Вирусные РНК подразделяют на следующие группы: плюс-нити РНК (+РНК), минус-нити РНК (-РНК). У различных вирусов геном могут образовывать нити +РНК либо -РНК, а также двойные нити, одна из которых -РНК, другая (комплементарная ей) — +РНК.
Плюс-нити РНК представлены одиночными цепочками, имеющими характерные окончания («шапочки») для распознавания рибосом. К этой группе относят РНК, способные непосредственно транслировать генетическую информацию на рибосомах заражённой вирусом клетки, то есть выполнять функции мРНК. Плюс-нити выполняют следующие функции: служат мРНК для синтеза структурных белков, матрицей для репликации РНК, упаковываются в капсид с образованием дочерней популяции. Минус-нити РНК не способны транслировать генетическую информацию непосредственно на рибосомах, то есть они не могут функционировать как мРНК. Однако такие РНК служат матрицей для синтеза мРНК.
Инфекционность нуклеиновых кислот вирусов
Многие вирусные нуклеиновые кислоты инфекционны сами по себе, так как содержат всю генетическую информацию, необходимую для синтеза новых вирусных частиц. Эта информация реализуется после проникновения вириона в чувствительную клетку. Инфекционные свойства проявляют нуклеиновые кислоты большинства +РНК- и ДНК-содержащих вирусов. Двухнитевые РНК и большинство -РНК не проявляют инфекционных свойств.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Как остановить операцию «Коронавирус»?
В ведущих мировых военных биологических лабораториях и институтах методами генетической инженерии создаются специализированные вирусы разного назначения, которые, в частности, стали эффективным оружием первой мировой биологической войны. Современные методы борьбы с вирусными эпидемиями, в частности с коронавирусной инфекцией, не позволяют избежать втягивания нашей страны в пагубную для национальной экономики кампанейщину по организации карантинов, которую навязывает ВОЗ.
Мы предлагаем взглянуть на ситуацию с иного ракурса. На наш взгляд, с вирусами самых разных модификаций следует обращаться адекватными методами, применяя инструментарий, соответствующий их природе.
Вирус является информационно-энергетической субстанцией со своей собственной целевой жизненной программой. Если вирус – это информационная программа, то можно ли дистанционно и бесконтактно изменить эту программу, применяя информационно-энергетический инструментарий? В рамках данного текста мы попытаемся дать ответ на этот вопрос, выдвинув позитивную рабочую гипотезу.
1. О происхождении и природе вирусов.
Вирусы существуют в двух формах – внеклеточной и внутриклеточной. Для обозначения внеклеточной формы вируса введен термин «вирион». Строение вириона несхоже со строением клеток и совершенно иной тип размножения, названный дисъюнктивной репродукцией (1).
Дисъюнктивная репродукция – это временная и территориальная разобщенность синтеза вирусных компонентов – генетического материала и белков – от последующей сборки и формирования вирионов, что напоминает бинарные отравляющие вещества.
Вирион состоит из нуклеиновой кислоты и капсида (оболочки, состоящей из белка и, реже, липидов), находясь вне живой клетки. Вирионы большинства вирусов не проявляют никаких признаков биологической активности, пока не соприкоснутся с клеткой-хозяином, после чего образуют комплекс «вирус—клетка», способный жить и «производить» новые вирионы. При заражении клетки вирион либо вводит в клетку-хозяина только свой геном (например, бактериофаги), либо проникает в клетку практически полностью (большинство других вирусов). Белки, остающиеся от вириона на поверхности клетки, служат «мишенью» для иммунной системы так же, как и сами вирионы.
