Роль гетерогенности экосистем в экотоксикологии. Гетерогенность популяции и токсическая нагрузка
Добавил пользователь Евгений Кузнецов Обновлено: 28.01.2026
Небрежное отношение к биосфере ведет к нарушению ее правильной работы и означает, что люди не смогут вообще существовать на Земле.
Н. В. Тимофеев-Ресовски
Влияние токсических факторов на процессы воспроизводства в природных популяциях животных и растений
Воспроизводство является одним из ключевых процессов, определяющих жизнеспособность природных популяций животных и растений. Особо значимы эти процессы в нестабильных условиях, в том числе при химическом загрязнении среды обитания, которое следует рассматривать в качестве одного из экстремальных факторов антропогенного происхождения.
Процессы воспроизводства в ценопопуляциях
Меристема, а также образующиеся из нее генеративные органы, в частности мужская генеративная сфера, являются одной из наиболее чувствительных к токсическому действию промышленных загрязнителей системой растений. При изучении реакции мужского гаметофита на воздействие внешних факторов принято различать оплодотворяющую способность (фертильность) и жизнеспособность пыльцы. Фертильность предполагает способность мужского гаметофита вызывать полное оплодотворение. Жизнеспособность пыльцы рассматривается как ее способность к росту на соответствующих тканях пестика.
На процессы микроспорогенеза, прорастания пыльцы и оплодотворения у растений влияет множество абиотических факторов: неблагоприятная погода, недостаток минерального питания, воздействие высоких или низких температур, высокие дозы химических веществ, радиоактивное загрязнение. Реакция пыльцы на действие физических и химических факторов может быть различна. Рядом исследователей установлено, что микроэлементы и УФ-радиация в малых дозах могут стимулировать ростовые процессы, а при высоких воздействиях — их подавляют.
Проведенные исследования по древесным и кустарниковым растениям свидетельствуют о негативном влиянии высоких концентраций тяжелых металлов и радиации на функциональное состояние и физиологическую активность пыльцы. Следствием подобного влияния могут стать замедленное прорастание пыльцы, нарушение роста пыльцевых трубок и полная остановка их роста, потеря жизнеспособности половых клеток (Третьякова и др., 2004; Beda, 1982).
Имеются сведения о влиянии высоких концентраций тяжелых металлов и радиационного излучения на функциональное состояние и физиологическую активность пыльцы травянистых растений (Бессонова, 1992; Попова и др., 1992; Реакция мужского гаметофита. 2007; Третьякова и др., 2004; Holub et al., 1988).
Под действием химического загрязнения наблюдается уменьшение размеров пыльцевых зерен, что может рассматриваться в качестве одного из показателей адаптированности растений к техногенным условиям среды. Многие исследователи указывают на то, что под действием внешних факторов происходит образование деформированных (сморщенных, разрушенных, линзовидных, гигантских) и недоразвитых пыльцевых зерен (Бессонова, 1992; Сафонов, 1999; Чукпарова, 2001; Третьякова и др., 2004; Beda, 1982).
В условиях загрязнения среды промышленными поллютантами и тяжелыми металлами (Fe, Си, Mn, Cr, Zn и др.) отмечается резкое снижение оплодотворяющей способности пыльцы. В популяциях растений, находящихся в зонах антропогенного воздействия, повышается стерильность пыльцы, которая может достигать 50—80 %, что обусловлено редукцией клеток археспория, цитопатологией генеративной ткани или нарушением мейоза (О возможностях. 1996; Оценка последствий. 1993).
Рассмотрим особенности функционального состояния мужского гаметофита (на примере форм одуванчика лекарственного Taraxacum officinale Wigg. s.l.).
Размеры пыльцевых зерен.
