Строение хромосом: хроматин, центромера, теломера
Добавил пользователь Евгений Кузнецов Обновлено: 27.01.2026
Часто залогом научного открытия является сочетание качеств учёного с удачным выбором объекта исследования. Перед вами авторы открытия роли хромосом в наследственности.
Томас Хант Морган (1866-1945) - выдающийся биолог, один из основателей генетики, лауреат Нобелевской премии в области физиологии и медицины (1933).
Дрозофила фруктовая (Drosophila melanogaster) (продолжительность жизни при 25 °С - около 14 дней) - насекомое ряда Двукрылые, которая является незаменимым модельным объектом генетических исследований. У неё всего 4 пары хромосом (2n = 8).
СОДЕРЖАНИЕ
Какова роль хромосом в наследственности?
ХРОМОСОМЫ (от греч. хроматос - окрашенный, сома - тельце) - структуры клеток эукариотов, обеспечивающие сохранение, распределение и передачу наследственной информации. Чаще других первооткрывателем хромосом называют немецкого анатома В. Флемминга (1843-1905). Именно он применил анилиновые красители для обработки ядерных структур. Термин «хромосомы» в 1888 г. ввёл в научный оборот Г. Вальдейер (1836-1921). Эти исследования положили начало цитогенетеке - разделу, изучающему роль хромосом в наследственности.
Хромосомы формируются из ядерного материала - хроматина перед делением клеток. В их составе основная доля приходится на ДНК и ядерные белки. Нуклеотиды ДНК кодируют наследственную информацию, а ядерные белки-гистоны организуют компактное пространственное расположение молекул ДНК для хранения этой информации. Перед делением клетки однохроматидные хромосомы удваиваются, уплотняются и становятся двухроматидными. Происходят эти процессы для распределения и передачи наследственной информации следующему поколению клеток.
Количество хромосом в клетках эукариотов различных видов различно и не зависит от уровня организации, а также не всегда указывает на филогенетическое родство (табл. 5).
Таблица 5. ЧИСЛО ХРОМОСОМ У НЕКОТОРЫХ ВИДОВ ЭУКАРИОТОВ
Организм
Количество
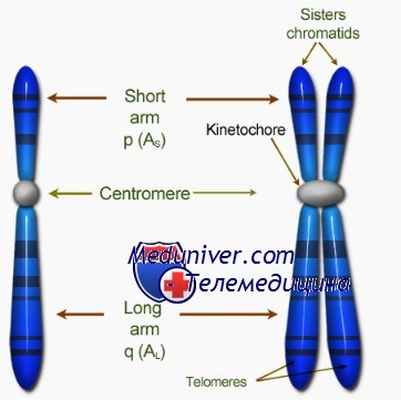
Ил. 58. Строение хромосомы: 1 - однохроматидная хромосома; 2 - двухроматидная хромосома; 3 - центромера; 4 - теломеры; 5 - сестринские хроматиды; 6 - нити веретена деления; 7 - кинетохоры
Основные функции хромосом взаимосвязаны со строением. Для сохранения, распределения и передачи наследственной информации в хромосомах имеются такие элементы строения, как сайты начала репликации, центромеры и теломеры. Сайты начала репликации - последовательность нуклеотидов, определяющая начало самоудвоения молекулы ДНК.
Центромера - часть хромосомы, разделяющая её на два плеча. На ней располагаются белки-кинетохоры, к которым при распределении генетического материала прикрепляются нити веретена деления. Теломеры - участки на концах хромосом, препятствующие их соединению между собой и восстановлению с помощью фермента теломеразы утраченных после разделения участков (ил. 58).
Итак, хромосомы являются материальными носителями наследственности всех эукариотических организмов, и их функции тесно взаимосвязаны с химическим составом и строением.
Как происходят спирализация и конденсация хромосом?
В каждой хромосоме содержится только одна молекула ДНК, которая достигает нескольких сантиметров. Так, у человека длина ДНК крупнейшей хромосомы - около 7 см, а общая длина всех 46 молекул ДНК в одном ядре клетки человека составляет около 2 м. Каким образом хромосомы помещаются в микроскопическом ядре, размером 5 мкм? Это достигается за счёт спирализации и конденсации (от лат. сondensatio - уплотнение) хромосом, обеспечивающих многоуровневую организацию хроматина (ил. 59). Возможности такой организации впечатляют. Так, у человека длина раскрученной ДНК малейшей хромосомы составляет около 14 000 мкм, а ее длина после уплотнения - только 2 мкм. Какие же есть уровни организации хромосом?
Ил. 59. Уровни организации хромосом: 1, 2, 3 - нуклеосомный; 4 - нуклеомерный; 5 - хромомерный; 6 - хроматидный; 7 - хромосомный
Уровень уплотнения нити ДНК достигает 40 раз. Хромомерный уровень связан уже не со спирализацией, а с образованием поперечных петель вдоль хромосомы при конденсации. Общий уровень уплотнения достигает уже 1 000 раз, а диаметр такой структуры - около 300 нм. Хроматидный уровень возникает в связи с последующим заключением структуры низшего уровня иерархии и образованием хроматиды диаметром 700 нм. Хромосомный уровень характерен для метафазной хромосомы, имеющей диаметр 1 400 нм.
Итак, организация хромосом имеет многоуровневый иерархический порядок, благодаря которому уровень общего уплотнения достигает 7 000 - 10 000 раз.
Каковы особенности организации набора хромосом?
Кариотип - совокупность признаков хромосомного набора (количество хромосом, форма, размеры). Каждому виду организмов присущ определённый кариотип (ил. 60). Основными правилами организации кариотипа являются:
Ил. 60. Кариотип дрозофилы фруктовой (самка - 6А + ХХ, самец - 6А + XY, где А - аутосомы, Х, Y - половые хромосомы)
• правило специфичности - особенности кариотипа особей вида зависят от количества, размеров и формы хромосом;
• правило стабильности - каждый вид эукариотических организмов имеет определённое и постоянное количество хромосом (например, у дрозофилы - 8 хромосом, у человека - 46);
• правило чётности - в диплоидном наборе (2n) каждая хромосома имеет себе пару, сходную по размерам и форме. Такие хромосомы называются гомологичными;
• правило индивидуальности - каждая пара гомологичных хромосом имеет свои особенности. Так, при сравнении хромосомных наборов мужских и женских особей одного вида наблюдается различие в одной паре хромосом. Эта пара получила название половых хромосом. Остальные пары гомологичных пар хромосом, одинаковые у обоих полов, имеют общее название аутосомы;
• правило преемственности (непрерывности) - в последующих поколениях клеток одного вида сохраняется не только постоянное число хромосом, но и их индивидуальные особенности. Хромосомный набор бывает диплоидным, гаплоидным, полиплоидным. Гаплоидный набор - это половинный набор, в котором все хромосомы отличаются друг от друга строением (его условно обозначают 1n). Диплоидный набор - это парный набор, в котором каждая хромосома имеет парную хромосому, сходную по строению и размерам (2n). Полиплоидный набор - это набор хромосом, кратный гаплоидному (3n, 4n, 5n т. д.).
Итак, клетки каждого вида эукариотов имеют свой особый набор хромосом - кариотип.
ДЕЯТЕЛЬНОСТЬ
Кроссворд «Хромосомы»
Получите в выделенных клетках название белка, который принимает участие в конденсации хромосом на первых её этапах.
1. Наука об особенностях хромосом.
2. Хромосомный набор.
3. Хромосомы, сходные у особей противоположного пола.
4. Первичная перетяжка хромосом.
5. Концевые участки хромосом.
6. Белки центромер, к которым прикрепляются нити веретена деления.
Задание на применение знаний
Рассмотрите иллюстрацию кариотипа человека, укажите особенности его организации (количество, форма, размеры, различия по полу). Сделайте сокращённую запись кариотипов женщины и мужчины.
ОТНОШЕНИЕ
Биология + Наука
Американские учёные Э. Блэкберн, К. Грейдер и Д. Шостак выяснили, каким образом хромосомы сохраняют свою целостность при делении клетки. Причина этого - в концах хромосом, известных как теломеры. За эти исследования учёные стали лауреатами Нобелевской премии 2009 г. в области физиологии и медицины. Какое значение для человека имеют эти и другие знания организации хромосом и кариотипа?
РЕЗУЛЬТАТ
Оценка
Задания для самоконтроля
1. Что такое хромосомы? 2. Назовите функции хромосом. 3. Что такое конденсация хромосом? 4. Назовите уровни организации хромосом. 5. Что такое кариотип? 6. Назовите правила организации кариотипа.
7. Какова роль хромосом в наследственности? 8. Как происходит конденсация хромосом? 9. Каковы особенности организации набора хромосом?
10. Какое значение для человека имеют знания организации хромосом и кариотипа?
Строение хромосом: хроматин, центромера, теломера
Строение хромосом: хроматин, центромера, теломера
Слово «хромосома» означает «окрашенное тело» (окрашивается более эффективно, чем остальные компоненты клетки). Каждая хромосома состоит из очень длинной молекулы ДНК, связанной с РНК и белками, образующей вещество, известное как хроматин. Во время интерфазы клеточного цикла (когда клетка не делится) хромосомы распределены по всему ядру, в то время как при митозе и мейозе они уплотняются, располагаясь ближе друг к другу.
ДНК содержится именно в хромосомах, вероятно, потому, что это облегчает разделение полного набора генов между дочерними клетками на стадии митоза, а также её упаковку в головки сперматозоидов после мейоза.
Способность к окрашиванию используют в диагностических целях для визуального исследования хромосом, их идентификации и выявления патологий. Различают слабоокрашенные части (эухроматин) и области с более выраженной окраской (гетерохроматин).
Генетическая информация (геном) заключена в молекуле ДНК и кодирована определённой последовательностью азотистых оснований в ДНК. Основная часть этой информации находится в составе хромосом в ядре клетки, однако небольшое количество ДНК в виде свободных цепочек локализовано в митохондриях цитоплазмы. Ядра присутствуют практически во всех клетках организма человека, за исключением эритроцитов и клеток хрусталика.
Обычно в каждом ядре содержится двухметровая цепь ДНК, разделённая между 23 парами хромосом (примерно по 4 см на хромосому). Однако непосредственно перед делением она уплотняется до 5 um (0,005 мм) посредством сложных процессов спирализации и уплотнения.
Структура хроматина хромосом
В каждой хромосоме цепь ДНК дважды обмотана вокруг октамерного комплекса, состоящего из восьми «белков хранения» (гистонов), формирующих нуклеосомы, при этом образованная структура напоминает «бусы на нити». Кора (сердцевина) нуклеосомы состоит из двух молекул каждого из четырех гистонов— Н2А, Н2В, НЗ и Н4. Гистоны заряжены положительно, а потому могут образовывать ионные связи с отрицательно заряженными фосфатными группами ДНК.
Последовательность аминокислот в гистонах у всех видов совпадает практически на 100%, указывая на огромную значимость данных элементов в поддержании структуры и функций хроматина. Каждая нуклеосома вмещает порядка 200 пар оснований молекулы ДНК и укорачивает длину цепи ДНК на одну десятую.
Затем подобная бусам нить спирализуется в соленоид, или зигзагообразную спираль (фибрилла длиной 30 нм), состоящую из 5—6 нуклеосом. При этом её структура поддерживается одной молекулой гистона HI на каждую нуклео-сому. Коэффициент упаковки при образовании соленоида примерно равен 5, что в свою очередь даёт общий коэффициент упаковки 50. Существует предположение, что в таком состоянии эухроматин находится во время интерфазы в местах, где гены не экспрессируются.
Во время митоза и мейоза хромосомы ещё больше уплотняются, при этом коэффициент упаковки по отношению к предыдущим уровням составляет 100, а общий коэффициент может достигать 5000. Считают, что фибриллы хроматина складываются в вытянутые петли, в основании которых расположен скаффолд (остов) из негистоновых хромосомных белков, которые прикрепляются к определённым последовательностям оснований, рассредоточенным вдоль молекулы ДНК. Сжатие этих негистоновых хромосомных белков предположительно обусловливает уплотнение хромосом.
Один из ключевых белков скаффолда — топоизомераза II, фермент, который расщепляет двойные цепи ДНК, переносит их через разрыв и вновь соединяет, что в свою очередь необходимо для релаксации суперспиралей ДНК во время репликации или транскрипции. Топоизомераза II прикрепляется к участкам связывания со скаффолдом, которые богаты А- и Т-основаниями (более 65% оснований составляют А и Т, см. главу 4). Считают, что каждая петля выступает в роли независимого функционального домена во время репликации или транскрипции ДНК.
На следующем этапе петли фибрилл хроматина спирализуются, в результате чего возникает наиболее уплотнённый гетерохроматин, входящий в состав хромосомы во время деления клеток.
Дифференциальное окрашивание хромосом
Наиболее уплотнённые части хромосом хорошо окрашиваются красителем Гимзы (G-окраска). При этом отчётливо видны сильно уплотнённые маленькие петли (G-полосы), так как участки связывания со скаффолдом тесно прилегают друг к другу. Их репликация происходит во время S-периода, они не активны во время транскрипции. Полосы, которые плохо окрашиваются раствором Гимзы, или R-полосы, состоят из более свободных петель, богатых Г- и Ц-основаниями, они отличаются высокой активностью во время транскрипции. В основе идентификации хромосом лежат различия в их окраске.
Центромера хромосом
Видимые на ранних стадиях митоза хромосомы состоят из двух абсолютно одинаковых структур, называемых сестринскими хроматидами, которые соединяет первичная перетяжка. Она представляет собой недуплицированный участок ДНК, или центромеру, которая дуплицируется в начале анафазы митоза.
Кинетохор — органелла, которая локализована на каждой стороне каждой центромеры в период ранней профазы митоза и облегчает полимеризацию димеров тубулина, необходимую для образования микротрубочек митотического веретена (веретена деления).
Тепомера хромосом
Теломерой называют специализированный участок конца хромосомы. К ней прикрепляются теломерспецифические белки, образующие «шапочку» (cap) для защиты конца хромосомы.
Теломеры предположительно препятствуют патологическому слиянию концов хромосом конец в конец, обеспечивают полноту репликации, участвуют в образовании пар хромосом во время мейоза и помогают восстанавливать внутреннюю структуру ядра на стадии интерфазы путём присоединения хромосом к ядерной мембране.
Эухроматин и гетерохроматин хромосом
Эухроматин находится в уплотнённом состоянии во время деления клеток и распаковывается на стадии интерфазы. Он обусловливает бледное окрашивание R-полос в уплотнённых хромосомах и содержит большинство структурных генов.
Гетерохроматин очень сильно уплотнён во время деления клетки и остаётся в таком состоянии даже на стадии интерфазы. Он локализован в основном на периферии ядра и около ядрышка и не активен во время транскрипции. Конструктивный (структурный) гетерохроматин одинаков во всех клетках организма, в то время как факультативный гетерохроматин различен в зависимости от типа генов, экспрессируемых в дифференцированных клетках определённых тканей.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Строение хромосом: хроматин, центромера, теломера
Это предположение подтверждается и проявлением центромерной активности за пределами нормальных центромер. Такие неоцентромеры ведут себя как обычные центромеры: образуют цитологически различимую перетяжку и формируют кинетохор, связывающий белки. Однако анализ ДНК двух неоцентромер человека и обычной центромеры общих последовательностей не выявил, что говорит о возможной роли других структурных компонентов хромосомы. Ими могут быть гистоновые и негистоновые белки, которые связываются с ДНК, формируя нуклеосомную структуру хроматина.
Функциональную роль центромерной структуры хроматина подтверждает присутствие специфических для каждого биологического вида варианта гистона Н3 в центромерном хроматине: у человека они названы CENP-A, у растений - CENH3. Среди множества имеющихся в кинетохоре белков только два, СЕNН3 и центромерный белок С (СЕNР-С), непосредственно связываются с ДНК. Возможно, именно CENH3, взаимодействуя с другими гистонами (Н2А, Н2В и Н4), формирует и определяет специфический для центромер тип нуклеосом. Такие нуклеосомы могут служить своего рода якорями для образования кинетохора. Варианты гистона Н3 в центромерах различных видов подобны канонической молекуле гистона Н3 в участках взаимодействия с другими гистоновыми белками (Н2А, Н2В, Н4). Однако участок центромерного гистона Н3, взаимодействующий с молекулой ДНК, видимо, находится под действием движущего отбора. Как уже говорилось, первичная структура центромерной ДНК отличается между видами, и было высказано предположение, что центромерный гистон Н3 коэволюционирует вместе с центромерной ДНК, в частности у дрозофилы и арабидопсиса [1].
Обнаружение центромерного гистона Н3 породило крайнюю точку зрения, согласно которой центромерная функция и ее полная независимость от первичной структуры ДНК определяется нуклеосомной организацией и этим гистоном. Но достаточно ли этих факторов для полноценной активности центромеры? Модели, игнорирующие роль первичной структуры ДНК, должны предполагать случайное распределение изменений в структуре центромерной ДНК в различных популяциях в отсутствие отбора. Однако анализ сателлитной ДНК в центромерах человека и Arabidopsis выявил консервативные районы, так же как и районы с более высокой, чем средняя, вариабильностью, что указывает на давление отбора на центромерную ДНК. Кроме того, искусственные центромеры удалось получить только с a-сателлитными повторами человека, амплифицированными из природных центромер, но не из a-сателлитов прицентромерных районов хромосом.
Меньше принципиальных трудностей для объяснения встречают модели, в которых решающим фактором в определении позиции центромеры (сохраняющейся от поколения к поколению) и ее функций служит третичная (или даже более высокого порядка) структура ДНК. Ее консерватизм допускает большие вариации в последовательности нуклеотидов и не исключает тонкую подстройку первичной структуры.
Хеникофф с коллегами [2] предложили модель, описывающую координированную эволюцию ДНК и белков и приводящую к появлению оптимально функционирующих центромер на примере деления женских половых клеток. Как известно, в процессе мейоза одна родительская клетка посредством следующих друг за другом двух делений дает начало четырем дочерним клеткам. Впоследствии только одна из них превращается в зрелую женскую половую клетку (гамету), передающую генетическую информацию следующему поколению, тогда как три других клетки отмирают. Согласно этой модели, в процессе эволюции вследствие мутаций и других механизмов в хромосомах могут возникать центромеры с более протяженными тяжами мономеров сателлитной ДНК или с первичной структурой нуклеотидов, более способствующей связыванию и координированной работе со специфическими формами гистонов CENH3 и СЕNР-С. При этом у одних организмов (арабидопсис, дрозофила) доказательства для положительного давления отбора получены для CENH3, тогда как для других видов (злаки, млекопитающие) для СЕNР-С (рис.4,а). В итоге такие центромеры с усовершенствованным кинетохором становятся “сильнее” и могут присоединять большее число микротрубочек веретена деления (рис.4,б). Если таких “сильных” центромер оказывается в гаметах больше, то происходит процесс мейотического драйва, который увеличивает количество таких центромер, и новый вариант фиксируется в популяции.
Рис. 4. Модель, объясняющая эволюцию центромер.
Вверху - центромеры (серые овалы) содержат специализированный набор белков (кинетохор), включающий гистоны CENH3 (H) и CENP-C (C), которые в свою очередь взаимодействуют с микротрубочками веретена деления (красные линии). В различных таксонах один из этих белков эволюционирует адаптивно и согласованно с дивергенцией первичной структуры ДНК центромер.Внизу - изменения в первичной структуре или организации центромерной ДНК (темно-серый овал) может создавать более сильные центромеры, что выражается в большем количестве присоединяемых микротрубочек.
Понять механизмы формирования и активности центромерных районов хромосом помогает сравнительная геномика. Уникальный пример разнообразной структуры центромер - хромосома 8 в геноме риса. В ней наряду с сателлитным повтором ДНК и ретротранспозонами обнаружены активно транскрибируемые гены; 48 из них имели последовательности с высокой гомологией к известным белкам [3]. Эти находки опровергают сложившееся на основе изучения центромер человека, дрозофилы и арабидопсиса мнение, что в центромерах нет активно работающих генов.
Если в молекулярной структуре центромер различных видов эукариот присутствуют некоторые универсальные характеристики (организация ДНК в виде тандемных, относительно коротких мономеров и специфические для данных локусов белки хроматина), то в размерах этих районов трудно выявить какие-либо закономерности. Так, у дрожжей Saccharomyces cerevisiae за минимальную функциональную центромеру принимают участок ДНК в 125 нп, а у дрожжей Schizosaccharomyces pombe она значительно сложнее и длиннее (от 40 до 120 тыс. нп), имеет несколько уровней организации. У человека основной компонент центромер хромосом - a -сателлитная ДНК - образует длинные тяжи тандемно организованных мономеров (от 250 тыс. до 4 млн нп). Среди 12 хромосом риса в хромосоме 8 длина тяжа с сателлитом CentO наименьшая (~64 тыс. нп); в ней определили позицию центромеры и ее примерный размер в 2 млн нп [3]. Удалось получить полную последовательность ДНК этого центромерного района и внутри него определить участок (~750 тыс. нп), где непосредственно формируется кинетохор. В этом районе находится основной кластер CentO.
Удивительная пластичность центромер, в частности активно работающие гены, обнаруженные в центромере хромосомы 8 риса, предполагает отсутствие строгой границы между центромерой и остальной частью хромосомы и даже возможность рассеянной структуры центромерного хроматина. Однако против существования нескольких кластеров в районе хромосомной перетяжки говорят недавно опубликованные данные о наличии хроматинового барьера между собственно центромерой и прицентромерным гетерохроматином у дрожжей Schizosaccharomyces pombe [4]. Барьер представляет собой ген тРНК аланина. Делеция или модификация барьерной последовательности ведет к выходу прицентромерного гетерохроматина за свои обычные границы. Более того, отсутствие барьера вызывает ненормальное расхождение хромосом в мейозе. Безусловно, следует помнить, что эти интереснейшие результаты касаются пока только одного вида дрожжей.
Термин “теломера” предложил Г.Мёллер еще в 1932 г. [5]. В его представлении она означала не только физический конец хромосомы, но и присутствие “терминального гена со специальной функцией запечатывания (пломбирования) хромосомы”, которое делало ее недоступной для вредных воздействий (хромосомных перестроек, делеций, действия нуклеаз и т.д.). Наличие терминального гена не подтвердилось в последующих исследованиях, однако функция теломеры была определена точно.
Позднее выявили еще одну функцию. Так как на концах хромосом обычный механизм репликации не работает, в клетке есть другой путь, поддерживающий стабильные размеры хромосом при клеточном делении. Эту роль выполняет специальный фермент, теломераза, которая действует подобно другому ферменту, обратной транскриптазе: использует одноцепочечную РНК-матрицу для синтеза второй цепи и восстановления концов хромосом. Таким образом, теломеры во всех организмах выполняют две важные задачи: защищают концы хромосом и поддерживают их длину и целостность.
Первые работы по определению природы ДНК теломер выявили тандемную организацию коротких мономеров у широкого спектра организмов (простейших, грибов, насекомых, растений и млекопитающих [6]), что вполне соответствовало универсальному характеру функций теломеры. Еще одна консервативная особенность теломерной ДНК - наличие относительно короткого одноцепочечного “хвоста”, состоящего из G-остатков с ориентацией 5’-3’ G-богатой цепи вперед к концу хромосомы. Считают, что такой выступ обеспечивает связывание теломер-специфических белков, образующих “колпак” (cap) для защиты конца хромосомы.
Однако по мере расширения изучаемых видов оказалось, что существуют альтернативные пути удлинения концов хромосом и их защита не зависит от короткого канонического повтора. Например, у Drosophila melanogaster ДНК теломер состоят из тандемных тяжей, образовавшихся в результате последовательных транспозиций ретротранспозонов, а одноцепочечных G-выступов пока не обнаружено. Несмотря на столь существенные различия в природе ДНК двух типов теломер, они имеют много общего. Например, и те, и другие поддерживают свою длину с помощью обратной транскрипции с РНК-матрицы и могут использовать для этой цели рекомбинацию (обмен генетическим материалом).
Список организмов, теломеры которых не имеют консенсусной последовательности, продолжает расширяться и сегодня включает некоторые виды из четырех отрядов насекомых: Diptera, Coleoptera, Heteroptera и Dermaptera . У комаров ( Diptera ) отмечен третий тип теломер: их ДНК представлена длинным тяжем регулярного тандемного повтора длиной 340 нп. Вероятно, здесь размеры поддерживаются подобно уже описанному механизму для первых двух типов, т.е. регенерация сложных повторов может происходить с помощью рекомбинации или опять-таки обратной транскрипцией РНК, продуцируемой с теломеры. Среди растений также описаны виды родов луковых и алое, не имеющих консенсусного теломерного повтора, TTTAGGG. Таким образом, подобно рассмотренным выше центромерам, теломеры выполняют исключительно важные и консервативные функции и имеют удивительно пластичную структурную организацию ДНК.
Согласно многим исследованиям, размер теломеры может колебаться при стрессовых воздействиях, укорачиваться при старении и онкогенезе. Вместе с тем другие работы демонстрируют относительное постоянство размеров теломеры у каждого конкретного вида, что указывает на функционирование регуляторного механизма, контролирующего теломеразу так, чтобы ее активность ограничивалась только компенсацией репликационных потерь теломерной ДНК. Следует отметить, что все приведенные количественные оценки носят относительный характер. Несмотря на это, можно определенно утверждать, что размеры теломер и центромер как среди различных видов эукариот, даже эволюционно близких, так и между различными хромосомами одного кариотипа характеризуются высокой гетерогенностью. В связи с этим возникает вопрос, а существует ли четкая, обусловленная различиями в молекулярной структуре граница, отделяющая эти районы от остальной хромосомы?
Анализ имеющихся данных показывает, что в случае теломер о существовании такой границы говорить трудно, если вообще возможно. Неясно, что считать настоящей (истинной) теломерой. Наиболее популярная точка зрения рассматривает в качестве теломеры весь тяж последовательности ДНК теломерного повтора вместе с многочисленными белками, связывающимися как с одно-, так и с двухцепочечной ДНК. Однако у многих видов эукариот (особенно если геномы большого размера) переход между тяжом теломерного повтора и субтеломерой характеризуется появлением мономеров с вырожденной структурой классического повтора и заканчивается копиями дегенеративных повторов с более чем одной нуклеотидной заменой. Более того, сам тяж теломерного повтора представляет собой отнюдь не такую гомогенную структуру, как было принято думать.
Мы с помощью флуоресцентной in situ гибридизации теломерного повтора на фибриллах ДНК показали, что наряду с гомогенными флуоресцирующими треками сигнала гибридизации присутствуют треки с разрывами, в которых, вероятно, помимо теломерного повтора находятся другие типы последовательностей ДНК (рис.5). Мы также обнаружили фибриллы с рассеянными одиночными сигналами теломерного повтора. Таким образом, теломерные повторы не всегда организованы как монотонные гомогенные тяжи мономеров. Они могут прерываться другими последовательностями ДНК и рассеиваться в виде коротких кластеров. Такая гетерогенная организация может приводить к завышенным количественным оценкам длины теломер и, кроме того, к неадекватной оценке участия соседних последовательностей в вариации размеров теломер.
Рис. 5. Гибридизация пробы ДНК на растянутых фибриллах ржи.
Различные типы организации теломерного повтора: монотонные треки ( вверху )
и треки с разрывами (спейсерами) различных размеров ( в середине и внизу ).
Теломерные повторы могут располагаться вдоль плеч в интерстициальных и даже в прицентромерных районах хромосом, но сами по себе не образуют функциональную теломеру. Вполне логично думать, что для ее формирования необходимы специализированные белки, обволакивающие теломерную ДНК и защищающие концы хромосомы от нежелательных воздействий. Теломерную ДНК можно рассматривать как платформу для сборки больших комплексов белков, ключевой среди которых - комплекс теломеразы. Кроме того, в реализации ее функций участвуют другие системы: ДНК-белковый комплекс, формирующийся на одноцепочечном G-выступе, и белковые комплексы на двухцепочечной ДНК теломерного повтора. Некоторые белки специфически локализованы на теломерах, но основная их часть присутствует и в других участках хромосом.
Предложена модель белкового комплекса из шести теломер-специфических белков, формирующегося на теломерах хромосом человека [7]. ДНК образует t-петлю, а одноцепочечный выступ внедряется в двухцепочечный участок ДНК, расположенный дистально (рис.6). Белковый комплекс позволяет клеткам отличать теломеры от мест разрыва хромосом (ДНК). Не все белки теломер входят в состав комплекса, который избыточен на теломерах, но отсутствует в других районах хромосом. Защитные свойства комплекса вытекают из его способности воздействовать на структуру теломерной ДНК по крайней мере тремя способами: определять структуру самого кончика теломеры; участвовать в образовании t-петли; контролировать синтез теломерной ДНК теломеразой. Родственные комплексы найдены и на теломерах некоторых других видов эукариот.
Рис. 6. Модель образования t-петлевой структуры на конце хромосомы.
Вверху - теломера в момент репликации хромосомы, когда ее конец доступен для комплекса теломеразы, который осуществляет репликацию (удвоение цепи ДНК на самом кончике хромосомы). После репликации теломерная ДНК (черные линии) вместе с находящимися на ней белками (показаны разноцветными овалами) образует t-петлю ( нижняя часть рисунка ).
Итак, у подавляющего большинства организмов основной тип последовательностей ДНК в центромерных и теломерных районах - это тандемно организованные мономеры короткой длины. Очевидно, что столь короткие последовательности (особенно в теломерах) обладают крайне ограниченной кодирующей способностью в первичной структуре и не соответствуют концепции Мёллера о терминальном гене [5].
В последние годы стало очевидным, что универсальных последовательностей ДНК, непосредственно определяющих функции центромер и теломер, нет. В этих районах хромосом ДНК служит платформой для сборки сложных, многокомпонентных ДНК-белковых комплексов, которые и обеспечивают выполнение этих функций. Более подробно о комплементарной организации этих комплексов и их координированного функционирования можно прочитать в нашем обзоре [2]. Наряду со специфическими для центромер и теломер компонентами этих комплексов в их состав входят и такие, которые участвуют в выполнении нескольких функций, иногда даже противоположных. Например, Ku70/80-гетеродимер входит в состав теломер и работает как позитивный регулятор длины теломер у дрожжей и негативный регулятор - у растения арабидопсис. В тоже время этот белок участвует в распознавании разрывов хромосом и их восстановлении. Без сомнения, одно из наиболее актуальных направлений исследований - выявление молекулярной природы механизмов регуляции разнообразных молекулярных комплексов, обеспечивающих активность центромер и теломер.
Работа выполнена при поддержке Российского фонда фундаментальных исследований (проект 04-04-48813), INTAS (03-51-5908)
и Программы интеграционных проектов СО РАН (проект 45/2).
1. Talbert P.B., Bryson T.D., Henikoff S. // J. Biol. 2004. V.3. Article 18.
2. Вершинин А.В. // Генетика. 2006. V.42. P.1200-1214.
3. Wu J., Yamagata H., Hayashi-Tsugane M. et al. // Plant Cell. 2004. V.16. P.967-976.
4. Scott K.C., Merrett S.L., Willard H.F. // Curr. Biol. 2006. V.16. P.119-129.
5. Muller H.J. Further studies on the nature and causes of gene mutations // Proc. Sixth Int. Congr. Genet. 1932. V.1. P.213-255.
Научная электронная библиотека

Геном человека – термин, используемый при описании всей генетической информации, закодированной в виде ДНК клеток человека. Клетка человека имеет два генома: сложный ядерный геном (хромосомная ДНК), содержащей более 99,9995 % всей генетической информации, и простой митохондриальный геном, в составе которого находится менее 0,0005 % ДНК. Ядерный геном распределен между 24-мя различными двуспиральными молекулами ДНК, которые в комплексе с различными гистоновыми и негистоновыми белками формируют хромосомы человека. С молекулярной точки зрения каждая хромосома клетки является сложно организованной структурой. Диплоидная клетка человека с интерфазным ядром при диаметре 5–10 мкм содержит около двух метров молекул ДНК, которые образуют 23 пары хромосом. Гаплоидный геном человека, который характеризуют половые клетки с хромосомами Х или Y, включает 23 различные хромосомы – 22 аутосомы и одну половую хромосому (гоносому) в зависимости от пола: Х – женский или Y – мужской пол. Индивидуальные хромосомы различаются своими морфологическими характеристиками. В среднем, хромосома человека содержит около 130 млн пар нуклеотидов (пн), однако содержание ДНК в разных хромосомах варьирует от 40 до 260 млн пн.
Данные о структуре, размерах и количестве митотических хромосом в клеточном ядре определяются как кариотип. Морфологическое строение хромосом на всех уровнях организации (молекулярном, микроскопическом и субмикроскопическом) определяется упаковкой нитей ДНК, организующих хромосому. На микроскопическом уровне нити ДНК формируют спирализованные хромонемы митотических хромосом. Упаковка хромонем по длине хромосом неравномерна. В них закономерно дифференцируются участки с резко выраженной и резко сниженной степенью спирализации хромонем. Последние формируют хромосомные перетяжки. Важным элементом структуры хромосом является первичная перетяжка, в участке которой расположена центромера. Она делит хромосому на два плеча – короткое (р) и длинное (q).
Структура хромосом претерпевает значительные изменения в ходе клеточного деления (клеточного цикла) и, следовательно, не является постоянной. Хромосомы интерфазных ядер (интерфазные хромосомы) в отличие от метафазных хромосом (см раздел, посвященный делению клетки) представляют собой более расправленные и диффузные клеточные структуры. Интерфазные хромосомы содержат одну хроматиду, в составе которой имеется одна двуспиральная молекула ДНК, в то время как метафазные хромосомы образованы двумя хроматидами и двумя молекулами ДНК. Хромосома как функционирующая клеточная органелла должна содержать минимум три типа последовательностей ДНК, формирующих её структурные компоненты: центромеру, теломеры и участок начала репликации ДНК.
Индивидуальные хромосомы содержат одну протяженную двуспиральную молекулу ДНК диаметром 2 нм, которая претерпевает несколько уровней упаковки, формируя хроматин или ДНК-белковый комплекс (дезоксирибонуклеопротеид – ДНП). Элементарной дискретной единицей упаковки хроматина является нуклеосома. Каждая нуклеосома имеет вид глобулы и состоит из четырех пар гистоновых белков Н2А, Н2В, Н3 и Н4, содержащих по 102–135 аминокислотных остатков. Вокруг гистоновой глобулы диаметром 10 нм (октамера) располагается молекула двуспиральной ДНК длиной 146 пн (или 68 нм), образуя виток в 1,75 оборота. Соседние нуклеосомы соединяются так называемой «спейсерной» ДНК длиной около 20 нм, образуя структуры в виде «бус на нитке». На следующем этапе компактизации 10-нанометровые нуклеосомы образуют фибриллы хроматина диаметром 30 нм (в каждом витке примерно 6 нуклеосом), которые образуют большие петли. На стадии метафазы степень компактизации достигает 1 на 10000 от первоначальной длины хромосомной ДНК. Петли 30-нанометровых фибрилл хроматина содержат 20–100 тыс. пн. ДНК на каждой петле прикрепляется к осевой структуре хромосомы (рис. 4). Осевые компоненты метафазной хромосомы состоят из негистоновых белков.
Как уже отмечалось, одним из структурных элементов хромосомы является центромера – место присоединения кинетохоров (макромолекулярных структур на белковой основе, участвующих в процессе расхождения хромосом при митотическом делении клетки), и слипания сестринских хроматид. Они также регулируют присоединение хромосом к веретену деления и направляют движение хромосом в ходе мейотического и митотического деления клетки. Хромосомы без центромеры (ацентрические фрагменты) не могут присоединяться к нитям веретена деления и теряются в ходе клеточного деления. У человека центромеры содержат особые типы повторяющихся ДНК, а также специфические «центромерные» белки, обеспечивающие сегрегацию хромосом в ходе митотического и мейотического деления клеток. В зависимости от положения центромеры в хромосоме она может быть метацентрической, субметацентрической и акроцентрической. В акроцентрических хромосомах довольно часто на коротких плечах видны небольшого размера хроматические тельца, так называемые спутники, и спутничные нити. Они содержат ядрышковый организатор – значимую структуру генома человека (рис. 5). Плечи некоторых хромосом человека содержат вторичные перетяжки.
Другими структурными элементами хромосом являются теломеры. Это специализированные структуры, содержащие особые типы ДНК и белки, которые образуют концевые участки хромосом. Теломеры выполняют несколько функций:
1) поддержание структурной целостности хромосомы;
2) обеспечение полной репликации концевых участков хромосомы;
3) поддержание организации хромосом в интерфазном ядре.
Рис. 4. Организация ДНК
Теломеры хромосом человека представляют собой повторы нуклеотидной последовательности ТТАГГГ, общий размер которой варьирует от 3 до 20 тыс. пн. Стабильность теломер поддерживается с помощью фермента теломеразы, нарушение функциональной активности которой приводит к нарушению структуры хромосом и, как следствие, к клеточной гибели. Укорочение последовательностей теломер, которому противодействует теломераза, связано с процессами старения и малигнизации.
Рис. 5. Метафазные хромосомы:
1 – метацентрическая хромосома; 2 – субметацентрическая хромосома; 3 – акроцентрическая хромосома. Видны:
а – центромеры; б – длинные плечи (q), в – короткие плечи (p);г – теломерные участки; д – спутники; е – спутничные нити
ДНК хромосом реплицируется в ходе периода синтеза ДНК клеточного цикла. Каждая хромосома содержит множество элементарных единиц репликации – репликонов, представляющих собой участки автономной репликации. Каждый репликон имеет одну точку инициации репликации, с которой начинается двунаправленный синтез ДНК (см предыдущий раздел, посвященный синтезу ДНК). Размеры репликонов могут различаться друг от друга. Репликоны у человека могут быть очень большими, достигая более чем 1 млн пн.
Строение хромосом: хроматин, центромера, теломера
Кафедра гистологии ХНМУ Histology KNMU запись закреплена
Хроматин. Уровни упаковки хроматина. Строение хромосомы
Хроматин (греч. χρώματα — цвета, краски) — это вещество хромосом, представляющее собой комплекс ДНК, РНК и белков. Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоида у прокариот. Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК.
До 25—40% сухого веса хроматина составляют гистоновые белки. Гистоны являются компонентом нуклеосом, надмолекулярных структур, участвующих в упаковке хромосом. Нуклеосомы располагаются довольно регулярно, так что образующаяся структура напоминает бусы. Нуклеосома состоит из гистонов четырёх типов: H2A, H2B, H3 и H4. Эти гистоны называются кóровыми. В одну нуклеосому входят по два кóровых гистона каждого типа — всего восемь белков. Линкерный гистон H1, более крупный, чем кóровые гистоны, связывается с ДНК в месте её входа на нуклеосому.
Нить ДНК с нуклеосомами образует нерегулярную соленоид-подобную структуру толщиной около 30 нанометров, так называемую 30 нм фибриллу. Дальнейшая упаковка этой фибриллы может иметь различную плотность. Если хроматин упакован плотно, его называют конденсированным или гетерохроматином, он хорошо видим под микроскопом. ДНК, находящаяся в гетерохроматине не транскрибируется, обычно это состояние характерно для незначащих или молчащих участков. В интерфазе гетерохроматин обычно располагается по периферии ядра (пристеночный гетерохроматин). Полная конденсация хромосом происходит перед делением клетки.
Если хроматин упакован неплотно, его называют эу- или интерхроматином. Этот вид хроматина гораздо менее плотный при наблюдении под микроскопом и обычно характеризуется транскрипционной активностью. Плотность упаковки хроматина во многом определяется модификациями гистонов — ацетилированием, фосфорилированием, метилированием и другими модификациями.
Считается, что в ядре существуют так называемые функциональные домены хроматина (ДНК одного домена содержит приблизительно 30 тысяч пар оснований), то есть каждый участок хромосомы имеет собственную «территорию». Вопрос пространственного распределения хроматина в ядре изучен пока недостаточно. Известно, что теломерные (концевые) и центромерные (отвечающие за связывание сестринских хроматид в митозе) участки хромосом закреплены на белках ядерной ламины.
Хромосо́мы (др.-греч. χρῶμα — цвет и σῶμα — тело) — нуклеопротеидные структуры в ядре эукариотической клетки, в которых сосредоточена бо́льшая часть наследственной информации и которые предназначены для её хранения, реализации и передачи. Хромосомы чётко различимы в световом микроскопе только в период митотического или мейотического деления клетки. Набор всех хромосом клетки, называемый кариотипом, является видоспецифичным признаком, для которого характерен относительно низкий уровень индивидуальной изменчивости.
Хромосома образуется из единственной и чрезвычайно длинной молекулы ДНК, которая содержит линейную группу множества генов. Необходимыми функциональными элементами хромосомы эукариот являются центромера, теломеры и точки инициации репликации. Точки начала репликации (сайты инициации) и теломеры, находящиеся на концах хромосом, позволяют молекуле ДНК эффективно реплицироваться, тогда как в центромерах сестринские молекулы ДНК прикрепляются к митотическому веретену деления, что обеспечивает их точное расхождение по дочерним клеткам в митозе.
Морфология метафазных хромосом
В ходе клеточного цикла облик хромосомы меняется. В интерфазе это очень нежные структуры, занимающие в ядре отдельные хромосомные территории, но не заметные как обособленные образования при визуальном наблюдении. В митозе хромосомы преобразуются в плотно упакованные элементы, способные сопротивляться внешним воздействиям, сохранять свою целостность и форму. Именно хромосомы на стадии профазы, метафазы или анафазы митоза доступны для наблюдения с помощью светового микроскопа. Митотические хромосомы можно увидеть у любого организма, клетки которого способны делиться митозом, исключение составляют дрожжи S.cerevisiae, чьи хромосомы слишком малы. Обычно митотические хромосомы имеют размеры в несколько микрон. Например, самая большая хромосома человека хромосома 1 имеет длину около 7 — 8 мкм в метафазе и 10 мкм в профазе митоза.
На стадии метафазы митоза хромосомы состоят из двух продольных копий, которые называются сестринскими хроматидами и которые образуются при репликации. У метафазных хромосом сестринские хроматиды соединены в районе первичной перетяжки, называемой центромерой. Центромера отвечает за расхождение сестринских хроматид в дочерние клетки при делении. На центромере происходит сборка кинетохора — сложной белковой структуры, определяющей прикрепление хромосомы к микротрубочкам веретена деления — движителям хромосомы в митозе. Центромера делит хромосомы на две части, называемые плечами. У большинства видов короткое плечо хромосомы обозначают буквой p, длинное плечо — буквой q. Длина хромосомы и положение центромеры являются основными морфологическими признаками метафазных хромосом.
В зависимости от расположения центромеры различают три типа строения хромосом:
акроцентрические хромосомы, у которых центромера находится практически на конце, и второе плечо настолько мало, что его может быть не видно на цитологических препаратах;
субметацентрические хромосомы с плечами неравной длины;
метацентрические хромосомы, у которых центромера расположена посередине или почти посередине.
Эту классификацию хромосом на основе соотношения длин плеч предложил в 1912 году российский ботаник и цитолог С. Г. Навашин. Помимо вышеуказанных трёх типов С. Г. Навашин выделял ещё и телоцентрические хромосомы, то есть хромосомы только с одним плечом. Однако по современным представлениям истинно телоцентрических хромосом не бывает. Второе плечо, пусть даже очень короткое и невидимое в обычный микроскоп, всегда присутствует.
Дополнительным морфологическим признаком некоторых хромосом является так называемая вторичная перетяжка, которая внешне отличается от первичной отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают различной длины и могут располагаться в различных точках по длине хромосомы. Во вторичных перетяжках находятся, как правило, ядрышковые организаторы, содержащие многократные повторы генов, кодирующих рибосомные РНК. У человека вторичные перетяжки, содержащие рибосомные гены, находятся в коротких плечах акроцентрических хромосом, они отделяют от основного тела хромосомы небольшие хромосомные сегменты, называемые спутниками. Хромосомы, обладающие спутником, принято называть SAT-хромосомами (лат. SAT (Sine Acid Thymonucleinico) — без ДНК).
Уровни компактизации хромосомной ДНК
Основу хромосомы составляет линейная макромолекула ДНК значительной длины. В молекулах ДНК хромосом человека насчитывается от 50 до 245 миллионов пар азотистых оснований. Суммарная длина ДНК из одной клетки человека составляет величину порядка двух метров. При этом типичное ядро клетки человека, которое можно увидеть только при помощи микроскопа, занимает объём около 110 мкм³, а митотическая хромосома человека в среднем не превышает 5 — 6 мкм. Подобная компактизация генетического материала возможна благодаря наличию у эукариот высокоорганизованной системы укладки молекул ДНК как в интерфазном ядре, так и в митотической хромосоме. Надо отметить, что у эукариот в пролиферирующих клетках осуществляется постоянное закономерное изменение степени компактизации хромосом. Перед митозом хромосомная ДНК компактизуется в 105 раз по сравнению с линейной длиной ДНК, что необходимо для успешной сегрегации хромосом в дочерние клетки, в то время как в интерфазном ядре для успешного протекания процессов транскрипции и репликации хромосоме необходимо декомпактизоваться. При этом ДНК в ядре никогда не бывает полностью вытянутой и всегда в той или иной степени упакована. Так, расчётное уменьшение размера между хромосомой в интерфазе и хромосомой в митозе составляет всего примерно 2 раза у дрожжей и 4 — 50 раз у человека.
Упаковка ДНК в хроматин обеспечивает многократное сокращение линейных размеров ДНК, необходимое для размещения её в ядре. Она происходит в несколько этапов. Наиболее изученными являются три первых уровня упаковки: (1) накручивание ДНК на нуклеосомы с образованием нуклеосомной нити диаметром 10 нм, (2) компактизация нуклеосомной нити с образованием так называемой 30-нм фибриллы и (3) сворачивание последней в гигантские (50 — 200 тысяч п. н.) петли, закреплённые на белковой скелетной структуре ядра — ядерном матриксе.
Одним из самых последних уровней упаковки в митотическую хромосому некоторые исследователи считают уровень так называемой хромонемы, толщина которой составляет около 0,1 — 0,3 мкм. В результате дальнейшей компактизации диаметр хроматиды достигает ко времени метафазы 700 нм. Значительная толщина хромосомы (диаметр 1400 нм) на стадии метафазы позволяет, наконец, увидеть её в световой микроскоп. Конденсированная хромосома имеет вид буквы X (часто с неравными плечами), поскольку две хроматиды, возникшие в результате репликации, соединены между собой в районе центромеры.
Читайте также:
- Лучевой туннельный синдром
- Факторы риска атеросклероза. Классификация факторов риска атеросклероза.
- Столбняк. Возбудитель столбняка. Clostridium tetani. Эпидемиология столбняка. Морфология столбняка. Тинкториальные свойства столбняка.
- Неврологическое обследование
- Неотложная помощь при острой почечной недостаточности.