«Клетки всех живых организмов содержат два вида нуклеиновой кислоты – ДНК (двухнитевая ДНК клеточного генома) и РНК (мРНК, тРНК, рРНК). В отличие от клеток, вирионы содержат только один вид нуклеиновой кислоты – ДНК или РНК. И та и другая являются хранителями наследственной информации и выполняют функции генома. Однако следует учитывать, что наличие одного вида нуклеиновой кислоты является характеристикой вириона, но не вируса. В жизненном цикле вируса его геномная нуклеиновая кислота транскрибируется, то есть ДНК-содержащие вирусы образуют РНК. Ряд РНК-содержащих вирусов имеют в цикле репродукции стадию обратной транскрипции и синтезируют ДНК на матрице РНК. Примерно 20 % всех вирусов имеют ДНК-геном, 80 % – РНК-геном. Способность РНК хранить наследственную информацию – уникальное свойство вирусов» (1).
Основным и абсолютным критерием отличия вирусов от всех других форм жизни является отсутствие у них собственных синтезирующих белок систем. Таким образом, вирусы не являются организмами, пусть даже мельчайшими, так как любые, даже мельчайшие организмы типа микоплазм и хламидий имеют собственные белоксинтезирующие системы. По определению академика В.М. Жданова, вирусы являются автономными генетическими структурами, способными функционировать только в клетках (с разной степенью зависимости от клеточных систем синтеза нуклеиновых кислот и полной зависимостью от клеточных белоксинтезирующих и энергетических систем) и подвергающимися самостоятельной эволюции.
С точки зрения паразитологии, вирусы – облигатные (то есть обязательные) внутриклеточные паразиты. Внутриклеточный паразитизм – это высшая стадия облигатного паразитизма, суть которого заключается в абсолютной зависимости метаболизма паразита от организма хозяина и характеризуется полной невозможностью размножения паразита за пределами клетки. Однако уровень паразитизма вирусов качественно иной, чем у внутриклеточных паразитов-микроорганизмов. Вирусы – это генетические паразиты. Крайним проявлением генетического паразитизма является способность ряда вирусов интегрироваться в геном клетки хозяина.
Наиболее популярная на сегодняшний день гипотеза происхождения вирусов предполагает их происхождение от генетических элементов клеток, ставших автономными. Эту гипотезу иронически назвали гипотезой «взбесившихся генов». Согласно этой гипотезе, вирусы являются дериватами генетических элементов клеток, но они возникали и эволюционировали вместе с возникновением и эволюцией клеточных форм жизни. «Природа как бы испробовала на вирусах все возможные формы генетического материала (разные виды РНК и ДНК), прежде чем окончательно остановила свой выбор на канонической его форме – двунитчатой ДНК, общей для всех клеточных форм организмов, начиная от бактерии и кончая человеком. Будучи, с одной стороны, автономными генетическими структурами, с другой стороны, неспособными развиваться вне клеток, вирусы на протяжении миллиардов лет биологической эволюции проделали настолько разнообразные пути развития, что отдельные их группы не имеют преемственной связи между собой. По-видимому, разные группы вирусов возникали в исторически разные времена из разных генетических элементов клеток и поэтому существующие в настоящее время разные группы вирусов имеют полифилетическое происхождение, т.е. не имеют единого общего предка» (1).
Вирусы могут проходить через самые микроскопические поры, задерживающие бактерии, так как их размеры на несколько порядков меньше, чем размеры самых мелких микроорганизмов. Размеры вирусных частиц, в свою очередь, колеблются в относительно широких пределах. Самые мелкие вирусы имеют диаметр чуть больше 20 нм (парвовирусы, пикорнавирусы), вирусы средних размеров – от 100 до 150 нм (аденовирусы, коронавирусы).
2. Концепция «вирусного генератора».
Группа российских ученых из ГУП «Научно-исследовательский институт новых медицинских технологий» выдвинула гипотезу «вирусного генератора», то есть испускания одноклеточными организмами, находящимися в непосредственном контакте (в составе с организмом-хозяином) с многоклеточным организмом, вирусов. Этот гипотетический процесс, как предполагается, интенсифицируется при облучении одноклеточного организма электромагнитным излучением (ЭМИ) как ответ на «информационное раздражение» – сигнал опасности (2).
В конце XIX века Дм. Ивановский, а затем Мартин Бейеринк, исследуя табачную мозаику, установили, что вирусы не есть «микробактерии» [4]. На сегодняшний день вирусы окончательно и однозначно не отнесены к миру живого или неживого. Таким образом, при всех серьезных достижениях медицинской вирусологии, на уровне теоретической вирусологии не решены фундаментальные вопросы определения сущности вирусов.
Основным моментом в гипотезе «вирусного генератора» является признание информационной доминанты в природе вирусов.
Главный вывод из этой концепции состоит в констатации того факта, что вирусы не являются паразитическими образованиями, не являются и некими автономно существующими живыми системами, а есть специфическое средство борьбы за существование в конгломерате одноклеточных живых существ.
Согласно данным современной палеонтологии, живая материя появилась на Земле 3,5–3,8 млрд лет тому назад в форме одноклеточных организмов. Вначале это были гетеротрофные бактерии, затем появились фотосинтезирующие цианобактерии (сине-зеленые водоросли), а около 1,2 млрд лет назад – простейшие. На рубеже 800 млн – 1 млрд лет тому назад начали формироваться колонии одноклеточных микроорганизмов, давшие начало многоклеточным. Таким образом, первые 2,5–3 млрд лет земная биосфера состояла только из одноклеточных. Будучи живыми, первоначально возникшие одноклеточные, как и существующие в наше время их потомки, обладали всеми атрибутами, соответствующими сущности живого – динамическому самосохранению. Им был присущ обмен веществ и энергии, они размножились, эволюционировали, воспринимали, перерабатывали и использовали информацию, боролись за свое существование. О грандиознейших масштабах борьбы за существование свидетельствуют факты, связанные с совпадением у них смерти индивидуального организма и его размножения в форме деления на два дочерних организма. Вследствие этого за время своего монопольного существования на Земле одноклеточные настолько «перенаселили» земную поверхность, что борьба «не на жизнь, а на смерть» неизбежно превратилась в специфический атрибут их существования. Ясно, что грандиозной масштабности анализируемой борьбы не может соответствовать один только фагоцитоз, т.е. поглощение мелких организмов более крупными одноклеточными собратьями. Должно существовать универсальное средство, обеспечивающее борьбу «всех против всех» в мире микроорганизмов. На наш взгляд, таким средством стала генерация вирусов.
Одноклеточным организмам внутренне присуще формирование вирусов как информационного средства борьбы за существование и стремление, порождая их, поражать другие организмы. Механизм формирования и использования вирусов – непременный атрибут бытия одноклеточных.
В организмах многоклеточных организмов этот процесс генерации вирусов мощно подавлен для того, чтобы клетки многоклеточного не уничтожали друг друга.
Предметом исследования в рамках концепции вирусного генератора являются отношения между вирусоизлучающими одноклеточными микроорганизмами и организмами многоклеточными. Авторы концепции считают, что избавление человечества от наиболее тяжелых вирусных заболеваний лежит не на пути «подавления» уже размножившихся в организме вирусов (так поступает современная медицинская вирусология), а в предупреждении инициации и развития цепной реакции размножения вирусов в организме, то есть в управлении отношениями одноклеточных и многоклеточных.
3. Биополе ДНК и вирусов.
Целевая программа вируса заложена в его ДНК или РНК. ДНК управляет и управляется биополем. Вдоль ДНК движутся электромагнитные волны, образующие солитоны, структурно устойчивые одиночные волны, распространяющиеся в нелинейной среде. Экспериментально подтверждено резонансное полевое взаимодействие молекул ДНК в гига-герцевом диапазоне. Обнаруженные частоты резонансного поглощения зависят от длины и топологии ДНК и лежат в районе 2-9 ГГц (3).
Биологи и иммунологи хорошо изучили механизм индивидуальной иммунной системы, но ничего определенного не могут сказать о коллективном иммунитете, о том, как и за счет чего происходит коллективная адаптация к вирусу, происходящая без вакцинации. Факты адаптации к мутациям вируса гриппа указывают на то, что адекватный иммунный ответ приобретает коллективный характер, охватывая естественным путем без вмешательства медиков целую популяцию. Коллективный иммунитет формируется естественным образом путём дистанционного межклеточного взаимодействия, иными словами, так называемого биополя.
Проблема полевой (нехимической) коммуникации биологических систем возникла более ста лет назад с целью объяснения механизма реализации наследственной информации в морфогенетическом процессе (5). «Описать сложную и изменяющуюся в онтогенезе архитектонику многоклеточного организма исходя только из химических, механических и электрических взаимодействий оказалось невозможным. Необходимо было, согласно Г. Дриша, найти «инженера развития», который мог бы на базе генетической информации выстраивать пространственную структуру. В различные годы на эту роль предлагались такие обобщающие понятия, как «энтелехия» Г. Дриша, «динамически преформированная морфа» А.Г. Гурвича, «организаторы» Г. Шпемана, «физиологические градиенты» Ч.М. Чайлда, «креоды» К.Х. Уоддингтона. Несмотря на терминологические различия, во всех случаях имелось в виду существование некоторого «генерализующего начала», определяющего целостность и неразрывность развития живых организмов. Пытаясь объяснить природу явления, «…биологи не смогли придумать ничего лучшего, как постулировать наличие морфогенетического поля, которое определяет облик образуемой структуры», – пишет основатель системной биологии Конрад Уоддингтон (6).
В биологической литературе поле впервые упоминается Г. Шпеманом – “organizationsfeld” (организующее поле) в 1921 году. Дальнейшее развитие этой гипотезы принадлежит А.Г. Гурвичу, который отмечал, что «…идея поля возникает из необходимости признания пространственных взаимоотношений между молекулами, не вытекающих из их ближнедействия».
Представления о полях биологических систем были весьма популярны среди эмбриологов и морфологов в 30–50 годы прошлого века. В различных трактовках это понятие использовали Г. Шпеман, Дж. Гексли, Р. Сноу и М. Сноу и другие для описания процессов морфогенеза. Несмотря на широкое использование термина «поле», его природа и механизм регуляции оставались отвлеченными понятиями, то есть дело свелось лишь к названию, а не объяснению механизма дистанционного взаимодействия живых организмов.
Однако, это факт не может служить препятствием для изучения механизмов воздействия на морфогенетические поля. Современная наука хорошо изучила электрические процессы, но ничего не знает о природе электричества. Самый цитируемый российский физик академик Людвиг Фаддеев пишет, что об электроне мы не знаем ничего, кроме его математических характеристик.
Особое место в исследовании биологических полей занимает дистанционное межклеточное взаимодействие (ДМВ). Оно получило надежное экспериментальное подтверждение и стало основным аргументом в пользу существования полевой (нехимической) коммуникации биологических систем. Впервые такая форма межклеточного взаимодействия была обнаружена в 1923 году А.Г. Гурвичем (5).
Дистанционное межклеточное взаимодействие наблюдают у одноклеточных организмов-бактерий различного уровня организации: от прокариот до высших эукариот. Наиболее вероятно его функционирование происходит посредством низкоинтенсивных электромагнитных полей оптической области спектра. Это вытекает из многочисленных экспериментальных данных (5).