Отмечено уменьшение средних размеров пыльцы с увеличением химической нагрузки (рис. 7.1). Влияние химического загрязнения на исследуемый признак статистически значимо только в случае f. pectinatiforme (rs = -0,96; df = 7; р 0,001; для f. dahlstedtii rs = -0,18; df = 7; p = 0,7). На максимально загрязненном участке у растений f. dahlstedtii пыльца крупнее, чем у f. pectinatiforme. Различия между формами по исследуемому показателю подтверждены однофакторным дисперсионным анализом (F (1; 348) = 6,82; р
Уменьшение размеров пыльцевых зерен в условиях техногенного загрязнения показано также в работах В. П. Бессоновой (1992), А. У. Чукпаровой (2001) и др. Изменение величины пыльцы связано с нарушением процесса роста и деления клеток при формировании первичных клеток археспория и при формировании тетрады микроспор (Бессонова, 1992). Уменьшение размеров пыльцы некоторыми авторами рассматривается в качестве одного из показателей адаптированное™ растений к техногенным условиям среды (Лебедева, 1993).
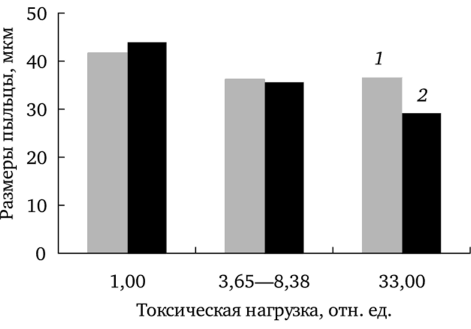
Рис. 7.1. Зависимость размеров пыльцы f. dahlstedtii (7) и f. pectinatiforme (2) от уровня химического загрязнения
Анализ вариабельности этого показателя позволил установить, что влияние химического загрязнения проявляется в элиминации особей с крупными пыльцевыми зернами (рис. 7.2). Сравнение выборочных распределений растений с разными размерами пыльцы из ценопопуля- ции фонового участка с остальными показало статистически значимые различия.
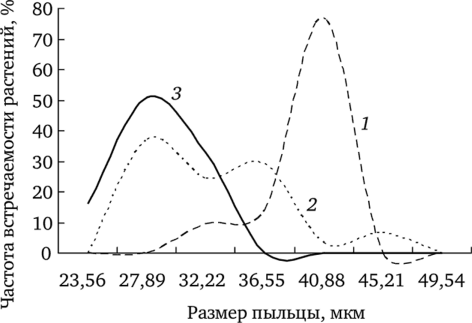
Рис. 7.2. Частота встречаемости растений с различными размерами пыльцы (по: Реакция мужского гаметофита. 2007):
1 — фоновый; 2 — буферный; 3 — импактный участки
С ростом химического загрязнения доля фертильной пыльцы снижается. Соответственно, на загрязненных участках при уменьшении фертильной пыльцы у исследуемого вида возрастает доля стерильных пыльцевых зерен.
Известно, что Т. officinale s.l. является видом с апомиктическим способом размножения. Вместе с тем высокие значения стерильности пыльцы у растений из ценопопуляций максимально загрязненных территорий по сравнению с фоновыми указывают на существенную роль химического загрязнения на процессы микроспорогенеза.
Оплодотворяющая способность пыльцы
Т. officinale s.l. в градиенте токсической нагрузки. Возникновение частичной или полной стерильности у растений часто связывают с нарушением правильности развития и строения пыльцы (Левина, 1981). В модели пошагового дискриминантного анализа показатель фертильности пыльцы обеих форм в нашем случае достоверно различается на всех участках (рис. 7.3).
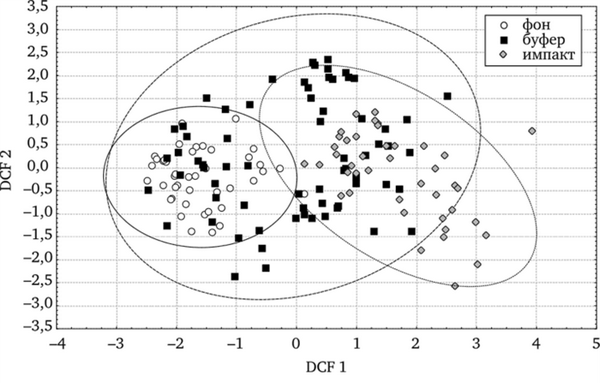
Рис. 7.3. Дискриминантный анализ фертильности пыльцы у ценопопуляций Т. officinale (по: Реакция мужского гаметофита. 2007)
Пространственная гетерогенность среды
Важнейшим обстоятельством, определяющим уровни накопления токсических веществ элементами биоты, является пространственная неоднородность территории. В реальных условиях эколого-климатические факторы проявляют свою прерывистость в пространственно-временных масштабах и формируют некоторую экологическую мозаику среды обитания природных популяций, определяя тем самым их структуру. На эту естественную мозаичность природной среды накладывается неоднородность полей загрязнения, вызванная неравномерностью воздушных потоков, особенностями рельефа местности, другими географическими и геохимическими параметрами территории. К этому добавляется пространственная неоднородность биотических механизмов, способствующих пространственной миграции и фиксации поступающих в среду токсикантов.
Таким образом, различие в содержании токсических веществ в биологических объектах, а значит, и те потоки загрязнителей, которые включаются в общий круговорот вещества в биоценозе через отдельные пространственные группировки организмов, являются отражением совместного влияния природных и техногенных факторов.
Подобная гетерогенность пространства особенно отчетливо прослеживается по отношению к загрязнению почвы, где распределение токсикантов носит наиболее сложный характер. Оно определяется не только вариабельностью их поступления, но и перераспределением в радиальных и латеральных потоках, сорбцией на почвенных минералах, поглощением почвообразующими организмами и т.д. Структура загрязнения почвы — это основная часть «матрицы», определяющей структуру токсической нагрузки на все остальные компоненты биоценоза.
Изучение варьирования в почвах тяжелых металлов было начато Н. Г. Зыриным (1968), который показал, что коэффициент вариации валового содержания тяжелых металлов в отсутствие локального загрязнения достигает 10—20 %, а для подвижных форм — 25—30 %. Причем неоднородность распределения концентраций одинакова при различных масштабах отбора проб. Эти выводы подтверждаются другими авторами (см., например, Воробейчик, 2002; Мотузова, 1992). Многие из них отмечают, что загрязнение почв наземных экосистем тяжелыми металлами и сернистым газом имеет следствием подавление процессов деструкции органического вещества почв. Если в качестве метода исследования этих процессов измерять убыль со временем в почвах воздушно-сухой массы чистой целлюлозы, то пространственное распределение данного показателя отражает общую изменчивость естественных деструкционных процессов и варьирование полей химического загрязнения. Подробные исследования этих характеристик в градиенте загрязнения почв тяжелыми металлами показали, что коэффициенты вариации скорости деструкционных процессов при различных масштабах отбора проб изменялись от 16,9 ± 2,2 % на фоновой территории до 136,9 ± 17,7 % для зон сильного загрязнения (Воробейчик, 2002).
Фитотоксичность подстилки в первую очередь определяется пространственно неоднородным распределением обменных форм металлов, что ведет к возникновению выраженной нелинейности в дозовой зависимости: при относительно небольшом перепаде уровня токсической нагрузки наблюдается максимальный разброс реакции организмов.
По мнению А. Д. Покаржевского и соавторов (2000), мозаичность распределения микроатропод или простейших в почвах отражает не только неоднородность концентраций токсических веществ, но и пространственное распределение экосистем низшего иерархического уровня.
С. В. Левин с соавторами (1989) также отмечают увеличение вариации численности почвенных микроорганизмов в условиях химического загрязнения почв до 200 %.
При оценке меры токсического влияния необходимо исходить из положения С. С. Шварца (1967), согласно которому популяция организмов представляет собой совокупность микропопуляций, связанных между собой общностью происхождения из единых популяционных стаций резервации. Поэтому интересующему нас популяционному уровню организации должен соответствовать некоторый пространственный уровень токсической нагрузки, отражающий, с одной стороны, пространственную гетерогенность загрязнения территории, с другой — возможность пространственного перемещения биологических объектов. Имеются в виду механизмы поддержания стабильности биоценозов за счет постоянного их пополнения растениями и животными с более чистых или полностью незагрязненных участков.
С этой точки зрения упомянутая выше мера токсического воздействия, определяемая по статистическому распределению токсикантов в живых организмах, составляющих изучаемую выборку, может зависеть не только от прямого загрязнения участков и гетерогенности этих уровней, но и от наличия рядом более чистых зон, откуда возможны миграционные потоки.
Следует подчеркнуть еще раз тот важный факт, что уровни токсической нагрузки зависят не только от уровня токсического загрязнения (это очевидно!), но в значительной мере контролируются рядом факторов внешней среды, а также важнейшими внутрипопуляционными процессами.
Роль гетерогенности экосистем в экотоксикологии. Гетерогенность популяции и токсическая нагрузка
Гетерогенность популяций в экотоксикологии, как фактор выживания
В условиях сильно деградированных местообитаний вблизи источника токсической эмиссии создание стабильных поселений оседлых особей затруднено, что свидетельствует о крайнем неблагополучии среды обитания для данного вида и невозможности существования животных в течение полного жизненного цикла. Население рыжей полевки в этих условиях представлено главным образом мигрирующими особями из сопредельных более благоприятных стаций, расположенных на больших расстояниях от источника загрязнения.
Методами математического моделирования показано, что площадь прямого токсического поражения, на которой резко снижается или полностью исчезает население мелких млекопитающих, зависит от конкретных условий. Если интенсивное токсическое загрязнение охватывает наиболее благоприятные стации обитания, а смежные территории слабо обеспечивают восполнение населения животных за счет мигрантов, то общая зона поражения может существенно превышать территорию интенсивного загрязнения.
Возможен и обратный вариант, когда наличие обильных, не тронутых токсическим влиянием и пригодных для обитания животных сопредельных территорий может поддерживать их численность на достаточно высоком уровне, ограничивая за счет миграционных перемещений зону токсического поражения.
Сказанное можно иллюстрировать, рассматривая две возможные ситуации. В первой в зону прямого токсического влияния попадает большинство стаций, благоприятных для укрытия и размножения животных. Во второй — на той же территории стации преимущественного обитания оказываются вне зоны интенсивного загрязнения. Можно предположить, что наряду с уменьшением количества сохранившихся стаций обитания в первом варианте снижается численность оседлых особей и возрастает доля мигрантов в общем обилии зверьков.
Согласно нашим данным, в этом случае можно ожидать большей плодовитости размножающихся самок. Площадь фактического поражения в первом варианте превышает зону прямого токсического загрязнения. Во втором — она меньше, так как благоприятные смежные территории за счет мигрирующих особей способны поддерживать численность населения животных на интенсивно загрязненных участках.
Таким образом, пространственная гетерогенность природных популяций позволяет им поддерживать жизнеспособность даже в условиях интенсивного токсического загрязнения территории. Миграционные перемещения животных при этом следует рассматривать как проявление обычных неспецифических механизмов популяционной адаптации, обусловленных состоянием среды обитания.
Изменчивость морфофизиологических параметров организмов, входящих в состав природных популяций, является важнейшей характеристикой, поскольку определяет способность последних адаптироваться к условиям внешней среды. При этом часто отмечают увеличение или снижение вариабельности этих показателей у животных в неблагоприятных условиях существования (обитание на границе ареала, химическое загрязнение и др.).
В экологической токсикологии важным является изменение вариабельности таких популяционных параметров, как плодовитость, выживаемость молодняка, времени полового созревания и т.д., связанных с процессами воспроизводства. Именно эти показатели в большей степени определяют благополучие популяции и ее способность к адаптации, в том числе к воздействию токсических факторов среды.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Токсичность на уровне популяции и биоциноза. Токсическая нагрузка в экотоксикологии
Казалось бы, дело обстоит предельно просто: достаточно знать содержание токсичных веществ в объектах внешней среды, чтобы прогнозировать их накопление в растительности, по содержанию в растительности — у животных-фитофагов и т.д., определяя тем самым токсическую нагрузку на отдельные компоненты биоты.
Однако в реальных условиях множество трудно учитываемых механизмов влияют на эти процессы. Все многообразие этих проявлений можно условно разделить на две группы действующих факторов:
• Пространственную мозаичность и различие уровней загрязненности территории, определяемых спецификой техногенного воздействия, локальными почвенно-климатическими и физико-химическими условиями среды.
• Особенности экологии растительных и животных сообществ, включающие видовую и сезонную специфику пищевых рационов, разнокачественность стаций обитания, миграционные потоки и т.д.
Влияние абиотических факторов среды. В качестве примера рассмотрим влияние кислотности среды на накопление некоторых токсичных веществ в биоте. Редко мы продавая или приобретая авто обращаем внимание на его токсичность. На сегодняшний день во многом автомобили являются источниками токсинов в биопопуляции. Как показывает исследование рынка, продажа авто не во всех регионах страны осуществляется под строгим контролем токсичности. Многие автомобили вторичного рынка имеют повышенную токсичность. Один из приоритетных загрязнителей атмосферного воздуха — выброс сернистого газа, в результате окисления которого наблюдается снижение рН дождевой воды (так называемые "кислые" дожди). Дренирование такой дождевой воды через почвенные горизонты приводит к снижению рН в почвенных водах и водоемах. Таким образом, конечное звено трансформации выбросов сернистого газа — водные экосистемы, в которых аккумулируются загрязнения значительных территорий и уже в силу этого в наибольшей степени проявляются токсические последствия. В результате подкисления вод в реках и водоемах наблюдаются эффекты, связанные не только с прямым токсическим действием на гидробионты низких рН, но и с опосредованным влиянием других факторов.
Из-за того что растворимые формы токсических элементов физиологически более активны, особое значение приобретают некоторые сопутствующие абиотические факторы, включающие процессы осаждения, гидролиза и комплексообразования, в конечном счете определяющие токсичность элементов для биоты естественных водоемов.
К таким факторам относят: адсорбцию токсических элементов на взвешенных частицах или гидроксилах железа, марганца и ряда других элементов.

Проблемы влияния этих показателей на токсичность ряда элементов широко обсуждаются в литературе. Однако до настоящего времени ученые не располагают четкой и однозначной картиной этих явлений.
Так, разнообразие реакции биоты на изменение только рН воды можно иллюстрировать данными некоторых авторов.
Показано, что возрастание рН с 0,6 до 8,0 снижает токсичность свинца для амфипод. Для радужной форели максимальная токсичность пятиокиси ванадия наблюдается при рН 7,7. При больших и меньших значениях этого показателя отмечено снижение токсичности.
На том же объекте изучали влияние токсичности меди при различной жесткости воды в диапазоне рН от 5,0 до 9,0. Влияние кислотности наиболее существенно при высокой жесткости и при значениях рН от 6,0 до 7,0. При меньших и больших значениях токсичность снижалась.
Аналогичным образом общее содержание токсичных веществ в почвах не всегда адекватно отражает меру токсического воздействия на компоненты природных популяций и биоценозы. Лишь подвижные формы химических элементов могут переходить в растительность и далее по трофической цепочке. По этой причине рекомендуемые в настоящее время ориентировочные допустимые концентрации некоторых тяжелых металлов и мышьяка могут отличаться в 3—5 раз в зависимости от типа почв.
Отметим, что, кроме чисто химических взаимодействий, в изменении метаболических свойств и токсичности ряда элементов может участвовать микробиота почв и водоемов. Так, большое значение имеет комплекс бактерий почв и донных отложений, который в анаэробных и аэробных условиях может способствовать изменению химических форм ряда токсических элементов и изменению связанной с этим их токсичности.
Примером может служить ртуть. Мы имеем в виду интенсивные процессы биологического метилирования, которые, по мнению многих авторов, протекают наиболее интенсивно в поверхностных слоях донных отложений. Известно, что интенсивность этих процессов пропорциональна величине рН водной среды. В этих условиях накопление метил- и диметилртути в придонной фауне и планктоне, а вместе с этим и у животных следующих трофических уровней также зависит от кислотности водоема. Это хорошо иллюстрируют данные И.К.Степановой и В.Г.Комова, установивших прямую зависимость накопления ртути в мышцах окуня в зависимости от рН воды озер северо-запада России.
Таким образом, в реальных природных экосистемах любые загрязняющие вещества являются источниками возникновения сложной смеси компонент, биологическое накопление которых биотой и их токсичность не всегда можно предвидеть.
В реальных условиях эколого-климатические факторы проявляют свою прерывистость во многих пространственно-временных масштабах и формируют некоторую экологическую мозаику среды обитания природных популяций, определяя тем самым их структуру. На эту естественную мозаичность природной среды накладывается неоднородность полей загрязнения, вызванная неравномерностью воздушных потоков, особенностями рельефа местности и другими географическими параметрами территории. Таким образом, различия в содержании токсичных веществ в биологических объектах, а значит, и те потоки загрязнителей, которые включаются в общий круговорот вещества в биоценозах через отдельные пространственные группировки организмов, являются отражением сочетанного влияния природных и техногенных факторов.
Сегодня принято считать, что любая популяция организмов представляет собой совокупность микропопуляций, связанных друг с другом общностью происхождения из единых популяционных стаций резервации. Поэтому интересующих нас популяционному или биоценотическому уровням биологических систем должны соответствовать некоторые пространственно-временные группировки изучаемых организмов. При этом характер пространственной структуры систем популяционного ранга определяется как наивыгоднейшим для них использованием ресурса стаций обитания в целом, так и мерой токсического воздействия на них.
В проблеме токсической деградации территории принципиальным является возможность пространственного перемещения биологических объектов. Имеется в виду возможность поддержания "пораженных" популяций и биогеоценозов за счет постоянного их пополнения растениями и животными с более чистых или полностью незагрязненных участков.
При этом восполнение фитоценозов ограничивается возможностью пространственного рассеивания семян или вегетативных побегов. Популяции животных, как правило, пространственно более мобильны, постоянный приток особей с чистых территорий может существенно "разбавить" население загрязненных участков. Таким образом, мера токсического воздействия, определяемая по содержанию токсикантов в организмах животных, может зависеть не только от прямого загрязнения участка, но и от наличия рядом более чистых стаций обитания, откуда возможен постоянный миграционный приток животных.

Как отмечали выше, токсическая нагрузка на животных, обладающих широкой кормовой территорией, так же как их миграционные перемещения, в определенной мере интегрирует пространственную неоднородность токсического загрязнения среды. Однако объем и состав рационов характеризуют не только среду обитания, но отражают энергетические потребности отдельных популяций и субпопуляционных групп организмов. Последнее обстоятельство часто обусловливает различие уровней токсикантов, накапливаемых отдельными видами и внутрипопуляционными группами.
Многочисленные данные показывают возрастные особенности накопления токсичных веществ. Так, у мелких млекопитающих максимальные уровни отмечены у перезимовавших, наиболее старых зверьков; меньшие уровни—у половозрелых и неполовозрелых сеголеток.
Отмечают половые различия в накоплении токсичных веществ. Чаще всего более высокие уровни отмечают у самцов. Это может быть обусловлено необходимостью иметь более высокий энергетический потенциал в связи с их ролью в поддержании активной иерархической структуры популяции. Эти и другие особенности экологии самцов и самок, обитающих в природных условиях (суточная активность зверьков, размеры индивидуальных участков, участие в размножении и т.д.), вероятно, могут быть сведены к энергетическим затратам зверьков и как следствие этого к количеству потребляемого корма и поступающих с ним токсичных веществ.
Отметим еще раз важную закономерность, согласно которой максимальная дифференциация животных (видовая, возрастная, половая) по накоплению ими токсичных веществ проявляется в наибольшей степени по мере возрастания загрязненности среды обитания.
Читайте также: