Рост клеток. Дифференцировка клеток.
Добавил пользователь Евгений Кузнецов Обновлено: 09.01.2026
Клеточный, или жизненный, цикл клетки — это время существования клетки от деления до следующего деления,или от деления до смерти. Для разных типов клеток клеточный цикл различен.
В организме млекопитающих и человека различают следующие три группы клеток, локализующиеся в разных тканях и органах: часто делящиеся клетки (малодифференцированные клетки эпителия кишечника, базальные клетки эпидермиса и другие);
редко делящиеся клетки (клетки печени — гепатоциты);
неделящиеся клетки (нервные клетки центральной нервной системы, меланоциты и другие).
Жизненный цикл у этих клеточных типов различен.
Жизненный цикл у часто делящихся клеток — это время их существования от начала деления до следующего деления. Жизненный цикл таких клеток нередко называют митотическим циклом. Такой клеточный цикл подразделяется на два основных периода: митоз или период деления;
интерфаза — промежуток жизни клетки между двумя делениями.
Варианты перехода в следующие стадии клеточного цикла:
1. Клетка вступает в митотический цикл, обязательным условием которого является репликация ДНК. Начинается синтетический период интерфазы.
2. Клетка прекращает рост и переходит в фазу дифференцировки и нормальной активности. Эту стадию иногда называют стадией пролиферативного покоя (G0). Однако слово покой не означает переход клетки в состояние анабиоза. В клетке активно осуществляются метаболические процессы, начинаются процессы дифференцировки клетки.
Дифференцировка – это процесс формирования морфологических особенностей клеток, обеспечивающих выполнение специфических функций. Процессы дифференцировки клеток наблюдаются на всех этапах онтогенеза, а сам процесс обусловлен избирательной активностью определенных генов. Другими словами, характерные морфологические и функциональные особенности клеток определяются активностью только частью генов из всего генотипа.
По степени специализации клетки можно разделить на недифференцированные и дифференцированные. Но только дифференцированные клетки могут полноценно выполнять свои функции. Поэтому любое нарушение дифференцировки приводит к нарушению или не выполнению функций (опухолевые клетки эндокринных органов, появление в крови незрелых лимфоцитов).
Нормальное функционирование в составе органа – это основной период жизненного цикла клеток. Продолжительность этого периода различна для каждого типа клеток. Но неизбежным завершением ЖЦК является переход в фазу старения и гибели.
Старение и гибель клеток. После определенного периода нормального функционирования у клетки начинается период старения, который морфологически проявляется:
- уменьшением объема клетки
- увеличением содержания крупных лизосом
- накоплением пигментных и жировых включений
- появлением вакуолей в цитоплазме и ядре
Гибель клетки – завершающий этап клеточного цикла. Гибель клетки – эволюционно обоснованный (как механизм клеточного гомеостаза и условие нормальной жизнедеятельности тканей) и генетически закрепленный процесс. У соматических клеток имеется запрограммированный предел возможных делений. В последнее время активно изучается особый участок хромосом - теломера, содержащий ген «бессмертия». Как полагают ученые, активность гена определяет количество последовательных митозов, но это количество у нормальных клеток ограничено. У опухолевых клеток функция гена нарушена, и они могут делиться неограниченное число раз.
При гибели клетки можно выделить два различных механизма ее развития: некроз и апоптоз.
Некроз – возникает под действием резко выраженных повреждающих факторов (температурных, гипоксия, химические и механические воздействия, и т.д.). Другими словами, некроз – «смерть в результате несчастного случая». На начальном этапе наблюдаются изменения органоидов клетки (набухание митохондрий и уменьшение в них крист, распад цистерн платинчатого комплекса), нарушения проницаемости плазмолеммы, повреждение мембран лизосом и выделение гидролаз. Наблюдаются изменения и ядра клетки – кариопикноз, кариорексис, кариолизис. Остаточные продукты распада клеток привлекают лейкоциты и макрофаги, вокруг очага некроза возникает воспалительная реакция (признаки воспаления – отек, гиперемия, боль).
Апоптоз – активный генетически контролируемый процесс гибели клетки. Апоптоз энерноемкий процесс и регулируется различными внутренними факторами клетки. Он обычно происходит в отдельных клетках и имеет место в норме у человека как в эмбриональной периоде так и у взрослого. Особенно значима роль апоптоза в следующих процессах: - формировании органов в ходе эмбрионального развития
- удаление стареющих клеток в зрелых тканях
- реакциях тканей на действие повреждающих факторов
- в развитии инфекционных заболеваний
Сигналы, запускающие генетическую программу апоптоза:
1. Нарушение регуляторных механизмов, поддерживающих нормальную дифференцировку и функциональную активность клеток, потеря контакта с другими клетками, изменения морфологии и функций органоидов при старении клеток.
2. Возникновение неустранимых повреждений ДНК.
3. Инфекционные заболевания (вирусные).
Стадии апоптоза: - утрата клеткой контакта с соседними клетками
- сжатие цитоплазмы и ядра клетки, образование вздутий и выпячиваний мембраны, кариопикноз
- распад клетки на фрагменты и их фагоцитоз соседними клетками, в отличие от некроза воспалительная реакция отсутствует.
Дифферон (гистогенетический ряд) – совокупность клеточных форм, составляющих ту или иную линию
дифференцировки. Исходными клетками дифферона являются стволовые клетки, далее идут несколько переходных этапов - полустволовые, молодые (бластные) и созревающие клетки, и наконец зрелые или дифференцированные клетки. Различают полный дифферон - когда в ткани содержатся клетки всех этапов развития (например, эритроцитарный дифферон в красном костном мозге или эпидермальный дифферон в эпидермисе кожи) и неполный дифферон - когда в тканях содержатся только переходные и зрелые или даже только зрелые формы клеток (например, нейроциты центральной нервной системы). В диффероне последовательно различают: стволовые клетки, клетки-предшественницы, зрелые клетки, достигшие состояния окончательной (терминальной) дифференцировки.
Стволовые клетки - самоподдерживающаяся популяция клеток, способных дифференцироваться в нескольких
направлениях и формировать различные клеточные типы.
Стволовые клетки обладают высокими пролиферативными потенциями, но, как правило, делятся редко.
Клетки-предшественницы. По мере дифференцировки их пролиферативные потенции постепенно уменьшаются.
Выделяют наиболее раннюю стадию клеток- предшественниц - коммитированные, или полустволовые,
Зрелые клетки. Ими заканчивается гистогенетический ряд. Способность к пролиферации полностью исчезает.
Дифферон (гистогенетический ряд) - совокупность клеточных форм, составляющих ту или иную линию дифференцировки. В диффероне различают: стволовые клетки - клетки-предшественницы - зрелые клетки, достигшие состояния окончательной (терминальной) дифференцировки.
• Стволовые клетки - самоподдерживающаяся популяция клеток, способных дифференцироваться в нескольких направлениях и формировать различные клеточные типы. Так, стволовые эпендимные клетки ЦНС дают начало разным нейронам и глиоцитам. Стволовые клетки обладают высокими пролиферативными потенциями, но, как правило, делятся редко.
• Клетки-предшественницы. По мере дифференцировки пролиферативные потенции клеток постепенно уменьшаются. Выделяют наиболее раннюю стадию клеток-предшественниц - коммитированные, или полустволовые клетки.
• Зрелые клетки. Ими заканчивается гистогенетический ряд.
Стволовыми клетками эмбриона, плода и взрослого организма считаются клетки, которые при определённых условиях способны длительное время воспроизводить себе подобные клетки и в течение жизни давать начало специализированным клеткам, образующих ткани и органы тела. Выделяют тотипотентные и плюрипотентные стволовые клетки.
♦ Тотипотентная клетка (лат. totus - целый, полный) обладает потенциалом давать начало всем специализированным клеткам, формирующим ткани эмбриона. Например, зигота и бластомеры по всем признакам относятся к тотипотентным клеткам.
♦ Плюрипотентные клетки (лат. plures - несколько, много) дифференцируются в разные полипотентные клетки всех трёх зародышевых листков - экто-, энто- и мезодермы. Клетки внутренней клеточной массы бластоцисты относятся к плюрипотентным клеткам.
• Стволовые клетки эмбриона. У эмбриона выделяют (т.е. получают in vitro) плюрипотентные эмбриональные стволовые клетки и эмбриональные половые клетки.
• Стволовые клетки взрослого организма выделены из красного костного мозга, периферической крови, пульпы зуба, спинного и головного мозга, кровеносных сосудов, скелетной мышцы, эпителия кожи и пищеварительной системы, роговицы и сетчатки глаза, печени и поджелудочной железы. Это полипотентные клетки, потомки которых дают начало ограниченному количеству типов унипотентных клеток-предшественниц
Рост клеток. Дифференцировка клеток.
Дифференциация клеток. Детерминация клеток. Клеточная гибель. Понятие о диффероне.
Дифференциация — это стойкое структурно-функциональное преобразование клеток в различные специализированные клетки. Дифференцировка клеток биохимически связана с синтезом специфических белков, а цитологически — с образованием специальных органелл и включений. При дифференцировке клеток происходит избирательная активация генов. Важным показателем клеточной дифференцировки является сдвиг ядерно-цитоплазменного отношения в сторону преобладания размеров цитоплазмы над размером ядра. Дифференцировка происходит на всех этапах онтогенеза. Особенно отчетливо выражены процессы дифференциации клеток на этапе развития тканей из материала эмбриональных зачатков. Специализация клеток обусловлена их детерминацией.
Детерминация — это процесс определения пути, направления, программы развития материала эмбриональных зачатков с образованием специализированных тканей. Детерминация может быть оотипической (программирующей развитие из яйцеклетки и зиготы организма в целом), зачатковой (программирующей развитие органов или систем, возникающих из эмбриональных зачатков), тканевой (программирующей развитие данной специализированной ткани) и клеточной (программирующей дифференцировку конкретных клеток). Различают детерминацию: 1) лабильную, неустойчивую, обратимую и 2) стабильную, устойчивую и необратимую. При детерминации тканевых клеток происходит стойкое закрепление их свойств, вследствие чего ткани теряют способность к взаимному превращению (метаплазии). Механизм детерминации связан со стойкими изменениями процессов репрессии (блокирования) и экспрессии (деблокирования) различных генов.
Клеточная гибель — широко распространенное явление как в эмбриогенезе, так и в эмбриональном гистогенезе. Как правило, в развитии зародыша и тканей гибель клеток протекает по типу апоптоза. Примерами программированной гибели являются гибель эпителиоцитов в межпальцевых промежутках, гибель клеток по краю срастающихся небных перегородок. Программированная гибель клеток хвоста происходит при метаморфозе личинки лягушки. Это примеры морфогенетической гибели. В эмбриональном гистогенезе также наблюдается гибель клеток, например при развитии нервной ткани, скелетной мышечной ткани и др. Это примеры гистогенетической гибели. В дефинитивном организме путем апоптоза погибают лимфоциты при их селекции в тимусе, клетки оболочек фолликулов яичников в процессе их отбора для овуляции и др.
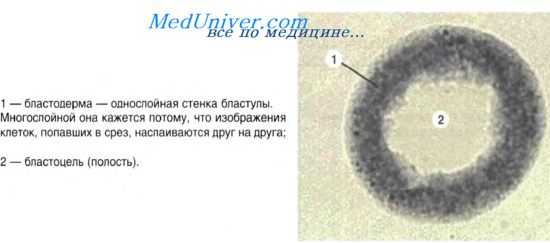
Понятие о диффероне. По мере развития тканей из материала эмбриональных зачатков возникает клеточное сообщество, в котором выделяются клетки различной степени зрелости. Совокупность клеточных форм, составляющих линию дифференцировки, называют диффероном, или гистогенетическим рядом. Дифферон составляют несколько групп клеток: 1) стволовые клетки, 2) клетки-предшественники, 3) зрелые дифференцированные клетки, 4) стареющие и отмирающие клетки. Стволовые клетки — исходные клетки гистогенетического ряда — это самоподдерживающаяся популяция клеток, способных дифференцироваться в различных направлениях. Обладая высокими пролиферативными потенциями, сами они (тем не менее) делятся очень редко.
Клетки-предшественники (полустволовые, камбиальные) составляют следующую часть гистогенетического ряда. Эти клетки претерпевают несколько циклов деления, пополняя клеточную совокупность новыми элементами, и часть из них затем начинают специфическую дифференцировку (под влиянием факторов микроокружения). Это популяция коммитированных клеток, способная дифференцироваться в определенном направлении.
Зрелые функционирующие и стареющие клетки завершают гистогенетический ряд, или дифферон. Соотношение клеток различной степени зрелости в дифферонах зрелых тканей организма неодинаково и зависит от основных закономерных процессов физиологической регенерации, присущих конкретному виду ткани. Так, в обновляющихся тканях обнаруживаются все части клеточного дифферона — от стволовой до высокодифференцированной и гибнущей. В типе растущих тканей преобладают процессы роста. Одновременно в ткани присутствуют клетки средней и конечной частей дифферона. В гистогенезе митотическая активность клеток постепенно снижается до низкой или крайне низкой, наличие стволовых клеток подразумевается только в составе эмбриональных зачатков. Потомки стволовых клеток некоторое время существуют как пролиферативный пул ткани, но их популяция быстро расходуется в постнатальном онтогенезе. В стабильном типе тканей имеются лишь клетки высокодифференцированной и гибнущей частей дифферона, стволовые клетки обнаруживаются лишь в составе эмбриональных зачатков и полностью расходуются в эмбриогенезе.
Изучение тканей с позиций их клеточно-дифферонного состава позволяет различать монодифферонные — (например, хрящевая, плотная оформленная соединительная и др.) и полидифферонные (например, эпидермис, кровь, рыхлая волокнистая соединительная, костная) ткани. Следовательно, несмотря на то, что в эмбриональном гистогенезе ткани закладываются как монодифферонные, в дальнейшем большинство дефинитивных тканей формируются как системы взаимодействующих клеток (клеточных дифферонов), источником развития которых являются стволовые клетки разных эмбриональных зачатков.
Ткань — это фило- и онтогенетически сложившаяся система клеточных дифферонов и их неклеточных производных, функции и регенераторная способность которой определяется гистогенетическими свойствами ведущего клеточного дифферона.
Ткань является структурным компонентом органа и в то же время частью одной из четырех тканевых систем — покровных, тканей внутренней среды, мышечных и невральных.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Эмбриональный гистогенез. Пролиферация клеток. Клеточный рост, миграция и межклеточные взаимодействия.
В развитии низших и высших позвоночных отчетливо прослеживается единая общебиологическая закономерность, выражающаяся в появлении зародышевых листков и обособлении основных зачатков органов и тканей. Процесс образования тканей из материала эмбриональных зачатков составляет суть учения о гистогенезах.
Эмбриональный гистогенез, по определению А.А. Клишова (1984), — это комплекс координированных во времени и пространстве процессов пролиферации, клеточного роста, миграции, межклеточных взаимодействий, дифференциации, детерминации, программированной гибели клеток и некоторых других. Все названные процессы в той или иной мере протекают в зародыше, начиная с самых ранних стадий его развития.
Пролиферация. Основной способ деления тканевых клеток — это митоз. По мере увеличения числа клеток возникают клеточные группы, или популяции, объединенные общностью локализации в составе зародышевых листков (эмбриональных зачатков) и обладающие сходными гистогенетическими потенциями. Клеточный цикл регулируется многочисленными вне- и внутриклеточными механизмами. К внеклеточным относятся влияния на клетку цитокинов, факторов роста, гормональных и нейрогенных стимулов. Роль внутриклеточных регуляторов играют специфические белки цитоплазмы. В течение каждого клеточного цикла существуют несколько критических точек, соответствующих переходу клетки из одного периода цикла в другой. При нарушении внутренней системы контроля клетка под влиянием собственных факторов регуляции элиминируется апоптозом, либо на некоторое время задерживается в одном из периодов цикла.
Метод радиографического анализа клеточных циклов в различных тканях выявил особенности соотношения клеточной репродукции и дифференцировки. Например, если в тканях (кроветворные ткани, эпидермис) имеется постоянный фонд пролиферирующих клеток, за счет которых обеспечивается непрерывное возникновение новых клеток взамен погибающих, то эти ткани относятся к обновляющимся. Другие ткани, например, некоторые соединительные, характеризуются тем, что в них увеличение количества клеток происходит параллельно с их дифференцировкой, клетки в этих тканях характеризуются низкой митотической активностью. Это растущие ткани. Наконец, нервная ткань характеризуется тем, что все основные процессы репродукции заканчиваются в период эмбрионального гистогенеза (когда формируется основной запас стволовых клеток, достаточный для последующего развития ткани). Поэтому она отнесена к стабильным (стационарным) тканям. Продолжительность жизни клеток в обновляющихся, растущих и стабильных тканях разная.
Наряду с обновлением клеточной популяции, в самих клетках постоянно наблюдается обновление внутриклеточных структур (внутриклеточная физиологическая регенерация).
Клеточный рост, миграция и межклеточные взаимодействия. Рост клеток проявляется в изменении их размеров и формы. При усилении функциональной активности и внутриклеточных биосинтезов наблюдается увеличение объема клетки. Если объем клетки превышает некую норму, то говорят о ее гипертрофии, и наоборот, при снижении функциональной активности происходит уменьшение объема клетки, а при переходе некоторых нормативных параметров возникает атрофия клетки. Рост клетки не беспределен и определяется оптимальным ядерно-цитоплазменным отношением.
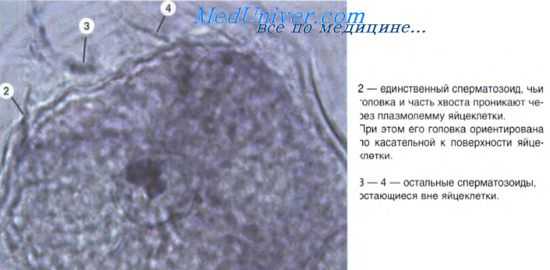
Важное значение для гистогенеза имеют процессы перемещения клеток. Миграция клеток наиболее характерна для периода гаструляции. Однако и в период гисто- и органогенеза происходят перемещения клеточных масс (например, смещения миобластов из миотомов в места закладки скелетных мышц; движение клеток из нервного гребня с образованием спинномозговых ганглиев и нервных сплетений, миграция гоноцитов и т. д.). Миграция осуществляется с помощью нескольких механизмов. Так, различают хемотаксис — движение клеток в направлении градиента концентрации какого-либо химического агента (перемещения спермиев к яйцеклетке, предшественников Т-лимфоцитов из костного мозга в закладку тимуса).
Гаптотаксис — механизм перемещения клеток по градиенту концентрации адгезионной молекулы (движение клеток протока пронефроса у амфибий по градиенту щелочной фосфатазы на поверхности мезодермы). Контактное ориентирование — когда в какой-либо преграде остается один канал для перемещения (описан у рыб при образовании плавников).
Контактное ингибирование — этот способ перемещения наблюдается у клеток нервного гребня. Суть способа заключается в том, что при образовании ламеллоподии одной клеткой и контакта ее с другой клеткой, ламеллоподия прекращает рост и постепенно исчезает, но в другой части мигрирующей клетки при этом формируется новая ламеллоподия.
В процессе миграции клеток важную роль играют межклеточные взаимодействия. Существует несколько механизмов такого взаимодействия (контактного и дистантного). Выделяется большая группа молекул клеточной адгезии (МКА). Так, кадгерины — это Са2+-зависимые МКА, отвечают за межклеточные контакты при образовании тканей, за формообразование и др. В молекуле кадгерина различают внеклеточный, трансмембранный и внутриклеточный домены. Например, внеклеточный домен ответственен за адгезию клеток с одинаковыми кадгеринами, а внутриклеточный — за форму клетки. Другой класс МКА — это иммуноглобулиновое суперсемейство Са2+-независимых МКА, обеспечивающих, например, адгезию аксонов к сарколемме мышечных волокон, или миграцию нейробластов вдоль радиальных глиоцитов в закладке коры большого мозга и др. Следующий класс МКА — это мембранные ферменты — гликозилтранферазы. Последние по типу "ключ-замок" соединяются с углеводными субстратами — гликозаминогликанами надмембранного комплекса клетки, осуществляя таким образом прочное сцепление клеток.
Кроме механизмов межклеточного взаимодействия, существуют механизмы взаимодействия клеток с субстратом. Они включают формирование рецепторов клетки к молекулам внеклеточного матрикса. К последним относят производные клеток, среди которых наиболее изученными адгезионными молекулами являются коллаген, фибронектин, ламинин, тенасцин и некоторые другие. Коллагены, среди которых различают несколько десятков типов, входят в состав межклеточного вещества рыхлой волокнистой соединительной ткани, базальной мембраны и пр. Фибронектин, секретируемый клетками, является связывающей молекулой между мигрирующей клеткой и межклеточным матриксом. Ламинин — компонент базальной мембраны, также связывает мигрирующие клетки с межклеточным матриксом (справедливо по отношению к эпителиоцитам и нейробластам).
Для осуществления связи мигрирующих клеток с межклеточным матриксом клетки формируют специфические рецепторы. К ним относятся, например, синдекан, который обеспечивает контакт эпителиоцита с базальной мембраной за счет сцепления с молекулами фибронектина и коллагена. Интегрины клеточных поверхностей связывают с внеклеточной стороны молекулы внеклеточного матрикса, а внутри клетки — белки цитоскелета (например, актиновые микрофиламенты). Так возникает связь внутри- и внеклеточных структур, что позволяет клетке использовать для перемещения собственный сократительный аппарат. Наконец, существует большая группа молекул, формирующих клеточные контакты, осуществляющие коммуникацию между клетками (щелевые контакты), механическую связь (десмосомы, плотные контакты).
Дистантные межклеточные взаимодействия осуществляются путем секреции гормонов и факторов роста (ФР). Последние — это вещества, оказывающие стимулирующее влияние на пролиферацию и дифференцировку клеток и тканей. К ним относятся, например, ФР, полученный из тромбоцитов и влияющий на переход клеток в фазу размножения (гладких миоцитов, фибробластов, глиоцитов); эпидермальный ФР — стимулирует пролиферацию эпителиоцитов, производных эктодермы; ФР фибробластов — стимулирует пролиферацию фибробластов. Особо выделяется большая группа пептидов (соматотропины, соматомедины, инсулин, лактоген), влияющих на развитие клеток плода.
Факторы, тормозящие пролиферацию и дифференцировку клеток, также принимают кооперативное участие в процессах развития клеток и тканей. К ним относятся, например, бета-интерферон и трансформирующий ФР. Последний, однако, в отношении разных клеточных типов действует по-разному: блокирует размножение многих типов эпителиоцитов, но стимулирует размножение соединительнотканных клеток.
- Вернуться в оглавление раздела "гистология"
Синтетическая деятельность клеток проявляется в изменении их величины. При усилении синтеза наблюдается увеличение клеточных размеров и, наоборот, при снижении синтетической активности может происходить их уменьшение. Все клетки в той или иной степени способны к росту. Однако их рост ограничен определенными рамками. Некоторые клетки, например яйцевые, благодаря накоплению в них желтка, могут достигать огромной величины. Значительно превышают обычные размеры гигантские клетки, образующиеся особенно часто в опухолевых тканях. Вместе с тем, для клеток некоторых типов (например, для клеток крови) характерна стабильность средних размеров.
Обычно клеточный рост сопровождается преимущественным увеличением объема цитоплазмы, тогда как величина ядра изменяется в меньшей степени. На протяжении онтогенеза строение клеток непрерывно изменяется. Прогрессирующие изменения клеток, связанные с появлением морфологических и функциональных различий между ранее индифферентными (однородными) клетками и обусловленные их специализацией в процессе развития, называют диф-ференцировкой клеток. Биохимической основой этого процесса является синтез специфических белков и других веществ. Молекулярные основы синтеза белков складываются из транскрипции первичной структуры матричной РНК на основе информации ДНК-гена, кодирующей области генов — экзонные области; процессинга мРНК, в результате которого из новообразованной цепи удаляются несмысловые последовательности нуклеотидов (интроны), перехода новообразованной мРНК в цитоплазму и трансляции — синтеза белка на аппарате синтеза белков клетки.

Дифференцировка сопровождается качественными, количественными и временными параметрами, т. е. характеризуется изменениями клеточной структуры, темпом развития (ускоренная или замедленная) и степенью (малодифференцированные — высокодифференцированные клетки). Усложнение структуры клетки сопровождается следующими изменениями: приобретением определенной формы и размеров ядра и клетки; сдвигом ядерно-цитоплазменного отношения в связи с более значительным ростом цитоплазмы по сравнению с ядром; развитием орга-нелл; образованием специализированных клеточных структур; синтезом специфических включений; образованием межклеточного вещества; появлением межклеточных взаимодействий и установлением межклеточных и специализированных контактов. Так, эпителиальные клетки приобретают кубическую, призматическую или плоскую форму. Клетки тканей внутренней среды более разнообразны по форме.
Некоторые соединительнотканные клетки вырабатывают межклеточное вещество. Мышечные клетки содержат миофибриллы. Между нейронами формируются синаптические контакты (подробно — см. соответствующие главы).
Для разных тканевых клеток характерны определенные взаимоотношения между процессами дифференцировки и деления. Однако в целом по мере повышения степени дифференцировки способность клеток к делению закономерно уменьшается.
В гистогенезе клетки определенного цитотипа интегрируются, частично теряя автономность, присущую ранней стадии (пролиферативной), вследствие формирования регуляторных механизмов, оказывающих влияние на цитодифференцировку.
Клетки в составе тканей многоклеточных организмов имеют различную продолжительность жизни. В некоторых тканях, например в эпителиях, смена клеточного состава происходит довольно быстро. По данным ряда исследователей, жизнь эпителиальных клеток (в частности, в кишечном эпителии) измеряется несколькими сутками. В других тканях (например, в мышечной и нервной) продолжительность жизни клеток сравнима с продолжительностью жизни организма. Так или иначе, клеточный состав организма не остается постоянным, он непрерывно изменяется в результате гибели части клеток и замены их новыми клеточными поколениями. В нормальных физиологических условиях гибели клеток предшествуют процессы старения. Они сопровождаются снижением функциональной активности, а также появлением ряда дистрофических изменений. В общих чертах гибель клеток характеризуется сморщиванием ядра и клетки в целом, диффузной окрашиваемостью и исчезновением специализированных структур, распадом клетки на фрагменты, которые фагоцитируются соседними клетками или макрофагами.
РОСТ И РАЗВИТИЕ
РОСТ И РАЗВИТИЕ. С тех пор, как в ходе эволюции возникли многоклеточные организмы, превращение оплодотворенного яйца во взрослую особь совершается в каждом поколении в процессе роста и развития. Рост, т.е. увеличение размеров, достигается за счет повышения количества таких субъединиц, как молекулы и клетки. Развитие, т.е. качественное изменение, обеспечивается синтезом новых соединений и образованием клеток разных типов в результате дифференцировки.
Процессам роста и развития присущи определенные физические ограничения, удерживающие увеличение размеров и изменения формы в известных пределах. С увеличением линейных размеров вдвое площадь поверхности увеличивается в 4 раза, а объем в 8 раз. Это имеет важнейшее значение для таких параметров, как регуляция температуры и прочность структуры, необходимой для поддержания возрастающей массы организма. Хотя клетки бывают самых разных размеров – от крошечного сперматозоида до огромного яйца страуса – их размеры тем не менее ограничиваются теми расстояниями, которые могут быстро преодолеть питательные вещества и продукты распада, диффундируя в цитоплазме. Некоторые из самых крупных клеток нашего тела – нервные и мышечные – справляются с этими ограничениями, сочетая увеличение длины с сильным сокращением диаметра. С другой стороны, уменьшение размеров клеток тоже не может быть безграничным: необходим некий минимальный объем, где могли бы разместиться все разнообразные внутриклеточные структуры.
Рост и развитие традиционно воспринимаются как процессы, идущие по нарастающей (со знаком «плюс»); на самом же деле они могут идти и со знаком «минус». Поэтому в общем смысле рост представляет собой изменение, а не «приращение». Фундаментальное свойство роста – обновление, т.е. утрата отдельных частей и добавление новых. При росте с положительным знаком процессы синтеза идут активнее, чем процессы распада. При старении преобладает обратное соотношение. На протяжении большей части жизни взрослого организма синтез и распад сбалансированы. Можно сказать, что в состоянии равновесия организм в каждый данный момент чуть-чуть умирает и чуть-чуть возрождается. Время полужизни содержащихся в организме веществ измеряется периодами от нескольких минут до нескольких месяцев. В состоянии постоянного обновления находятся все органеллы клетки. Продолжительность жизни клеток многих типов ограниченна, а это означает, что их число остается постоянным только потому, что образуются новые клетки данного типа. Обновление возможно даже на тканевом уровне – например, в яичниках созревают новые фолликулы для замещения утраченных в предыдущем менструальном цикле.
РОСТ КЛЕТОК
Все живое состоит из клеток. Поскольку клетки не могут быть крупнее некоторых максимальных размеров, рост организма возможен только за счет увеличения числа клеток. Последнее достигается с помощью митоза – клеточного деления, при котором сначала на две части делится ядро, а затем цитоплазма.
Каждая из двух клеток, образовавшихся в результате митоза, вдвое меньше исходной. Поэтому прежде чем приступить к следующему делению, клетки должны пройти период роста, в ходе которого у них удваивается число органелл и пополняется количество цитоплазмы. Лишь после восстановления нормальных размеров клетки готовы к следующего делению. См. также КЛЕТКА.
Форма и размеры клеток зависят от их функции. Тело человека построено из клеток нескольких сот разных типов, которые по их способности к делению можно разбить на три категории. Наивысшей митотической активностью обладают клетки обновляющихся тканей, названных так потому, что они постоянно обновляются на клеточном уровне. Например, эпидермальные клетки делятся, находясь в базальном слое кожи; затем по мере продвижения к поверхности кожи они дифференцируются, а оказавшись на поверхности, отмирают и слущиваются, прожив лишь несколько недель. Эпителиальные клетки, выстилающие пищеварительный тракт, иногда живут всего несколько дней, после чего отмирают и выводятся с фекальными массами. Сперматозоидам, яйцеклеткам и клеткам крови уготована та же судьба: они рождаются, стареют и гибнут, и процесс замены их новыми клетками повторяется многократно.
Клетки второй категории способны к митозу, но потенциально могут существовать до тех пор, пока жив организм в целом. Такие клетки составляют т.н. разрастающиеся ткани: они растут только в период роста всего тела, а после того, как организм достигает окончательных размеров, митотическая активность прекращается. Разрастающиеся ткани образуют многие внутренние органы – печень, почки и железы, как эндо-, так и экзокринные.
К третьей категории относятся клетки, которые по окончании ранних стадий развития совершенно утрачивают способность к делению. Примерами могут служить клетки таких тканей, как нервная и мышечная. Хотя эти клетки могут оставаться живыми до тех пор, пока жив организм, они настолько высокоспециализированы, что митоз для них невозможен. Именно поэтому сердце и головной мозг не способны к регенерации. Их клетки могут увеличиваться в размерах, но не в числе, и эти органы, во всяком случае у высших животных, расходуют в процессе развития весь запас эмбриональных клеток, которые могли бы обеспечить в дальнейшем восстановление поврежденной ткани. У низших позвоночных животных – рыб и хвостатых амфибий – сохраняется достаточное количество недифференцированных клеток, чтобы обеспечить регенерацию некоторых частей как головного и спинного мозга, так и сердца. Среди тритонов есть виды, способные регенерировать даже хрусталик и сетчатку глаза после полного иссечения этих структур.
РАСТЕНИЯ
В семенах растений имеется эндосперм, снабжающий зародыш питательными веществами подобно тому, как желток обеспечивает питание развивающемуся зародышу животных. Семена сосудистых растений при прорастании образуют корни и побеги.
Несмотря на значительные различия между корнями и побегами, у них много общего. И те и другие многократно ветвятся, а их растущие кончики, состоящие из недифференцированных клеток, образуют конусы нарастания (верхушечные меристемы). Многократные митотические деления в конусе нарастания постоянно поставляют новые клетки, обеспечивающие рост в длину. Непосредственно за этой зоной пролиферации находятся зоны дифференцировки и растяжения; здесь новообразованные клетки превращаются в специализированные клетки ксилемы и флоэмы – проводящих тканей растения. В процессе дифференцировки эти клетки сильно растягиваются в длину, что обеспечивает очень быстрый рост побегов (например, у бамбука). Между ксилемой и флоэмой расположен слой камбиальных клеток, за счет которых происходит утолщение стеблей и корней.
Приведенное выше описание относится в основном к деревьям и кустарникам. В отличие от них, у многих травянистых растений зона нарастания листьев находится у основания, а не на верхушке. Листья растут у них снизу, и именно поэтому газон приходится подстригать многократно. Деревья и живые изгороди тоже подстригают, чтобы придать им определенную форму, однако при этом их зоны нарастания срезаются. В результате после обрезки ветвей кусты и деревья растут гуще, потому что при повреждении верхушки побега меристемы, отдаленные от его кончика, принимают на себя функции утраченной части. До удаления верхушечной меристемы, оказывавшей на них тормозящее воздействие, эти латеральные меристемы пребывали в латентном состоянии; освободившись от торможения, они дают начало боковым ветвям.
Это явление иллюстрирует механизм, регулирующий рост растения. Верхушечная меристема вырабатывает гормональные вещества (ауксины), которые, перемещаясь вниз по стеблю, тормозят рост других меристем. Ауксины определяют также тропизмы растений, например тенденцию расти в сторону источника света. Инактивируясь на освещенной стороне стебля, они стимулируют удлинение стебля на теневой стороне, заставляя его склоняться в направлении к источнику света.
От света зависят также сроки вегетации: каждый вид растений начинает и заканчивает рост, цветет и производит семена в определенное время года. В умеренных широтах жизненные циклы растений приспособлены к колебаниям температуры и к удлинению или укорочению светового дня. Некоторым видам для цветения необходим длинный, а другим короткий день. Там, где колебания температуры и длины светового дня минимальны, прежде всего в тропиках, в координации жизненных циклов растений может участвовать чередование периодов дождей и засухи.
Однолетние растения запрограммированы на прекращение роста и отмирание в первый (и единственный) год своей жизни, а продолжение существования вида обеспечивается семенами. В отличие от них многолетние растения, в частности деревья, обладают способностью к потенциально неограниченному росту. За счет верхушечных меристем всех побегов объем тканей ежегодно увеличивается, а за счет камбия происходит рост ствола в толщину и повышается его прочность. Способность деревьев расти до тех пор, пока они живут, а жить до тех пор, пока они растут, демонстрирует пример секвойи с ее гигантскими размерами и потенциальным бессмертием.
Жизнь многолетников удается продлить с помощью вегетативного размножения. У отводков можно вызвать образование корней (иногда при помощи гормонов) и вырастить из них новые растения, обладающие теми же генетическими признаками, что и родительское растение. См. также ГОРМОНЫ РАСТЕНИЙ.
ЖИВОТНЫЕ
В отличие от растений, рост которых происходит путем удлинения и разрастания в стороны, большинство развивающихся животных растут за счет увеличения размеров каждого органа или ткани. Головной мозг растет вначале быстро, но по мере того, как его клетки прекращают деление и только увеличиваются в размерах, его рост замедляется. Рост и развитие половых органов происходит в основном в период полового созревания. Хотя каждый орган следует своему собственному «расписанию», существует также механизм общего контроля, регулирующий конечные размеры тела животного. У позвоночных эту роль выполняет в основном гормон роста, вырабатываемый гипофизом. Под действием гормона роста происходит в первую очередь удлинение костей, каждая из которых прекращает рост в длину на определенной стадии развития. Связанные с костями ткани (мышцы, нервы, кровеносные сосуды, кожа) перестают расти, когда кривая роста животного достигает плато.
Описанный механизм роста свойствен животным с детерминированным, или ограниченным, ростом, в первую очередь – наземным животным: их размеры не могут перейти некий предел, за которым утрачивается способность поддерживать массу тела. У многих водных животных, напротив, рост продолжается неопределенно долго даже после наступления половой зрелости, и они достигают очень крупных размеров. Это объясняется тем, что в водной среде животные находятся как бы в состоянии невесомости и им не приходится поддерживать свое тело, а потому в процессе эволюции у них не возник механизм ограничения роста. В этом отношении рост рыб сходен с ростом многолетних растений.
Рост рыб на протяжении всей жизни происходит за счет увеличения числа функциональных единиц в их органах и тканях, т.е. в структурах, клетки которых у более высоко организованных животных перестают делиться на относительно ранней стадии жизни. Так, у рыб по мере роста добавляются новые клетки в головном мозге и новые палочки и колбочки в сетчатке глаз; возможна также дифференцировка дополнительных мышечных волокон в сердечной и скелетных мышцах. Кости у рыб растут за счет отложения на их поверхности нового материала. По мере увеличения челюстей на них вырастают как совершенно новые зубы, так и замещающие утраченные. Чешуи увеличиваются в результате добавления новых колец, а плавники удлиняются за счет формирования дополнительных сегментов на кончиках их костных лучей.
Многие животные в процессе развития претерпевают метаморфоз. При этом они получают возможность использовать на разных стадиях жизни разные местообитания и разную пищу. Например, у чешуекрылых личиночная стадия представлена листоядными гусеницами, а взрослая – бабочками, которые питаются нектаром, перелетая с цветка на цветок. На стадии куколки личиночные ткани постепенно разрушаются, а из скоплений недифференцированных клеток – т.н. имагинальных дисков – развиваются крылья и ноги. У лягушек из икры вылупляются растительноядные головастики, которые вначале обитают в воде, а затем превращаются в наземных плотоядных животных, дышащих воздухом. Хвосты и жабры головастиков резорбируются, а взамен развиваются ноги и легкие.
У некоторых животных свойственная зародышу способность к развитию сохраняется во взрослом состоянии, обеспечивая регенерацию утраченных частей тела.
ПРОЦЕСС РОСТА У ЧЕЛОВЕКА
Рост в высоту каждого человека предопределен его генами, о чем свидетельствуют расовые различия, например между пигмеями и бурунди. У высоких родителей дети обычно бывают тоже высокими, а дети тучных родителей предрасположены к полноте. Однако характер телосложения зависит также от питания и гормональных воздействий. Современный человек несколько выше ростом, чем были его предки, жившие несколько веков назад; это отчасти можно объяснить улучшением питания и здравоохранения, а отчасти – проявлением «гибридной мощности», создающейся в результате смешения генофондов при браках между людьми разных национальностей или рас.
Гормон роста способствует росту в детском и юношеском возрасте, но с наступлением зрелости его влияние ослабевает. Избыток гормона роста приводит к гигантизму, а его недостаточность – к карликовости.
Неудивительно, что питание оказывает глубокое влияние на рост, особенно в раннем возрасте. Плохое питание в период развития плода может вызвать нарушения пролиферации клеток в развивающемся головном мозге и привести к умственной отсталости. Дети, которые недоедают, растут медленнее тех, кто питается нормально, но если вовремя перевести их на достаточное питание, они догоняют по росту своих однолеток и, став взрослыми, мало или совсем не отличаются по росту от других людей.
На рост в утробе матери оказывают также влияние условия в матке, причем немалое значение имеет ограниченность пространства. У близнецов масса при рождении обычно бывает меньше, чем у ребенка, родившегося в результате одноплодной беременности, а у троен – меньше, чем у двоен. В таких случаях последующий ускоренный рост может, в конечном счете, сгладить прежнее отставание.
Читайте также:
- Спленэктомия, лейкаферез при хроническом лимфолейкозе - результаты
- Клиника и диагностика венесуэльского энцефаломиелита. Лечение венесуэльского энцефаломиелита
- Лечение опухоли желточного мешка и ее прогноз
- Роль натрия и калия в регуляции бронхиальной проводимости.
- Ретракционный карман барабанной перепонки и его лечение