Вот схема типичного эксперимента с пыльцой растений, описанная в профильной литературе:
Дистанционное межклеточное взаимодействие моделировали на пыльце сливы домашней (Prunus domestica L.). Пыльцу наносили на поверхность предметных стекол, покрытых тонким слоем питательной среды. Выполненные таким образом препараты служили в качестве индуктора, детектора и контроля. Индуктор облучали когерентным светом гелий-неонового лазера. Инкубация проходила 24 часа при температуре 28°C. В этот период индукторы и детекторы имели между собой оптический контакт, а контрольные препараты были изолированы от них светонепроницаемыми перегородками. Далее пыльцу инактивировали хлороформом. О биологическом эффекте судили по числу проросших пыльцевых зерен, для чего просматривали по 50 независимых полей зрения у каждого типа препаратов.
Лазерное облучение в 2–4 раза повысило прорастаемость пыльцы препаратов-индукторов в сравнении с интактным контролем. Необлученная пыльца биодетекторов на большинстве препаратов также показала статистически значимые различия. Функциональная активность пыльцы обработанного лазерным излучением индуктора и необработанного детектора, наоборот, была практически одинаковой. Столь заметное совпадение функциональной активности детектора и индуктора говорит о наличии дистанционного взаимодействия, возникающего между пыльцевыми зернами при наличии оптического контакта. Такое явление наблюдали на многих (но не на всех) режимах, и оно было особенно выражено в области коротких (0,5-4 мин) длительностей облучения. Расстояние, на котором происходило взаимодействие, составляло в среднем 12,5 мм.
Полученные результаты подтвердили наличие дистанционного взаимодействия в системе двух изолированных препаратов с облученной и необлученной пыльцой. Эффект наиболее устойчиво воспроизводился при длительности лазерного облучения индуктора в течение 30 секунд. Заключался он в существенной (в 3–5 раз) и статистически значимой (α < 0,001) по отношению к контролю стимуляции функциональной активности как индукторов, так и сопряженных с ними биодетекторов (5).
Британский биолог Руперт Шелдрейк провел ряд экспериментов, в частности, на грызунах. В течение 20 лет эксперимента лабораторных крыс заставляли от поколения к поколению проходить все более сложный лабиринт. Крысы справлялись с этим все лучше, и можно было предположить, что старики учат молодых. Но оказалось, что крысы за свето- и звуконепроницаемой стенкой, которых каждый раз брали просто с улицы, тоже от раза к разу проходили лабиринт все лучше, как будто могли учиться у своих лабораторных родичей с помощью телепатии (4).
4. Управлять распространением вирусов с помощью биополей.
Достаточно странным, если не сказать больше, является тот факт, что огромное число экспериментов, доказывающих существование дистанционного межклеточного взаимодействия живых организмов любого уровня сложности – от бактерий до лабораторных крыс – не подвинули научное сообщество к изучению аналогичных процессов в наномире вирионов и вирусов, а также в сфере их симбиоза с многоклеточными организмами.
Можно предположить, что в данном случае имеет место самоцензура биологов и вирусологов, введённая под давлением «глобальной мафии» ВОЗ и корпораций Big Pharma.
Между тем, исследования дистанционного взаимодействия в мире вирусов и бактерий требуют осторожного и взвешенного подхода. Ряд биологов ставят перед собой задачу уничтожить вирусы в целом. Но это может привести к необратимым и непредсказуемым последствиям, учитывая, что сейчас вирусы играют роль своего рода «санитаров биологического леса», как бы прагматично это ни звучало.
Можно вспомнить, что когда современная медицина решила проблему бактериальных инфекций, на смену бактериям пришли вирусы, так как природа не терпит пустоты. И если мы избавим людей от вирусных атак, то что или кто придет им на смену.
Новые методы борьбы с вирусными инфекциями, в первую очередь, с коронавирусными, должны дать медикам возможность подавлять размножение любых болезнетворных вирусов в организме человека, так как точечная война с каждым новым вирусом не имеет шансов на победу.
Тем самым медицина даст человечеству стратегическое оружие противодействия организаторам инфодемий, которые хотят превратить нашу планету в свой карантинный барак, развязывая новые и новые волны пандемий.
Владимир Прохватилов,
старший научный сотрудник Академии военных наук
Читайте также: