Строение и функции ксилемы
Добавил пользователь Alex Обновлено: 30.01.2026
Состав ксилемы.В состав ксилемы входят проводящие, механические, запасающие и некоторые другие элементы. Рассмотрим подробнее проводящие элементы как наиболее важные, определяющие характер всей ткани. Остальные элементы будут рассмотрены позднее.
Строение и функции трахеальных элементов.Различают два типа проводящих, или трахеальных элементов — трахеиды и трахеи, или сосуды (иногда к трахеальным, элементам причисляют и древесинные волокна, поскольку между ними и трахеидами нет резкой границы).
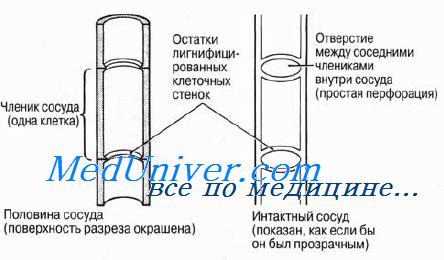
Трахеида представляет собой сильно вытянутую в длину водопроводящую клетку с ненарушенными первичными стенками. Проникновение растворов из одной трахеиды в другую происходит путем фильтрации через эти стенки, точнее, через окаймленные поры. Сосуд состоит из многих клеток, называемых члениками сосуда. Членики расположены друг над другом, образуя трубочку. Между соседними члениками одного и того же сосуда путем растворения возникают сквозные отверстия (перфорации). По сосудам растворы передвигаются значительно легче, чем по трахеидам.
Схема строения и сочетания трахеид (1) и члеников сосуда (2) (отдельная трахеида и членик сосуда зачернены)
В сформировавшемся, т.е. в функционирующем, состоянии трахеальные элементы состоят лишь из оболочек, так как протопласты распадаются, а полости трахеальных элементов заполняются растворами.
Сосуды и трахеиды передают растворы не только в продольном, но и в поперечном направлении в соседние трахеальные элементы и в живые клетки. Боковые стенки трахеид и сосудов сохраняются тонкими на большей или меньшей площади. Но в то же время они обладают различного рода утолщениями, придающими стенкам прочность. В зависимости от характера боковых стенок трахеальные элементы называются кольчатыми, спиральными, сетчатыми, лестничными и точечно-поровыми.
Типы утолщения (1—5) и поровости (6—8) боковых стенок у трахеальных элементов:
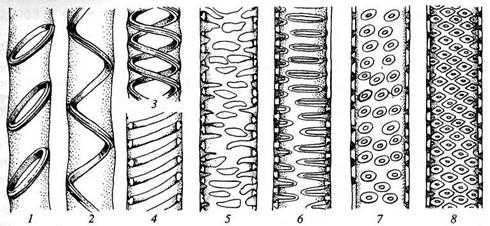
Вторичные кольчатые и спиральные утолщения прикрепляются к тонкой первичной оболочке посредством узкого выступа. При сближении утолщений и затем при образовании между ними перемычек возникает сетчатое утолщение, переходящее в окаймленные поры.
Эту серию можно рассматривать как морфогенетический, эволюционный ряд. Но эта же последовательность (хотя представленная не всеми членами) возникновения сначала кольчатых и спиральных элементов, а потом остальных наблюдается в гистогенезе одного и того же проводящего пучка. Наличие различных элементов в одном пучке объясняется тем, что первые из них возникают в то время, когда еще не закончился рост в длину всего органа, а кольчатые и спиральные элементы не препятствуют этому росту, так как способны растягиваться. Когда рост органа прекращается, возникают сетчатые, лестничные и точечно-поровые элементы, неспособные к продольному растяжению.
Эволюция трахеальных элементов.Трахеиды появились у высших растений в связи с выходом на сушу. Они найдены у ринии и других риниофитов. Сосуды появились значительно позже путем преобразования трахеид.
Эволюция перфораций:
1 - лестничная поровость трахеиды (перфорации отсутствуют); 2,3- лестничные перфорации; 4, 5- простые перфорации
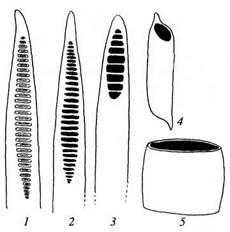
На рисунке показаны превращение трахеиды в членик сосуда и постепенное видоизменение последнего в процессе эволюции. Окаймленные поры превратились в сквозные отверстия — перфорации. Концы возникших сосудов, первоначально сильно скошенные, заняли горизонтальное положение, а примитивная лестничная перфорация со многими перекладинами превратилась в простую перфорацию. Параллельно этому членики сосудов становились более короткими и широкими. Сосудами обладают почти все покрытосеменные растения. Папоротникообразные и голосеменные растения, как правило, лишены сосудов и обладают только трахеидами. Лишь в виде редкого исключения сосуды встречены у таких высших споровых, как селагинелла, хвощи и некоторые папоротники, а также у немногих голосеменных (гнетовых). Однако у этих растений сосуды возникли независимо от сосудов покрытосеменных.
Возникновение сосудов у покрытосеменных растений означало важное эволюционное достижение, так как облегчило проведение воды; покрытосеменные растения оказались более приспособленными к жизни на суше.
Гистогенез трахеальных элементов.Членики сосуда возникают из живых клеток, которые имеют тонкие растяжимые оболочки и растут в ширину. После достижения окончательных поперечных размеров протопласт утолщает боковые стенки и в то же время растворяет поперечные стенки (образует перфорации). Превращение исходной живой клетки в зрелый членик сосуда (или в трахеиду) протекает быстро, иногда за несколько часов. За этот короткий срок протопласт выполняет большую работу, а затем разрушается. Соответственно этому изменяется его характер: на первых этапах в протопласте хорошо развиты структуры, ответственные за построение оболочки (эндоплазматический ретикулум, диктиосомы, микротрубочки). После утолщения боковых и растворения поперечных стенок развиваются лизосомы, происходят сильная вакуолизация и лизис всего протопласта. Остается оболочка, а полость заполняется жидкостью.
Схема гистогенеза членика сосуда:
1 - клетка, образованная камбием; 2 -растяжение; 3 -утолщение боковых стенок; 4 - образование перфораций и отмирание протопласта
Древесинные волокна имеют толстые оболочки и узкие простые (неокаймленные) поры. Древесинные волокна эволюционно возникли, как и сосуды, из трахеид, но их преобразование шло в другом направлении, именно в сторону потери проводящей функции и повышения механической прочности. Наличие волокон в составе древесины делает эту ткань прочнее.
Эволюционный переход от трахеиды 1 к волокнистой трахеиде 2 и к древесинному волокну 3
Строение и функции ксилемы
Синтез целлюлозы в плазматической мембране клетки
• Целлюлоза полимеризуется с участием комплексов, входящих в структуру плазматической мембраны
• Эти комплексы движутся по поверхности плазматической мембраны
Мы убедились в том, что клеточная стенка обеспечивает функционирование растительных клеток и что структура ее компонентов играет важную роль в их росте. Как клетка строит свою стенку? Особенно важно знать, каким образом она собирает основные компоненты, целлюлозные микрофибриллы, обеспечивающие прочность стенки.
Вначале считали, что целлюлоза образуется в самой стенке, с тем чтобы обеспечить восстановление слоев, разрушенных в процессе роста клетки. Сейчас известно, что в действительности целлюлоза полимеризуется с участием мультиферментного целлюлозо-синтезирующего комплекса белков, который встроен в плазматическую мембрану. Место синтеза целлюлозы в мембранах клеток растений, образующих новую стенку, было обнаружено, когда при приготовлении образцов для электронной микроскопии стали использовать метод замораживания-скалывания.
Этот метод позволяет разделить два слоя мембраны, и при последующем исследовании одного из слоев в электронном микроскопе эти комплексы видны в виде частиц. Как показано на рисунке ниде, на плазматической мембране растительной клетки видно много очень крупных трансмембранных комплексов характерной формы, которые представляют собой шесть частиц, собранных в виде гексагона. По размеру, каждая из этих гексагональных «розеток» напоминает рибосому и содержит много копий молекул ферментов, синтезирующих целлюлозу.
На рисунке ниже показано вероятное расположение в розетке отдельных копий молекул этих ферментов. Активированные формы глюкозы поступают на цитоплазматическую сторону мембраны, где используются комплексом для одновременной полимеризации множества целлюлозных нитей. Полимерные продукты постоянно выходят с наружной стороны мембраны. По мере синтеза целлюлозы розеткой, комплекс движется в плоскости мембраны, и за ним тянется новосинтезированный полимер. Относительно механизма движения розеток существует предположение, которое заключается в том, что при выходе линейного полимера, который скрепляется с клеточной стенкой, развивается усилие, обеспечивающее движение розетки по поверхности текучей плазматической мембраны.
В каждой розетке содержатся ферменты синтеза целлюлозы в количествах, достаточных для выработки 30-50 полимерных цепей, составляющих микрофибриллу. Эти линейные цепи упаковываются в параллельные пучки, образующие линейную полукристаллическую структуру — целлюлозную микрофибриллу. Поскольку отдельные цепи целлюлозы в длину составляют 1-5 мкм, в то время как микрофибриллы могут быть гораздо длиннее, чем 100 мкм (достаточно для того, чтобы обернуться вокруг большинства клеток), то концы отдельных целлюлозных цепей должны сплетаться по длине микрофибриллы.
Использование мембраносвязанных ферментов для синтеза основного волокна внеклеточного матрикса растительной клетки представляет собой уникальное явление и резко отличается от секреции коллагена, основного волокнистого белка внеклеточного матрикса клеток животных (коллаген секретируется в виде предшественника и собирается на поверхности клеток). За исключением каллезы, которая также синтезируется непосредственно на плазматической мембране, но с участием других синтаз, образование целлюлозы принципиально отличается от синтеза остальных компонентов клеточной стенки. Остальные компоненты предварительно собираются в клетке и экспортируются в составе везикул аппарата Гольджи, высвобождаясь при последующем их слиянии с плазматической мембраной.
Фотография в электронном микроскопе препарата плазматической мембраны клетки растения,
приготовленного методом замораживания-скалывания.
Видны несколько гексагональных «розеток». На врезке представлена одна из розеток при большом увеличении. Каждая зрелая розетка содержит 36 копий фермента синтазы целлюлозы.
Такая сборка структуры позволяет образующимся цепям целлюлозы объединяться в микрофибриллу. За движущимся комплексом синтеза целлюлозы постоянно происходит сборка микрофибрилл.
Комплекс перемещается в плоскости мембраны.
Пространственная близость полимеров целлюлозы, выходящих из розетки, обеспечивает их сборку сразу же после синтеза.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Проводящие ткани – флоэма и ксилема, структура, функции, происхождение.
Ксилема.Восходящий ток - ток минеральных солей, растворенных в воде, идущих от корней по стеблю к листьям. Восходящий ток осуществляется по сосудам и трахеидам ксилемы (древесины).
Ксилема состоит из:
1. Проводящих элементов (сосуды и трахеиды)
2. Механических элементов (волокна либриформа = древесинные волокна)
3. Запасающих элементов (древесинная паренхима)
Проводящие элементы ксилемы. Наиболее древние проводящие элементы ксилемы трахеиды представляют собой вытянутые клетки с заостренными концами. Трахеиды имеют одревесневшую клеточную стенку. По характеру утолщения оболочек, размерам и расположению в них участков первичных оболочек различают четыре типа трахеид: кольчатые, спиральные, пористые и лестничные. К наиболее древним относят лестничные трахеиды.
Сосуды (или трахеи) представляют собой однорядный продольный тяж клеток, называемых члениками. В филогенезе членики трахеи произошли из трахеид. Благодаря перфорациям между члениками вдоль всего сосуда свободно осуществляется ток жидкости. Утолщения клеточных оболочек у сосудов, так же как и у трахеид, бывают кольчатыми, спиральными, лестничными, сетчатыми и пористыми.
· Мертвые клетки, без протопласта
· Одревесневшие вторичные оболочки
· Трахеиды не имеют перфораций, растворы фильтруются через поры
· Сосуды имеют перфорации – сквозные отверстия в КС
Флоэма. Нисходящий ток - ток органических веществ, направляющийся от листьев к корням по ситовидным элементам флоэмы (луба).
Флоэма состоит из:
1. Проводящих элементов (ситовидные трубки и клетки-спутницы)
2. Механических элементов (лубяные волокна = камбиформ)
3. Запасающих элементов (лубяная паренхима)
Проводящие элементы флоэмы. У архегониальных растений, кроме мхов, проводящие элементы флоэмы представлены ситовидными клетками. На их продольных стенках имеются сквозные отверстия, напоминающие сито, и поэтому их называют ситовидными полями.
У покрытосеменных растений в процессе эволюции сформировался второй тип проводящих элементов - ситовидные трубки. Ситовидная трубка представляет собой продольный тяж клеток, называемых члениками. Конечные стенки члеников превращены в ситовидные пластинки - простые, если они имеют по одному ситовидному полю, и сложные, если есть несколько ситовидных полей. Ситовидные поля могут встречаться и на боковых стенках члеников (у березы). В ситовидном элементе разрушается тонопласт (оболочка вакуоли) и вакуолярный сок смешивается с гиалоплазмой. Ядро, как правило, разрушается. Каждому членику ситовидной трубки сопутствуют (одна или несколько) специализированные паренхимные клетки - клетки-спутницы, выполняющие вспомогательную роль в транспорте органических веществ, создавая нисходящий ток.
· Клетки живые, с первичными неодревесневшими клетками
· На стенках имеются ситовидные поля – группы мелких отверстий через которые соединяются протопласты соседних клеток
Ситовидные клетки – заметно вытянутые в длину клетки, с заостренными концами, ситовидные поля рассеяны по боковым стенкам.
- в зрелом состояние имеют ядра
- лишены ситовидных пластинок
- лишены сопровождающих клеток
- у высших споровых и голосеменных
Ситовидные трубки – состоят из члеников и сопровождающих клеток-спутниц. Характерны для покрытосеменных.
- в зрелом состоянии не имеют ядер, но имеют клетки-спутниц (с ядром)
- имеют ситовидные пластинки – ситовидные поля с более крупными отверстиями
Ксилема. Строение ксилемы. Функции ксилемы.
Ксилема выполняет в растении две основные функции: по ней движется вода вместе с растворенными минеральными веществами и она служит опорой органам растения. Таким образом, ксилема играет в растении двоякую роль — физиологическую и структурную. В состав ксилемы входят гистологические элементы четырех типов: трахеиды, сосуды, паренхимные клетки и волокна. На рис. 6.9 эти гистологические элементы представлены и поперечном и продольном разрезах.
Трахеиды ксилемы
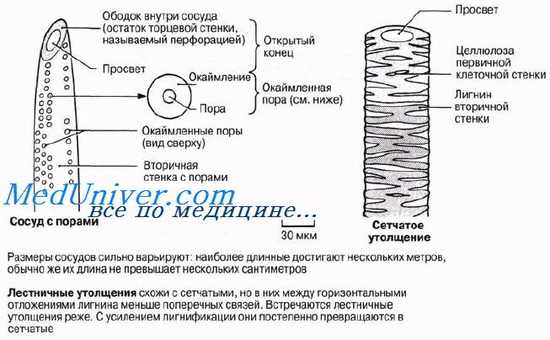
Трахеиды — это одиночные лигнифицированные клетки веретеновидной формы. Концы соприкасающихся трахеид перекрываются так же, как и заостренные концы волокон склеренхимы. Это придает трахеидам механическую прочность и обеспечивает органам растения опору. Трахеиды — мертвые клетки; в зрелом состоянии их просвет ничем не заполнен. Среди водопроводя-ших клеток сосудистых растений трахеиды представляют первичную примитивную форму; у древних сосудистых растений это единственные водопроводящие клетки. Из них развились описанные ниже сосуды и волокна ксилемы высших растений. Несмотря на свой примитивный характер, трахеиды, несомненно, функционируют эффективно; об этом свидетельствует тот факт, что у голосеменных растений доставка воды от корней к надземным частям обеспечивается исключительно трахеидами, а ведь большинство голосеменных — древесные породы. Вода движется по пустым просветам трахеид, не встречая на своем пути помех в виде живого содержимого. Из одной трахеиды в другую она переходит либо через поры, через их «замыкающие пленки», либо через нелигнифицированные части клеточных стенок. Характер лигнификации (одревеснения) клеточных стенок трахеид близок к тому, который описан ниже для сосудов.
На рисунке представлено строение трахеид. У покрытосеменных число трахеид по сравнению с числом сосудов относительно невелико. Сосуды считаются более эффективным приспособлением для транспорта воды, нежели трахеиды; появление сосудов связано, как полагают, с тем, что у покрытосеменных с их большой листовой поверхностью транспира-ция идет более активно.
Сосуды ксилемы
Сосуды — характерные проводящие элементы ксилемы покрытосеменных. Они представляют собой очень длинные трубки, образовавшиеся в результате слияния ряда клеток, соединившихся «конец в конец». Каждая из клеток, образующих сосуд ксилемы, соответствует трахеиде и называется члеником сосуда. Однако членики сосуда короче и шире трахеид. Первая ксилема, появляющаяся в растении в процессе сто развития, носит название первичной ксилемы; она закладывается у кончика корня и на верхушке побегов. Дифференцированные членики сосудов ксилемы появляются рядами на концах прокамбиальных тяжей. Сосуд возникает, когда соседние членики в данном ряду сливаются в результате разрушения перегородок между ними. Внутри сосуда сохраняются в виде ободков остатки разрушенных торцевых стенок. Слияние члеников сосудов изображено на рисунке.
Протоксилема и метаксилема
Первые по времени образования сосуды — протоксилема — закладываются на верхушке осевых органов, непосредственно под верхушечной меристемой, там, где окружающие их клетки еше продолжают вытягиваться. Зрелые сосуды про-токсилемы способны растягиваться одновременно с вытягиванием окружающих клеток, поскольку их целлюлозные стенки еще не сплошь одревеснели —лигнин откладывается в них лишь кольцами или по спирали (рис. 6.12). Эти отложения лигнина позволяют трубкам сохранять достаточную прочность во время роста стебля или корня. С ростом органа появляются новые сосуды ксилемы, которые претерпевают более интенсивную лигнификацию и завершают свое развитие в зрелых частях органа; так формируется ме-гаксшема. Тем временем самые первые сосуды протоксилемы растягиваются, а затем разрушаются. Зрелые сосуды метаксилемы не способны растягиваться и расти. Это мертвые, жесткие? полностью одревесневшие трубки. Если бы их развитие завершалось до того, как закончилось вытягивание окружающих живых клеток, то они бы очень сильно мешали этому процессу.
У сосудов метаксилемы обнаруживаются три главных типа утолщений: лестничные, сетчатые и точечные.
Длинные полые трубки ксилемы — идеальная система для проведения воды на большие расстояния с минимальными помехами. Так же как и в трахеидах, вода может переходить из сосуда в сосуд через поры или через неодревеснев-шие части клеточной стенки. Вследствие одревеснения клеточные стенки сосудов обладают высокой прочностью на разрыв, что тоже очень важно, потому что благодаря этому трубки не спадаются, когда вода движется в них под натяжением (разд. 13.4).
Вторую свою функцию — механическую — ксилема выполняет также благодаря тому, что она состоит из ряда одревесневших трубок. В первичном теле растения ксилема в корнях занимает центральное положение, помогая корню противостоять тянущему усилию надземных частей, изгибающихся под порывами ветра, В стебле проводящие пучки либо образуют по периферии кольцо, как у двудольных, либо располагаются беспорядочно, как у однодольных; в обоих случаях стебель пронизывается отдельными тяжами ксилемы, обеспечивающими ему определенную опору. Особенно важное значение опорная функция ксилемы приобретает там, где имеет место вторичный рост. Во время этого процесса быстро нарастает количество вторичной ксилемы; к ней переходит от колленхимы и склеренхимы роль главной механической ткани, и именно она служит опорой у крупных древесных и кустарниковых пород. Рост стволов в толщину определяется в известной мере нагрузками, которым подвергается растение, так что иногда наблюдается дополнительный рост, смысл которого состоит в усилении структуры и обеспечении ей максимальной опоры.
Древесинная паренхима ксилемы
Древесинная паренхима ксилемы содержится как в первичной, так и во вторичной ксилеме, однако в последней ее количество больше и роль важнее. Клетки древесинной паренхимы, подобно любым другим паренхимным клеткам, имеют тонкие целлюлозные стенки и живое содержимое.
Во вторичной ксилеме имеются две системы паренхимы. Обе они возникают из меристемати-ческих клеток, называемых в одном случае лучевыми инициалями, а вдругом — веретеновидны-ми инициалями (гл. 22). Лучевая паренхима более обильна. Она образует радиальные слои ткани, так называемые сердцевинные лучи, которые, пронизывая сердцевину, служат живой связью между сердцевиной и корой. Здесь запасаются различные питательные вещества, скапливаются таннины, кристаллы и т. п., и здесь же осуществляется радиальный транспорт питательных веществ и воды, а также газообмен по межклетникам.
Из веретеновидных инициалей обычно развиваются сосуды ксилемы и ситовидные трубки флоэмы вместе с их клетками-спутницами, однако время от времени они дают начало также и паренхимным клеткам. Эти паренхимные клетки образуют во вторичной ксилеме вертикальные ряды.
Древесинные волокна ксилемы
Полагают, что древесинные волокна, так же как и сосуды ксилемы, ведут свое происхождение от трахеид. Они короче и уже трахеид, а стенки их гораздо толще, но поры их сходны с порами, имеющимися в трахеидах, и на срезах волокна иногда трудно отличить от трахеид, поскольку между теми и другими есть ряд переходных форм. Древесинные волокна очень напоминают уже описанные волокна склеренхимы; их торцевые стенки также перекрываются. В отличие от сосудов ксилемы древесинные волокна не проводят воду; поэтому у них могут быть гораздо более толстые стенки и более узкие просветы, а значит, они отличаются и большей прочностью, т. е. придают ксилеме дополнительную механическую прочность.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
• Блоки клеток ксилемы претерпевают программированную гибель, образуя водопроводящие трубки
• Трубки защищены от внешних воздействий за счет образования поперечных утолщений вторичной стенки
• Микротрубочки кортекса образуют структуры, способствующие формированию профиля вторичных утолщений
До сих пор мы рассматривали организацию клеточных стенок и цитоплазмы при делении клеток и росте меристемы. Однако после прекращения деления клетки начинают дифференцироваться, образуя специализированную ткань. При дифференцировке происходит как общая, так и специфическая модификация клеточной стенки. Хотя стенки незрелых клеток, находящихся в области деления и роста, чрезвычайно прочны и способны противостоять силам тургора, они не обладают такой толщиной и жесткостью, чтобы функционировать в зрелых частях растения.
По мере дифференцировки, происходит утолщение клеточной стенки за счет добавления новых химически модифицированных слоев. Этот процесс называется вторичным утолщением. Утолщенные стенки клеток обеспечивают меристеме поддержку и по мере роста растения не дают ему упасть под тяжестью собственного веса.
Наряду с общим утолщением стенок всех клеток растения по мере дифференцировки, у некоторых из них происходят другие изменения стенки. Одним из примеров путей дифференцировкй растительных клеток в специализированную ткань является образование ксилемы. Находящиеся в корнях и побегах скопления или колонки элонгированных клеток дифференцируются с образованием соосудистых тканей — ксилемы и флоэмы. В ксилеме концы клеток примыкают друг к другу, образуя трубочки, проводящие воду от корневой системы ко всему растению, т. е. по всем его воздушным частям.
Вода и растворенные минеральные соли поглощается на большой площади поверхности, которая создается за счет корневых волосков, и через плазмодесмы, соединяющие корневые клетки, поступает в сердцевину растения и в ксилему. В наземных частях растения за счет солнечного нагрева, через отверстия устьиц листьев происходит ее испарение. Такое испарение или «транспирация» создает всасывающее усилие, благодаря которому вода «протаскивается» вверх по ксилеме. Этот ток воды, создающийся за счет транспирации и направленный снизу вверх, в структурном отношении важен, поскольку поддерживает тургор и предотвращает увядание растения.
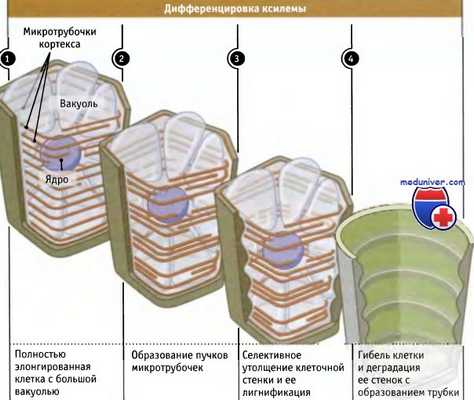
Последовательность событий при дифференцировке элемента трубки ксилемы.
При формировании непрерывных трубочек ксилемы (которые у некоторых деревьев могут достигать многих десятков метров в длину) дифференцировка составляющих их клеток имеет своеобразный характер. Процесс схематически представлен на рисунке ниже. Он начинается в клетках, которые вышли из зоны роста. Клеточные стенки у этих элонгированных и уже не растущих клеток утолщаются вторично. При этом не происходит равномерного утолщения, а в зависимости от вида клеток оно имеет определенный профиль (напоминающий кольца, лесенку, сетку, спираль и т. д.).
Общая особенность этих профилей состоит в том, что они в основном представляют собой поперечные полосы или ребра, которые укрепляют стенку зрелой ксилемы. Затем стенка модифицируется химически, в результате чего становится водонепроницаемой. После того как все эти подготовительные этапы заканчиваются, наступает программированная гибель клеток, и горизонтальные стенки между ними деградируют, в результате чего образуются непрерывные полые трубки, проходящие вдоль растения сверху донизу. На рисунке ниже видны несколько таких зрелых элементов ксилемы.
Из-за того что профили утолщения стенок этих трубок часто имеют форму обручей или ребер, они напоминают трахеи — воздухоносные пути нашей респираторной системы — и называются трахеальными элементами. Утолщения стенок предохраняют трахеальные элементы от сжатия под действием силы всасывания при транспирации и от сдавливания соседними клетками.
В экспериментах на декоративном растении Zinnia elegans были исследованы некоторые клеточные процессы, способствующие формированию трахеальных элементов. Интересно, что суспензию мезофильных клеток (клеток внутренней части листа), которая получается при осторожном размельчении листа, можно вырастить в культуре и индуцировать ее дифференцировку в одиночные трахеальные элементы.
В такой системе in vitro можно изучать некоторые процессы, которые должны происходить в растении. Как показано на рисунке ниже, кортикальные микротрубочки обычно образуют связки, формирующие несколько основных поперечно расположенных колец. Затем на эти кольца наслаиваются микрофибриллы целлюлозы, образуя массивные, вторично утолщающиеся ребра, основным компонентом которых является целлюлоза. Профиль расположения микротрубочек точно соответствует профилю утолщения стенок, и когда микротрубочки изменяются под действием химических соединений, вызывающих их деполимеризацию, у стенки возникает аномальный профиль. Среди микротрубочек также присутствуют актиновые филаменты. Это позволяет предполагать что обе системы филаментов совместно участвуют в организации вторичной клеточной стенки.
Одним из возможных механизмов формирования профиля клеточной стенки является доставка экзоцитозных везикул к местам утолщения стенки.
Такие везикулы содержат белковые комплексы синтеза целлюлозы, которые доставляются на плазматическую мембрану, и органичение доставки определенными сайтами ограничивает и отложение целлюлозы. В везикулах также содержатся такие секретируемые продукты, как полисахариды и структурные белки вторичной клеточной стенки. Как только утолщение стенки завершено, происходит ее модификация при взаимодействии с веществом, называемым лигнин. Он представляет собой гидрофобный полимер, который синтезируется за счет окислительной полимеризации до трех остатков ароматических спиртов (монолигнолов), и служит основным компонентом (20-35%) древесины. Благодаря своему гидрофобному характеру он создает гидроизоляцию внутренних стенок трубок. Лигнификация является дополнительным средством укрепления клеточных стенок, помогая им выдерживать вес частей растения, расположенных выше.
После завершения всех модификаций клеточной стенки происходит лизис клетки, и она погибает. Поперечные стенки между клетками диссоциируют, и остается утолщенная полая трубка, состоящая из зрелой специализированной ткани.
Фотография участка листа Zinnia в сканирующем электронном микроскопе.
Видны несколько трубочек ксилемы.
Представлен поперечный срез препарата,
и на внутренних стенках трубочек можно видеть кольцеобразные утолщения. Распределение микротрубочек в изолированной клетке Zinnia, дифференцирующейся в клетку ксилемы.
Микротрубочки промечены тубулином, содержащим флуоресцентный зонд,
и примерные границы клетки показаны с помощью пунктирной линии.
Клетка находится в начальной стадии процесса дифференцировки.
Организация микротрубочек в кортексе изменилась от относительно неупорядоченной,
характерной для большинства клеток, до крупных пучкоа расположенных вокруг длинной внутренней оси клетки.
Читайте также:
- Деформации челюсти при утомлении организма. Носовое дыхание и деформации челюсти
- Побочные эффекты лазеров и их безопасность
- Гуморальная регуляция секреции пищеварительных соков и моторики желудка и кишечника. Гормональная регуляция пищеварительного тракта.
- КТ при резидуальной кисте челюсти
- Ишемическая дисфункция левого желудочка. Метаболические нарушения ишемии миокарда