Векторы, плазмиды в молекулярной генетике. Что такое вектор, плазмида?
Добавил пользователь Алексей Ф. Обновлено: 29.01.2026
ФГБУН Институт биологии гена Российской академии наук, Москва, Россия, 119334
ФГБУН Институт биологии гена Российской академии наук, Москва, Россия, 119334
Оптимизированный метод получения рекомбинантных аденовирусных плазмид без использования электропорации
Журнал: Молекулярная генетика, микробиология и вирусология. 2019;37(4): 186‑191
ФГБУН Институт биологии гена Российской академии наук, Москва, Россия, 119334
Рекомбинантные аденовирусы широко используются для доставки и экспрессии трансгенов как для терапевтических целей, так и в фундаментальных исследованиях. Система аденовирусных векторов AdEasy позволяет быстро и эффективно создавать рекомбинантные репликативно-дефицитные аденовирусы. Получение рекомбинантной аденовирусной плазмиды за счет гомологичной рекомбинации в клетках E. coli BJ5183 между шаттл-вектором, кодирующим трансген, и плазмидой pAdEasy-1, несущей большую часть генома аденовируса человека серотипа 5 (Ad5), является одним из важнейших этапов при создании рекомбинантных аденовирусов. Во многих опубликованных протоколах указывается на необходимость трансформации шаттл-вектора и плазмиды pAdEasy-1 в клетки E. coli путем электропорации, что требует наличия соответствующего оборудования и расходных материалов. В данной работе нами был предложен оптимизированный протокол получения рекомбинантных аденовирусных плазмид с использованием только химически компетентных клеток E. coli, без применения электропорации. Показана эффективная трансформация неочищенного линеаризованного шаттл-вектора в химически компетентные клетки штамма E. coli BJ5183-pAdEasy-1, позволяющая идентифицировать клоны с корректными рекомбинантными аденовирусными плазмидами. С использованием оптимизированного протокола были получены четыре функциональных репликативно-дефицитных аденовируса, экспрессирующих целевые белки в клетках HEK293. Предложенный протокол позволяет существенно упростить и удешевить методологию получения рекомбинантных аденовирусов, так как отпадает необходимость использования оборудования и расходных материалов для электропорации.
ФГБУН Институт биологии гена Российской академии наук, Москва, Россия, 119334
ФГБУН Институт биологии гена Российской академии наук, Москва, Россия, 119334
В настоящее время аденовирусы широко применяются для доставки и экспрессии трансгенов для терапевтических целей и в фундаментальных исследованиях [1]. Система аденовирусных векторов AdEasy TM является одной из наиболее часто используемых для быстрого и эффективного создания рекомбинантных репликативно-дефицитных аденовирусов. В системе AdEasy рекомбинантная аденовирусная плазмида создается посредствам гомологичной рекомбинации в клетках E. coli BJ5183 между шаттл-вектором, кодирующим ген интереса, и плазмидой pAdEasy-1, несущей большую часть генома аденовируса человека серотипа 5 (Ad5). Далее полученная рекомбинантная аденовирусная плазмида трансфицируется в упаковочную клеточную линию (например, HEK293), в которой происходит сборка вирусных частиц и репликация вируса [2, 3].
Создание рекомбинантной аденовирусной плазмиды является одним из важнейших этапов получения рекомбинантных аденовирусов. В исходном варианте системы AdEasy в клетки штамма BJ5183 котрансформируют шаттл-вектор и плазмиду pAdEasy-1, отбирая канамицин-устойчивых рекомбинантов [2]. Повысить эффективность получения рекомбинантной аденовирусной плазмиды можно за счет создания штамма клеток E. coli BJ5183-pAdEasy-1, который получают путем трансформации клеток BJ5183 плазмидой pAdEasy-1. Далее в клетки штамма E. coli BJ5183-pAdEasy-1 трансформируют шаттл-вектор, что существенно увеличивает эффективность гомологичной рекомбинации по сравнению с котрансформацией двух плазмид в штамм BJ5183 [4].
Согласно приведенным выше протоколам [2, 4], для получения рекомбинантной аденовирусной плазмиды в системе AdEasy необходимо проведение электропорации плазмидной ДНК, что требует наличия дорогостоящего оборудования и расходников (кюветы), а также приготовления в лаборатории или приобретения высокоэффективных электрокомпетентных клеток E. coli. Однако в ряде работ для получения рекомбинатной аденовирусной плазмиды использовали химически компетентные клетки E. coli [5, 6].
Задачей данной работы была оптимизация протокола получения рекомбинатной аденовирусной плазмиды в системе AdEasy с использованием химически компетентных клеток, приготовленных в лабораторных условиях по одной из самых простых методик. В результате с использованием химически компетентных клеток нами были успешно созданы рекомбинантные аденовирусные плазмиды и получены репликативно-дефицитные аденовирусы, экспрессирующие целевые белки. Описанный нами протокол позволяет быстро и эффективно получать рекомбинантные аденовирусные плазмиды с помощью системы AdEasy без проведения электропорации, а также без дополнительной очистки линеаризованного шаттл-вектора.
Материал и методы
Плазмиды. Плазмиды pShuttle-CMV и pAdEasy-1 были приобретены у компании «Agilent Technologies» (США). Для создания шаттл-векторов, кодирующих кДНК слитого белка Egfp-Pdcd4 с мутациями в кДНК Pdcd4 RBM12 S457A или RBM12 D235A D418A, NheI (обработан фрагментом Кленова) — SalI фрагменты кДНК клонировали из соответствующих векторов pEgfp-C2-Pdcd4 RBM12 S457A или pEgfp-C2-Pdcd4 RBM12 D235A D418A в вектор pShuttle-CMV по сайтам BglII (обработан фрагментом Кленова) и SalI. Мутантная форма Pdcd4 RBM12, несущая аминокислотные замены K60A, R61A, R62A, R64A, K65A, R102A и R103A, была описана ранее [7]. Точечные мутации вводили с помощью сайт-направленного мутагенеза с помощью метода с двумя комплементарными олигонуклеотидными праймерами на основе протокола QuikСhange Site-Directed Mutagenesis kit («Agilent Technologies»). В качестве матрицы в ПЦР использовали плазмиду pCR-Blunt («Invitrogen», США), несущую кДНК белка Pdcd4 человека, фланкированную сайтами рестрикции EcoRI-SalI. ПЦР проводили с использованием ДНК полимеразы AccuPrime Pfx («Invitrogen») в реакции объемом 50 мкл, включавшей 1-кратный реакционный буфер, 300 нМ прямого и обратного праймеров, 100 нг плазмидной матрицы и 2,5 ед. ДНК-полимеразы при следующих условиях: 94 °C — 3 мин (94 °С — 40 с, 55 °C — 1 мин, 68 °C — 5 мин) — 15 циклов, 68 °C — 3 мин. Продукт ПЦР обрабатывали рестриктазой DpnI в течение 2 ч для расщепления плазмидной матрицы и трансформировали 10 мкл в химически компетентные клетки E. coli XL-1 Blue MRF’. Мутации в различных участках кДНК Pdcd4 вводили последовательно. Далее кДНК белка Pdcd4 с нужными мутациями субклонировали в вектор pEGFP-C2 («Clontech», США) по сайтам EcoRI-SalI. Для мутагенеза использовали следующие олигонуклеотидные праймеры: S457A (5`-gcagaaagcgttttgtagccgaaggagatggaggtc-3` и 5`-gacctccatctccttcggctacaaaacgctttctgc-3`); D235A (5’-gggacagtaatgagcacaactgctgtggaaaaatcatttgataaa-3’ и 5’-tttatcaaatgatttttccacagcagttgtgctcattactgtccc-3’); D418A (5’-ttccggacattaatctggctgtcccacattcatactc-3’ и 5’-gagtatgaatgtgggacagccagattaatgtccggaa-3’); K60A, R61A, R62A, R64A, K65A (5’-atcgcctctgccagagtcccgggatgagtttgccgctagtgccgctgctgccttggcattaattctagcttcgttaatg-3’ и 5’-cattaacgaagctagaattaatgccaaggcagcagcggcactagcggcaaactcatcccgggactctggcagaggcgat-3’); R102A, R103A (5’-caaagggaaggttgctggatgcggcatccagatctgggaaagg-3’ и 5’-cctttcccagatctggatgccgcatccagcaaccttccctttg-3’). Для создания шаттл-вектора, кодирующего внутриклеточный домен белка Notch1 крысы (аминокислотные остатки 1749 — 2531) c c-myc эпитопом на C-конце (NICD-myc), EcoRI (обработан фрагментом Кленова) — NotI фрагмент кДНК NICD-myc клонировали из вектора pEgfp-N2-NICD-myc в вектор pShuttle-CMV по сайтам BglII (обработан фрагментом Кленова) и NotI. Вектор для продукции белков с C-концевым c-myc эпитопом (pEGFP-N2-myc) создавали на основе вектора pEGFP-N2 («Clontech») путем клонирования по сайтам BamHI-NotI двухцепочечных олигонуклеотидов, полученных путем гибридизации одноцепочечных олигонуклеотидов, кодирующих аминокислотную последовательность c-myc эпитопа (5`-gatccgtcgacgagcagaagctgatctccgaggaggacctgtaagc-3` и 5`-ggccgcttacaggtcctcctcggagatcagcttctgctcgtcgacg-3`). кДНК NICD амплифицировали в ПЦР с праймерами 5`-gaattcgccaccatgaagcgcaggcggcagcat-3` и 5`-gtcgaccttaaatgcctctggaatg-3` и клонировали в вектор pEGFP-N2-myc по сайтам EcoRI-SalI, получая вектор pEgfp-N2-NICD-myc. Для создания шаттл-вектора, кодирующего протеинкиназу Pak5 человека, слитую с Egfp, NheI (обработан фрагментом Кленова) — SmaI фрагмент кДНК Egfp-Pak5 клонировали из вектора pEgfp-C1-Pak5, описанного ранее [8], в вектор pShuttle-CMV по сайту BglII (обработан фрагментом Кленова). Для генно-инженерных манипуляций с плазмидами и получения плазмидной ДНК в препаративных количествах использовали штамм E. coli XL-1 Blue MRF’ («Stratagene», США). Выделение плазмидной ДНК из клеток E. coli проводили с использованием набора GenElute Plasmid MiniPrep Kit («Sigma», США) в соответствии с протоколом производителя. Нуклеотидные последовательности всех амплифицированных в ПЦР кДНК верифицировали путем секвенирования ДНК с помощью набора реактивов ABI PRISM BigDye™ Terminator v. 3.1 с последующим анализом продуктов реакции на автоматическом секвенаторе Applied Biosystems 3730 DNA Analyzer в ЦКП «Геном» Института молекулярной биологии им. В.А. Энгельгардта РАН, Россия.
Культивирование клеток E. coli. Клетки E. coli культивировали в жидкой среде Miller’s LB broth (0,171 М NaCl, 1% триптон, 0,5% дрожжевой экстракт) («BD Bioscience», США) при +37 °С. Для заливки чашек Петри к среде LB добавляли бактериальный агар («BD Bioscience») до концентрации 1.5%. Клетки штамма E. coli BJ5183 («Agilent Technologies») культивировали в присутствии 30 мкг/мл стрептомицина, клетки штамма E. coli BJ5183-pAdEasy-1 — в присутствии 30 мкг/мл стрептомицина и 50 мкг/мл ампициллина, клетки штамма E. coli XL-1 Blue MRF’ — в присутствии 12,5 мкг/мл тетрациклина. В работе использовали следующие реагенты: ампициллин натрия (ОАО «Синтез», Россия), канамицин сульфат (ОАО «Биохимик», Россия), тетрациклин гидрохлорид («Sigma», США), стрептомицин (ОАО «Биохимик»).
Приготовление и трансформация химически компетентных клеток E. coli. Химически компетентные клетки штаммов E. coli BJ5183, E. coli BJ5183-pAdEasy-1 и E. coli XL-1 Blue MRF’ готовили в соответствии с протоколом, описанным в [9]. В частности, клетки соответствующих штаммов высевали на селективную чашку и растили в течение ночи. На следующий день инокулировали 10 мл ночной культуры среды LB без антибиотиков и растили в течение ночи. С утра переносили 2 мл ночной культуры в 200 мл среды LB без антибиотиков и растили в течение 3—4 ч до достижения оптической плотности 0,45—0,55 при длине волны 600 нм. Клетки охлаждали на льду в течение 2 ч и центрифугировали при 2500 g в течение 15 мин при +4 °С. Осадок ресуcпендировали в ледяном буфере (40 мМ натрия ацетат pH=5,5, 100 мМ CaCl2, 70 мМ MgCl2) и инкубировали на льду в течение 45 мин. Далее клетки осаждали центрифугированием при 1800 g в течение 10 мин при +4 °С, осадок ресуспендировали в буфере (40 мМ натрия ацетат pH=5,5, 100 мМ CaCl2, 70 мМ MgCl2, 15% глицерин), суспензию клеток разливали в аликвоты по 100 мкл, замораживали в жидком азоте и хранили при –80,°С. Трансформацию химически компетентных клеток E. coli проводили, как описано в [9]. В частности, к аликвоте компетентных клеток добавляли 3 мкл диметилсульфоксида, перемешивали, добавляли плазмидную ДНК, перемешивали и инкубировали на льду в течение 30 мин. Для проведения теплового шока клетки помещали на 50 с в водяную баню (+42 °С), потом быстро переносили в лед и инкубировали на льду 1 мин. Далее суспензию клеток переносили в 1 мл среды LB без антибиотиков и инкубировали клетки при интенсивном перемешивании при +37 °С в течение 1 ч, после чего клетки осаждали центрифугированием (30 с, 16 000 g), отбирали часть супернатанта и высевали клетки на предварительно прогретые до +37 °С чашки с LB агаром, содержавшим соответствующие антибиотики.
Результаты и обсуждение
В опубликованных протоколах создания рекомбинантных аденовирусов на основе системы AdEasy для трансформации клеток E. coli штаммов BJ5183 или E. coli BJ5183-pAdEasy-1 используется электропорация плазмидной ДНК, что обеспечивает высокую эффективность трансформации, до 1·10 7 КОЕ/мкг плазмидной ДНК при использовании коммерчески доступных электрокомпетентных клеток E. coli BJ5183-pAdEasy-1 [2—4]. В ряде других работ сообщалось об использовании химически компетентных клеток штамма E. coli BJ5183 для получения рекомбинантных аденовирусных плазмид путем котрансформации линеаризованных шаттл-вектора и аденовирусной плазмиды [5, 6], без использования штамма, аналогичного штамму E. coli BJ5183-pAdEasy-1 [4]. При этом проводили очистку шаттл-вектора из агарозного геля и переосаждение линеаризованной аденовирусной плазмиды перед котрансформацией [6] или оптимизацию соотношения количества шаттл-вектора и аденовирусной плазмиды [5]. Мы предположили, что протокол получения рекомбинантных аденовирусных плазмид можно оптимизировать и упростить за счет использования химически компетентных клеток E. coli, приготовленных по одной из простейших методик, создания клеток штамма E. coli BJ5183-pAdEasy-1 для повышения эффективности гомологичной рекомбинации и отказа от очистки линеаризованного шаттл-вектора из агарозного геля перед трансформацией.
Сначала готовили химически компетентные клетки E. coli BJ5183 и трансформировали их плазмидой pAdEasy-1. Эффективность трансформации химически компетентных клеток E. coli BJ5183 составляла 3·10 5 КОЕ/мкг плазмидной ДНК при использовании плазмиды pAdEasy-1 (33.4 т.п.н.). Обычно подобная эффективность считается недостаточной для использования компетентных клеток для генно-инженерных манипуляций, однако при трансформации очищенной плазмидной ДНК, и, учитывая большой размер плазмиды pAdEasy-1, подобная эффективность трансформации позволяет получить достаточное количество колоний на чашке для дальнейшего отбора и анализа корректных клонов. Далее, из нескольких колоний выделяли плазмиду pAdEasy-1 и анализировали ее с помощью эндонуклеаз рестрикции BglII и HindIII. Клетки одного из клонов с корректным паттерном рестрикции плазмиды pAdEasy-1 (данные не показаны) использовали для приготовления глицеринового стока клеток штамма E. coli BJ5183-pAdEasy-1 и далее готовили химически компетентные клетки штамма E. coli BJ5183-pAdEasy-1.
Таким образом, нами было установлено, что эффективность трансформации химически компетентных клеток штамма E. coli BJ5183-pAdEasy-1 неочищенным линеаризованным шаттл-вектором позволяет получить число маленьких колоний (890±270 в пересчете на 1 мкг линеаризованного шаттл-вектора), достаточное для идентификации корректной рекомбинантной аденовирусной плазмиды. Более того, нами было установлено, что химически компетентные клетки E. coli BJ5183-pAdEasy-1 после года хранения при –80 °С сохраняют эффективность трансформации, достаточную для идентификации клонов, содержащих аденовирусную плазмиду с корректной рекомбинацией (данные не показаны).
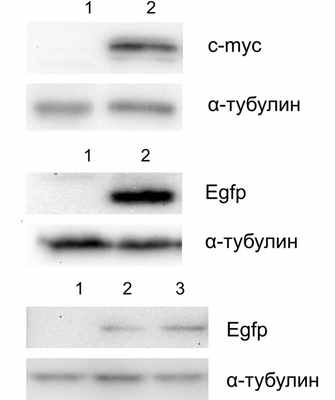
Далее для проверки функциональности рекомбинантных плазмид получали аденовирусы в соответствии с опубликованным протоколом [3]. Проводили 3 раунда амплификации вирусных стоков, после чего инфицировали клетки HEK293 для проверки продукции соответствующих белков. На рис. 2 показаны Рис. 2. Вестерн-блот анализ лизатов клеток линии HEK293, инфицированных рекомбинантными аденовирусами. Клетки HEK293 рассевали в количестве 1·10 5 в лунку 24-луночного планшета, на следующий день инфицировали аденовирусами, через 48 ч после инфекции получали клеточные лизаты и проводили Вестерн-блот анализ с использованием указанных антител. Клетки инфицировали аденовирусами для продукции белков (а) NICD-myc (дорожка 2), (б) Egfp-Pak5 (дорожка 2) и (в) Egfp-Pdcd4 RBM12 S457A и Egfp-Pdcd4 RBM12 D235A D418A, дорожки 2 и 3 соответственно. Дорожка 1 во всех панелях — неинфицированные клетки. результаты вестерн-блот анализа лизатов клеток, инфицированных стоками аденовирусов для продукции белков Egfp-Pdcd4 RBM12 S457A, Egfp-Pdcd4 RBM12 D235A D418A, Egfp-Pak5 и NICD-myc. Как видно, все полученные аденовирусы экспрессируют кодируемые шаттл-векторами кДНК.
Таким образом, нами был предложен оптимизированный протокол получения рекомбинантных аденовирусных плазмид на основе системы AdEasy, который не требует проведения электропорации и очистки линеаризованного шаттл-вектора. Нами было продемонстрировано эффективное получение рекомбинантных аденовирусных плазмид в результате трансформации химически компетентных клеток штамма E. coli BJ5183-pAdEasy-1, приготовленных в лаборатории с помощью простой и доступной методики. Химически компетентные клетки E. coli BJ5183-pAdEasy-1 сохраняют высокую эффективность трансформации на протяжении как минимум одного года, что позволяет быстро и эффективно создавать рекомбинантные аденовирусные плазмиды, используя единожды приготовленный запас компетентных клеток. Предложенный нами оптимизированный метод получения рекомбинантных адено-вирусных плазмид основан на использовании химически компетентных клеток, создание которых доступно для практически любой молекулярно-биологической лаборатории и не требует дорогостоящих реактивов, специальных навыков или оборудования. Кроме того, показанная нами эффективность трансформации компетентных клеток E. coli BJ5183-pAdEasy-1, в отличие от ранее опубликованных протоколов [3, 5, 6], позволяет отказаться от проведения очистки из геля или переосаждения линеаризованного шаттл-вектора, что также существенно упрощает процедуру получения рекомбинантных плазмид. Оптимизированный нами про-токол подтверждает возможность отказа от проведения электропорации, что делает создание рекомбинантных аденовирусов более доступным для широкого круга пользователей за счет отсутствия необходимости использования оборудования и расходных материалов для электропорации или приобретения высокоэффективных электрокомпетентных клеток. Валидность предложенного нами подхода была подтверждена успешным получением функциональных аденовирусов, экспрессирующих соответствующие рекомбинантные белки в клетках HEK293.
В работе была использована инфраструктура Центра коллективного пользования Института биологии гена Российской академии наук «Биология живой клетки и биомедицинские нанотранспортеры лекарств».
Финансирование. Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований (проекты № 16−04−00686 и № 16−04−00376).
Современные методы бионанотехнологии. Демонстрационный
Лекция 1.4. Определение вектора. Классификация векторов: по функциям, по происхождению, по месту применения. Челночные вектора. Состав вектора. Универсальность векторов. Вектора для клонирования. Вектора на основе плазмидной ДНК
Ферменты, описанные в предыдущем разделе, позволяют производить тонкие манипуляции как с протяженными молекулами ДНК, так и с их фрагментами. В частности, с помощью рестриктаз можно с большой точностью разрезать молекулы ДНК, а образовавшиеся в результате фрагменты соединять в любой желаемой последовательности друг с другом, восстанавливая сахаро-фосфатный остов молекулы ДНК с помощью ДНК-лигазы. Однако с использованием только этих ферментов еще нельзя решить одну из основных методических задач молекулярной генетики – выделение любой требуемой нуклеотидной последовательности в чистом виде и в количестве, достаточном для исследования этих последовательностей биохимическими методами. Исключение составляет метод ПЦР, однако его применение ограничивается короткими последовательностями нуклеотидов.
Основная идея, позволяющая решать эту задачу, заключается в том, чтобы присоединить исследуемые фрагменты ДНК к молекуле-переносчику, которая могла бы автономно существовать внутри бактериальных или эукариотических клеток в виде одной или нескольких копий и передаваться вместе со встроенным в нее фрагментом ДНК от одной клетки к другой. Такие молекулы-переносчики фрагментов нуклеиновых кислот были созданы, их называют векторами, и они являются одним из важнейших инструментов генной инженерии.
Идеальная векторная молекула должна обладать несколькими обязательными свойствами. Во-первых, любой вектор должен длительное время существовать в популяции клеток-хозяев, т.е. реплицироваться автономно или вместе с хромосомами клеток. Во-вторых, в любом векторе должны быть биохимические или генетические маркеры, которые позволяли бы обнаруживать его присутствие в клетках. В-третьих, структура векторной молекулы должна допускать встраивание в нее чужеродной последовательности нуклеотидов без нарушения ее функциональной целостности. Для конструирования векторов в генной инженерии используют небольшие молекулы нуклеиновых кислот, способные к автономной репликации в бактериальных и эукариотических клетках – плазмиды, хромосомы вирусов, а также фрагменты хромосом эукариотических клеток.
1.1.1.Плазмидные векторы
Различные векторы для клонирования ДНК и их рестрикционные карты
Обозначены положения уникальных сайтов рестрикции, а также функционально значимых генов
а – векторная плазмида pBR322; б – экспрессирующая векторная плазмида pUC18; в – векторная плазмида pAN7, предназначенная для отбора рекомбинантных клонов с использованием гомологичной рекомбинации; г – многофункциональная векторная плазмида Bluescript
Первые эффективные векторы для клонирования фрагментов чужеродной ДНК, не утратившие своего значения и поныне, были получены с использованием бактериальных плазмид. Большая серия векторных плазмид, обозначенных символом pBR, создана на основе репликона природной плазмиды ColEI, придающей клеткам E. coli устойчивость к колицину путем его объединения с генами устойчивости к антибиотикам. Таким образом, бактериальные клетки, несущие подобные комбинированные плазмиды, приобретали устойчивость к соответствующим антибиотикам, и их было легко отличить от бесплазмидных клеток путем простого посева на питательную среду с антибиотиками. Генетическая карта одного широко распространенного вектора этой серии – pBR322 изображена на рис. II.5,а. Такая плазмида представляет собой кольцевую ковалентно замкнутую молекулу ДНК длиной 4363 п.о. Последовательность нуклеотидов pBR322 полностью известна. Плазмида содержит гены устойчивости к тетрациклину (Tc) и ампициллину (Ap), которые были перенесены в плазмиду pBR322 из плазмиды pSC101 и транспозона Tn3 соответственно. Оба этих гена являются селектируемыми генетическими маркерами плазмиды, т.е. позволяют проводить отбор бактериальных клеток с плазмидой pBR322 по их способности к росту на питательных средах в присутствии тетрациклина и(или) ампициллина. Плазмида pBR322 содержит также обеспечивающий ее стабильную репликацию в клетках E. coli участок ДНК, который включает область начала репликации (ori). Характерной чертой плазмиды pBR322, как и любого современного вектора, является наличие в ней нескольких уникальных сайтов рестрикции, обозначенных на генетической карте. Следует иметь в виду, что встраивание в плазмиду клонируемых чужеродных фрагментов ДНК по сайтам рестрикции, расположенным в генах Ар или Tc (например PstI или BamHI), будет нарушать целостность этих генов и их функциональную активность. В результате происходит утрата бактериальными клетками, содержащими рекомбинантные плазмиды, устойчивости к соответствующим антибиотикам. По такому признаку легко различить бактериальные клетки, не содержащие плазмиды (не растут в присутствии ампициллина и тетрациклина), клетки с плазмидой, не содержащей вставки клонируемой ДНК (растут в присутствии обоих антибиотиков), и клетки с рекомбинантными плазмидами (в зависимости от локализации вставки могут расти на среде только с одним из двух вышеупомянутых антибиотиков). Следовательно, наличие в векторных молекулах селектируемых маркеров резко повышает эффективность клонирования из-за возможности проведения быстрого отбора рекомбинантных плазмид на селективных питательных средах.
Помимо генов устойчивости к антибиотикам в качестве селектируемых маркеров используют и другие гены или их фрагменты. В частности, для этих целей часто применяются гены различных ферментов, присутствие которых в клетках в составе плазмиды обнаруживают по появлению соответствующей ферментативной активности. В часто используемых векторах серии pUC таким селектируемым маркером является 5’-концевая часть гена b‑галактозидазы E. coli – lacZ’ (см. рис. II.5,б). Эта часть гена, находящаяся под контролем lac-промотора, кодирует N-концевую часть b-галактозидазы (так называемый a-пептид), которая путем объединения с недостающей С-концевой частью полипептида без образования пептидной связи восстанавливает ферментативную активность b-галактозидазы. b‑Галактозидаза обладает способностью расщеплять искусственный субстрат Xgal (5-бром-4-хлор-3-индолил-b-D-галактопиранозид) с образованием окрашенного в голубой цвет продукта реакции. В том случае, когда в бактериальных клетках, которые содержат в хромосоме недостающую экспрессирующуюся 3’-концевую часть гена lacZ и выращиваются на среде с Xgal, в результате комплементации a-пептидом вектора появляется активность b-галактозидазы, образованные этими клетками бактериальные колонии окрашиваются в голубой цвет. Уникальные сайты рестрикции для клонирования ДНК локализованы в начале гена b-галактозидазы в составе полилинкера (синтетической последовательности нуклеотидов, содержащей несколько перекрывающихся уникальных сайтов рестрикции), который не нарушает функциональной целостности последовательности нуклеотидов a-пептида. Вставка рекомбинантной ДНК в эти векторы разрывает структурную часть гена b-галактозидазы и инактивирует его, в связи с чем колонии бактерий с подобными рекомбинантными плазмидами, выросшие на питательной среде с Xgal, не окрашены. Поскольку векторы серии pUC одновременно содержат и ген устойчивости к ампициллину, отбор бактерий, несущих рекомбинантные плазмиды, можно проводить одновременно по этим двум маркерам. На питательной среде с ампициллином и Xgal вырастают только бактерии, устойчивые к антибиотику, т.е. содержащие плазмиду pUC, а среди выросших колоний лишь неокрашенные содержат вставку чужеродной ДНК. В качестве селектируемых маркеров в векторных молекулах часто используются гены, присутствие которых может быть обнаружено косвенно по комплементации генетических дефектов бактериальных клеток-хозяев, что делает их жизнеспособными в определенных селективных условиях.
За короткий период развития генной инженерии было сконструировано труднообозримое количество векторных плазмид, обеспечивающих конкретные потребности исследователей. Одной из вершин генно-инженерного искусства, прекрасно иллюстрирующей возможности генной инженерии, в настоящее время являются полифункциональные векторы серии Bluescript, полученные фирмой "Stratagene" (США) (см. рис. II.5,г). Вектор Bluescript M13+ представляет собой кольцевую ковалентно замкнутую молекулу ДНК длиной около 3 т.п.о. Он включает в себя ген устойчивости к ампициллину Amp r , ген b-галактозидазы lacZ, в N-концевую часть которого встроен полилинкер, содержащий уникальные сайты рестрикции для 21 рестриктазы, промоторно-операторную область lacZ, а также ген lac-репрессора lacI. В результате встраивания клонируемого фрагмента ДНК в полилинкер происходят разрыв кодирующей части гена lacZ и инактивация b-галактозидазы, что, как и в случае вектора pUC18, можно обнаружить по исчезновению окраски колоний бактерий, содержащих этот вектор со вставкой клонированной ДНК. Кроме того, встроенный в полилинкер фрагмент ДНК попадает под контроль промоторно-операторной регуляторной последовательности гена lacZ и в присутствии индуктора IPTG может быть экспрессирован в клетках E. coli. В дополнение к этому полилинкер в векторной плазмиде содержит на одном конце промотор для Т7-, а на другом – для Т3-РНК-полимераз, которые ориентированы навстречу друг другу. Это позволяет транскрибировать любую из цепей клонированного фрагмента ДНК in vitro с помощью той или другой РНК-полимеразы и получать препаративные количества мРНК или же комплементарной ей антисмысловой РНК. Кроме того, вектор Bluescript M13+ обладает межгенной областью (IG) фага f1, родственного фагу M13. Эта область детерминирует все цис-действующие функциональные последовательности нуклеотидов фага, необходимые для репликации его хромосомы и упаковки ее в фаговые частицы. В присутствии фага-помощника M13 происходит преимущественная упаковка образовавшейся в результате репликации одноцепочечной плазмиды в фаговые частицы M13. Одноцепочечная ДНК Bluescript M13+ после очистки может быть использована непосредственно для секвенирования клонированной ДНК или проведения сайт-специфического мутагенеза. Векторы типа Bluescript M13+, способные существовать либо в виде плазмиды, либо в составе фаговых частиц нитевидных бактериофагов, называют фагмидами.
Векторы, плазмиды в молекулярной генетике. Что такое вектор, плазмида?
Векторы, плазмиды в молекулярной генетике. Что такое вектор, плазмида?
Вектор — молекула ДНК, способная к автономному копированию в клетке-хозяине, например бактериальной или дрожжевой клетке, из которой ее можно впоследствии выделить в чистой форме для анализа. Если с помощью ДНК-лигазы включить в вектор фрагмент ДНК человека, такую новую молекулу ДНК можно внести в бактериальную клетку для размножения включенного фрагмента вместе с молекулой вектора.
Репликация векторов может привести к появлению большого числа копий на одну клетку, а бактерии-хозяева могут расти в лаборатории бесконечно долго, производя значительные и легкодоступные количества включенной ДНК-последовательности. Объединение молекулы ДНК из различных источников, например фрагментов ДНК человека и вектора, называется технологией рекомбинантной ДНК. Обычно с этой целью используют множество векторов, каждый со своими преимуществами и ограничениями, но мы ограничим внимание наиболее часто используемым вектором — плазмидой.
Плазмиды
Плазмиды, используемые как векторы — кольцевые двойные молекулы ДНК, отделившиеся от бактериальной или дрожжевой хромосомы, способные к копированию независимо от собственных хромосом микроорганизма. Векторные плазмиды — производные от молекул естественного происхождения, сначала были обнаружены в бактериях.
Поскольку они несли гены устойчивости к антибиотикам и могли легко переходить от одной бактерии к другой, плазмиды быстро распространяли устойчивость к антибиотику во всей микробной популяции.
Специально созданные для молекулярного клонирования плазмиды обычно невелики (несколько килобаз величиной) и содержат три обязательных компонента: оригинал для репликации (копирования в кишечной палочке или в дрожжах), один или более избирательных маркеров (например, ген, определяющий устойчивость к антибиотикам) и один или более сайтов рестрикции, которые могут быть использованы для удаления чужеродной ДНК.
Важные этапы, связанные с клонированием в плазмиде чужеродной ДНК с сайтом рестрикции EcoRI. Идентификация колоний, которые содержат искомую рекомбинантную плазмиду с последующим массовым ростом и выделением чистой плазмидной ДНК, позволяет накопить большие количества клонированной вставки. Некоторые плазмиды, оказавшиеся особенно полезными в ходе молекулярного клонирования, используются как ВАС (ВАС-векторы). ВАС-векторы — специально разработанные плазмиды, содержащие большие вставки ДНК, от 100 до 350 килобаз.
Разработка технологии ВАС потребовала многочисленных модификаций в генах плазмид и несущих бактерий, гарантирующих, что большие вставки чужеродной ДНК останутся стабильными и будут правильно копироваться в ходе деления бактерии-хозяина. Технология ВАС-векторов сыграла критически важную роль в проекте «Геном человека», позволив разделить весь геном человека на фрагменты, пригодные для анализа методом секвенирования.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Вектор (молекулярная биология)
Вектор (в генетике) — молекула нуклеиновой кислоты, чаще всего ДНК, используемая в генетической инженерии для передачи генетического материала другой клетке.
- плазмиды
- фазмиды
- векторы на основе вируса SV40
- векторы на основе аденовирусов
- векторы на основе герпесвирусов
- векторы на основе ретровирусов
- векторы на основе аденоассоциированного вируса
В качестве примера рассмотрим процесс клонирования участка чужеродной ДНК бактерией E. coli при помощи плазмиды pBR322 [1] .
pBR322 — искусственная плазмида, созданная Франциско Боливаром и Раймондом Родригесом с целью клонирования генетического материала. Она представляет собой циклический фрагмент ДНК длиной 4361 нуклеотидных пары (рис. 1). Плазмида содержит ген устойчивости к тетрациклину tet, взятый из естественной плазмиды pSC 101; ген устойчивости к ампициллину amp, взятый из транспозона Tn3; и участок начала репликации ori, заимствованный из плазмиды pMB 1. Тетрациклин и ампициллин — сильные антибиотики. Наличие в плазмиде генов устойчивости к ним (активных или блокированных) играет существенную роль в выделении бактерий со встроенным участком чужеродной ДНК. Плазмида содержит также сайты рестрикции Pst I, Bam HI и Sal I, причём первый находится в гене amp, а два остальных — в гене tet. Это важное обстоятельство помогает модифицировать плазмиду.
Предположим, что в плазмиду необходимо встроить фрагмент, который ранее был вырезан из другой ДНК рестриктазой Bam HI (то есть он имеет на концах последовательность нуклеотидов, характерную для сайта рестрикции Bam HI). Для этого плазмиды обрабатываются рестриктазой Bam HI, (которая разрежет кольцевую молекулу в сайте рестрикции и образует линейный участок ДНК) и добавляются участки чужеродной ДНК. Поскольку на концах всех фрагментов ДНК находятся комплементарные последовательности нуклеотидов, они начнут «склеиваться», причём возможны два варианта склейки (рис. 2) :
- Соединятся концы линейной плазмиды pBR 322, образовав исходную (восстановленную) кольцевую плазмиду.
- Между концами линейной плазмиды pBR 322 вклинится участок чужеродной ДНК, образовав кольцевую плазмиду со встроенным фрагментом.
Поскольку плазмиды со встроенным фрагментом являются целью процесса, необходимо выделить такие плазмиды и клонировать их. Процесс идёт следующим образом:
- Плазмиды внедряются в клетки E. coli. Для этого клетки обрабатываются ионами Ca 2+ , что делает их мембраны проницаемыми для ДНК.
- Полученные бактерии высевают на среду, содержащую ампициллин. В этой среде нормально растут колонии бактерий, содержащие плазмиды, остальные колонии угнетаются. По этому признаку можно отличить бактерии, содержащие плазмиды;
- Колонии, содержащие плазмиды, перепечатываются на среду, содержащую тетрациклин. Поскольку чужеродная ДНК вклинивается внутрь гена tet, дезактивируя его, колонии бактерий с модифицированными плазмидами угнетаются тетрациклином. Таким образом они визуально отличимы от колоний бактерий с восстановленными плазмидами.
- В результате этих действий выделяются колонии E. coli, в плазмиды которых встроен участок чужеродной ДНК. Они высеваются в обычную среду для дальнейшего клонирования.
Процедуру клонирования можно также вести с помощью рестриктаз Sal I и Pst I. В первом случае процесс будет аналогичен, в последнем бактерии с модифицированными плазмидами будут, наоброт, чувствительны к ампициллину и нечувствительны к тетрациклину.
Векторы, плазмиды в молекулярной генетике. Что такое вектор, плазмида?
Молекулярная генетика. Генная инженерия
Развитие молекулярной генетики с 70-х гг. в значительной степени основано на разработке и совершенствовании методов анализа и манипулирования ДНК. Так называемая «генная (генетическая) инженерия» имеет практические приложения во многих областях. Например, были разработаны новые методы диагностики и лечения заболеваний и стало возможным проводить направленные изменения определенных свойств организма. Поскольку полностью исключить биологический риск невозможно, при использовании методов генной инженерии необходим осторожный подход. В этом и последующих разделах дан краткий обзор методов, используемых в генной инженерии.
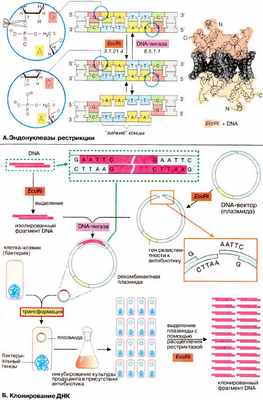
А. Эндонуклеазы рестрикции
Большинство методик в генной инженерии включают выделение определенных фрагментов ДНК (DNA) и последующее их соединение с другими фрагментами для получения новых комбинаций генов. Для этих целей используются ферменты, которые специфически разрезают и вновь сшивают молекулы ДНК. Наиболее важной группой ферментов являются эндонуклеазы рестрикции (рестриктазы), катализирующие специфическое расщепление двунитевой ДНК. Известно большое число рестриктаз. Для их обозначения используются сокращенные названия микроорганизмов - продуцентов. В качестве примера рассмотрим Eco RI — эндонуклеазу, выделенную из Escherichia coli. Подобно многим другим рестриктазам, этот фермент расщепляет ДНК по палиндромной последовательности, т. е. короткому сегменту ДНК, в котором обе цепи при считывании в направлении 5'→3' имеют одинаковую последовательность. Для Eco Rl это последовательность 5'-GAATTC-3'. Гомодимер Eco RI расщепляет фосфодиэфирные связи обеих цепей между G и А. Это приводит к образованию комплементарных « липких» концов (ААТТ), которые удерживаются вместе за счет спаривания оснований. Их, однако, можно легко отделить друг от друга путем небольшого нагревания. При охлаждении липкие концы гибридизуются вновь в правильной ориентации. Места расщепления можно соединить с помощью ДНК-лигазы.
Б. Клонирование ДНК
Обычно содержание в клетке какого-либо сегмента ДНК, например отдельного гена, очень незначительно. Поэтому для проведения экспериментов с фрагментами ДНК их необходимо многократно копировать ( клонировать ). В классической методике клонирования ДНК используется способность клеток бактерий поглощать и реплицировать короткие кольцевые молекулы ДНК, известные как плазмиды . Сначала клонируемый фрагмент ДНК вырезается из исходной ДНК с помощью рестриктазы (см. выше). Для демонстрации метода на схеме показано расщепление с помощью Eco RI. На практике обычно используются два разных фермента. В качестве переносчика ( «вектора» ) служит плазмида с единственным участком, узнаваемым Eco RI. Кольцевая плазмида линеаризуется с помощью Eco RI и затем смешивается с изучаемым фрагментом ДНК. Поскольку фрагмент и вектор имеют одинаковые липкие концы, некоторые из молекул будут гибридизоваться таким образом, что клонируемый фрагмент окажется интегрированным в структуру вектора. Затем концы линейной молекулы ковалентно сшиваются с помощью ДНК-лигазы с образовании новой («рекомбинантной") плазмиды. При обработке большого количества клеток некоторые из них поглощают рекомбинантную плазмиду (так называемая трансформация ). Трансформированные клетки реплицируют плазмиду вместе с собственным геномом. Обычно используют плазмиды, придающие трансформированной клетке устойчивость (резистентность) к определенному антибиотику. При инкубации популяции клеток в присутствии антибиотика будут реплицироваться только те клоны, которые содержат плазмиду. Из полученного клона выделяют плазмиду и после расщепления рестриктазой Eco RI получают множество копий клонированного фрагмента ДНК.
Читайте также:
- Карциноидные опухоли желудка. Неэпителиальные опухоли желудка.
- Кардиогенный шок. Гиповолемия при инфаркте миокарда.
- Трехстороннее отверстие. Топография трехстороннего отверстия. Четырехстороннее отверстие. Топография четырехстороннего отверстия.
- Патогенез кандидоза. Клиника кандидоза.
- Дренирование подногтевой гематомы