Значение пуфов (вздутий) политенных хромосом
Добавил пользователь Дмитрий К. Обновлено: 28.01.2026
пуффы (франц. pouf, англ. puff), утолщения на гигантских многонитчатых (политенных) хромосомах, являющиеся результатом деспирализации функционально активных участков хромосом с образованием открытых петель ДНК. Чем крупнее П. (крупные П. наз. кольцами Бальбиани) и чем более разрыхлён в них материал, тем выше скорость транскрипции (синтеза иРНК) в данном участке. Наиб, изучены П. в ядрах клеток слюнных желёз нек-рых двукрылых (комар-дергун, дрозофила). Число и положение П. на политенных хромосомах стабильно для каждой стадии онтогенеза особи, поэтому П. используют для определения возраста. Изучение динамики образования П. и, следовательно, активности отд. генов позволяет понять, как один и тот же хромосомный набор, принципиально сходный во всех клетках организма, участвует в дифференцировке разл. клеточных систем.
.(Источник: «Биологический энциклопедический словарь.» Гл. ред. М. С. Гиляров; Редкол.: А. А. Бабаев, Г. Г. Винберг, Г. А. Заварзин и др. — 2-е изд., исправл. — М.: Сов. Энциклопедия, 1986.)
Смотреть что такое "ПУФЫ" в других словарях:
ПУФЫ — в генетике вздувшиеся участки гигантских многонитчатых (политенных) хромосом, обнаруживаемые в слюнных железах некоторых двукрылых. В пуфах происходит активный синтез РНК (транскрипция), поэтому по динамике образования пуфов изучают механизм… … Большой Энциклопедический словарь
пуфы — (генетич.), вздувшиеся участки гигантских многонитчатых (политенных) хромосом, обнаруживаемые в слюнных железах некоторых двукрылых. В пуфах происходит активный синтез РНК (транскрипция), поэтому по динамике образования пуфов изучают механизмы… … Энциклопедический словарь
пуфы — ЭМБРИОЛОГИЯ ЖИВОТНЫХ ПУФЫ – участки локальной деспирализации хромосом, обеспечивающие в сочетании с биохимическими механизмами переход генов, локализованных в них, в активное состояние … Общая эмбриология: Терминологический словарь
Пуфы — в цитогенетике, вздутия, обнаруженные на т. н. политенных хромосомах; совокупность П. соответствует набору активных (функционирующих) генов в клетке на данной стадии её дифференцировки. Возникновение П. связано с деспирализацией… … Большая советская энциклопедия
ПУФЫ — (генетич.), вздувшиеся участки гигантских многонитчатых (политенных) хромосом, обнаруживаемые в слюнных железах нек рых двукрылых. В П. происходит активный синтез РНК (транскрипция), поэтому по динамике образования П. изучают механизмы развития и … Естествознание. Энциклопедический словарь
пуф — I. ПУФ I а, м. pouf m. 1. Мягкий низкий табурет. БАС 1. Они селу у стола: она на диван, аПетр Иванович на расстроившийся пружинами .. маленький пуф. Толст. Смерть Ив. Ильича. Серпухов по указанию лакея прошел ччерез ряд богато убранных комнат и… … Исторический словарь галлицизмов русского языка
Рабат — столица Марокко. Основан в XII в. как укрепленное поселение для борьбы с иноземцами и первоначально назывался Рибат эль Фатх, где араб. рибат укрепленный военный лагерь , пограничный военный пост , фатх победа , т. е. в целом лагерь победы .… … Географическая энциклопедия
цитогенетика — (от цито. и генетика), раздел генетики, изучающий закономерности наследственности и изменчивости на уровне клетки и субклеточных структур (главным образом хромосом). * * * ЦИТОГЕНЕТИКА ЦИТОГЕНЕТИКА, область генетики, изучающая цитологические… … Энциклопедический словарь
Ближний Восток — (Near East) Содержание Содержание 1. История Ближнего Востока Доисторический Период Древней Истории Период средневековый Крестовые Походы Период Новой Истории, с XV го по XVIII й в Период Новой Истории с XVIII по XX вв. Дело Святых Мест… … Энциклопедия инвестора
Политения — (от Поли. и лат. taenia повязка, лента) наличие в ядре некоторых соматических клеток гигантских многонитчатых (политенных) хромосом, превышающих в сотни раз обычные. П. приводит к значительному увеличению плоидности (См. Плоидность)… … Большая советская энциклопедия
Эндомитоз, полиплоидия, политения, амитоз. Примеры и значения
Ярыгин: На основе митотического цикла возник ряд механизмов, с помощью которых в том или ином органе количество генетического материала и, следовательно, интенсивность обмена могут быть увеличены при сохранении постоянства числа клеток. Удвоение ДНК клетки не всегда сопровождается ее разделением на две. Поскольку механизм такого удвоения совпадает с предмитотической редупликацией ДНК и оно сопровождается кратным увеличением количества хромосом, это явление получило название эндомитоза.
С генетической точки зрения, эндомитоз — геномная соматическая мутация, о чем будет сказано ниже. Другое явление, сходное по результату и названное политенией, заключается в кратном увеличении содержания ДНК в хромосомах при сохранении их диплоидного количества. Эндомитоз и политения приводят к образованию полиплоидных клеток, отличающихся кратным увеличением объема наследственного материала. В таких клетках в отличие от диплоидных, гены повторены более чем два раза. Пропорционально увеличению числа генов растет масса клетки, что повышает ее функциональные возможности. В организме млекопитающих полиплоидизация с возрастом свойственна печеночным клеткам.
Эндомито?з (от лат. эндо. и митоз) — процесс удвоения числа хромосом в ядрах клеток многих протистов, растений и животных, за которым не следует деления ядра и самой клетки. В процессе эндомитоза (в отличие от многих форм митоза) не происходит разрушения ядерной оболочки и ядрышка, не происходит образование веретена деления и не реорганизуется цитоплазма, но при этом (как и при митозе) хромосомы проходят циклы спирализации и деспирализации.
Повторные эндомитозы приводят к возникновению полиплоидных ядер, отчего в клетке увеличивается содержание ДНК.
Также эндомитозом называют многократное удвоение молекул ДНК в хромосомах без увеличения числа самих хромосом; как результат образуются политенные хромосомы. При этом происходит значительное увеличение количества ДНК в ядрах.
Пример: У винограда эндомитоз был обнаружен в кончиках молодых корней сорта «Фоль бланш». По своему происхождению, большинство известных полиплоидных сортов винограда возникло на основе соматических мутаций в результате спонтанного образования полиплоидных клеток путем эндомитоза. При определённых благоприятных условиях эти клетки занимают апикальное положение и, делясь в дальнейшем путём митоза, дают начало полиплоидным побегам на диплоидных кустах.
От таких побегов возникли, например, тетраплоидные клоны:
Значение: Генетическое и функциональное значение эндомитоза заключается в увеличении копийности (т.е. числа копий) генов. За счет этого клетка может получить больше продуктов этих генов (белков) и + увеличивается генетическая стабильность, т.к. при мутации одного гена остается еще масса неповрежденных копий этого гена. Это тупиковый путь регуляции экспрессии генов, поэтому он довольно редко встречается в природе. Т.е. увеличение количества белков - продуктов генов достигается не интенсивно, не путем увеличения экспрессии генов. а экстенсивно, путем увеличения количества копий гено. И стабильность генома достигается тоже экстенсивно (путем увеличения количества копий генов), а не путем совершенствования системы репарации.
Полиплоидия - (от греч. polyploos - многократный и eidos - вид) - наследственное изменение, заключающееся в кратном увеличении числа наборов хромосом в клетках организма. Широко распространена у растений (большинство культурных растений - полиплоиды), среди раздельнополых животных встречается редко. Полиплоидия може быть вызвана искусственно (напр., алкалоидом колхицином). У многих полиплоидных форм растений более крупные размеры, повышенное содержание ряда веществ, отличные от исходных форм сроки цветения и плодоношения. На основе полиплоидии созданы высокоурожайные сорта сельскохозяйственных растений (напр., сахарной свеклы).
Значение: Многие культурные растения полиплоидны, т. е. содержат более двух гаплоидных наборов хромосом. Среди полиплоидов оказываются многие основные продовольственные культуры; пшеница, картофель, онес. Поскольку некоторые полиплоиды обладают большой устойчивостью к действию неблагоприятных факторов и хорошей урожайностью, их использование и селекции оправдано.
ПОЛИТЕНИЯ (от поли … и лат. taenia - лента), наличие в ядрах некоторых соматических клеток гигантских многонитчатых (политенных) хромосом. Результат многократных удвоений хромосом, не сопровождающихся клеточным делением. Обнаружена в слюнных железах ряда двукрылых, а также у некоторых растений и простейших.
Амито?з, или прямо?е деле?ние кле?тки (от греч. α — частица отрицания и греч. μ?τος — «нить») — деление клеток простым разделением ядра надвое.
Впервые он описан немецким биологом Робертом Ремаком в 1841 году, термин предложен гистологом Вальтером Флеммингом в 1882 году. Амитоз — редкое, но иногда необходимое явление. В большинстве случаев амитоз наблюдается в клетках со сниженной митотической активностью: это стареющие или патологически измененные клетки, часто обреченные на гибель (клетки зародышевых оболочек млекопитающих, опухолевые клетки и др.).
При амитозе морфологически сохраняется интерфазное состояние ядра, хорошо видны ядрышко и ядерная оболочка. Репликация ДНК отсутствует. Спирализация хроматина не происходит, хромосомы не выявляются. Клетка сохраняет свойственную ей функциональную активность, которая почти полностью исчезает при митозе. При амитозе делится только ядро, причем без образования веретена деления, поэтому наследственный материал распределяется случайным образом. Отсутствие цитокинеза приводит к образованию двуядерных клеток, которые в дальнейшем не способны вступать в нормальный митотический цикл. При повторных амитозах могут образовываться многоядерные клетки.
В настоящее время считается, что все явления, относимые к амитозу — результат неверной интерпретации недостаточно качественно приготовленных микроскопических препаратов, или интерпретации как деления клетки явлений, сопровождающих разрушение клеток или иныепатологические процессы. В то же время некоторые варианты деления ядер эукариот нельзя назвать митозом или мейозом. Таково, например, деление макронуклеусов многихинфузорий, где без образования веретена происходит сегрегация коротких фрагментов хромосом.
Амитоз встречается в отживающих, дегенерирующих клетках, неспособных дать новые жизнеспособные клетки. В норме амитотическое деление ядер встречается в зародышевых оболочках животных, в фолликулярных клетках яичника. Амитотически делящиеся клетки встречаются при различных патологических процессах (воспаление, злокачественный рост и др.
Исчерченность
Характерная форма и размеры политенных хромосом достигаются вследствие их максимальной деспирализации и многократного воспроизедения хромосом без их последующего расхождения, то есть они образуются как результат эндомитоза [2] .
Политенные хромосомы имеют характерную поперечную исчерченность, обусловленную наличием участков более плотной спирализациихромонем — хромомер. В тёмных участках (то есть хромомерах) располагается спирализованный неактивный хроматин, в то время как тёмные полосы указывают на участок с повышенной транскрипционной активностью. Чёткое различение тёмных дисков и светлых междисковых участков объясняется нерасхождением дочерних хромонем. По этой причине все особенности отдельной хромонемы, в том числе хромомерный рисунок, становятся выраженными более контрастно [2] .
По сути, политенные хромосомы представляют собой пару гигантских гомологичных хромосом, находящихся в состоянии идеально точной соматической конъюгации. При этом диски и междисковые участки гомологов расположены строго паралелльно и тесно сближены. Такая конъюгация не характерна для подавляющего большинства соматических клеток [2] .
Впервые карта политенных хромосом была составлена в 1935 году Кэлвином Бриджесом (англ. Calvin B. Bridges), и она широко используется и по сей день.
Уникальность строения политенных хромосом, а именно возможность чётко различать детали их строения, была использована Т. Пайтнером для изучения их перестроек и характера конъюгации [2] . Вообще, исчерченность политенных хромосом исключительно полезна для исследований, в частности, на примере политенных хромосом была получена визуализация участков активного и неактивного хроматина. На них также можно изучать общую структуру хроматина.
Кроме того, политенные хромосомы помогают идентифицировать личинок комаров-звонцов (Chironomid), которых другим способом отличить сложно [3] [4] .
Пуфы
В политенных хромосомах процесс транскрипции сопровождается формированием т. н. пуфов — характерных вздутий определённых дисков, образующихся в результате локальной декомпактизации в них ДНК. На активную транскрипцию в этих регионах указывает активное включение 3 H-уридина в районе пуфов. Крупные пуфы называются кольцами Бальбиани (в некоторых источниках термины «пуф» и «кольца Бальбиани» употребляют как синонимичные) [2] .
Пуфирование характерно для стадии личинки. Образование и исчезновение пуфов регулируется внутренней средой организма в соответствии со стадией развития. Одним из важнейших регуляторов образования пуфов у насекомых являются стероидные гормоны, в частности, гормонлиньки — экдизон. Выявлено также влияние белков, синтезированных более ранними пуфами, на развитие более поздних пуфов [2] .
Таким образом, образование пуфов является ярким примером дифференциальной транскрипции. Другим известным примером этого процесса является формирование хромосом типа ламповых щёток [2] .
Функции[
Помимо увеличения размеров ядра и размеров клетки, политенные хромосомы, так как содержат большое число копий генов, усиливают ихэкспрессию. Это, в свою очередь, увеличивает производство необходимых специализированной клетке белков. Например, в клетках слюнных желёз личинок D. melanogaster хромосомы подвергаюся множеству кругов эндоредупликации, чтобы образовывать большое количество клейкого вещества до окукливания.
В других случаях тандемная дупликация участков, расположенных вблизи центромеры Х-хромосомы, наблюдающаяся в клетках слюнных желёз и кишечника, приводит к возникновнении мутации Bar, проявляющеся в изменении формы глаза [5] .
Гомологи́чные хромосо́мы — пара хромосом приблизительно равной длины, с одинаковым положением центромеры и дающие одинаковую картину при окрашивании. Их гены в соответствующих (идентичных) локусах представляют собой аллельные гены - аллели, т.е. кодируют одни и те же белки или РНК. При двуполом размножении одна гомологичная хромосома наследуется организмом от матери, а другая — от отца [1] .
В ходе удвоения ДНК в S-фазе интерфазы, предшествующей митозу, образуется две идентичные хроматиды, несущие один и тот же генетический материал. В дальнейшем в каждую дочернюю клетку попадает по одной такой хроматиде из пары хроматид данной хромосомы. В результате дочерняя клетка оказывается точной генетической копией материнской (это, правда, не касается тех случаев, когда имеют место различные мутации и перестройки) и имеет такое же число хромосом, что и материнская.
У диплоидных (2n) организмов геном представлен парами гомологичных хромосом. При мейозе гомологичные хромосомы обмениваются своими участками. Это явление лежит в основе рекомбинации генетического материала и носит название кроссинговер.
Гомологичные хромосомы не идентичны друг другу. Они имеют один и тот же набор генов, однако они могут быть представлены как различными (у гетерозигот), так и одинаковыми (угомозигот) аллелями, то есть формами одного и того же гена, ответственными за проявление различных вариантов одного и того же признака. Кроме того, в результате некоторых мутаций (дупликаций, инверсий, делеций и транслокаций) могут возникать гомологичные хромосомы, различающиеся наборами или расположением генов.
Диплоидные клетки — это живые клетки, в отличие от гаплоидных клеток (содержащих половинный набор), содержащие полный набор хромосом — по одной паре каждого типа. Большинство клеток человеческого организма являются диплоидными, за исключением гамет.
В норме в жизненном цикле организма человека происходит правильное чередование гаплоидной и диплоидной фаз развития клеток. Гаплоидные клетки образуются в результате мейоза и мейотического деления диплоидных клеток. После этого клетки могут размножаться при помощи митоза и митотических делений с образованием многоклеточного тела состоящего из диплоидных соматических клеток и нескольких поколений гаплоидных половых клеток (потомков).
Диплоидные клетки образуются из гаплоидных в результате полового процесса (оплодотворения, слияния половых клеток, гамет) с образованием зиготы.
Хроматин, его классификация. Строение хромосом.
В ядре клеток обнаруживаются мелкие зернышки и глыбки материала, который окрашивается основными красителями и поэтому был назван хроматином (от греч. chroma – краска).
Хроматин представляет собой дезоксирибонуклеопротеид (ДНП) и состоит из ДНК, соединённой с белка-ми-гистонами или негистоновыми белками. Гистоны и ДНК объединены в структуры, которые называются нук-леосомами. Хроматин соответствует хромосомам, которые в интерфазном ядре представлены длинными перекру-ченными нитями и неразличимы как индивидуальные структуры. Выраженность спирализации каждой из хромо-сом неодинакова по их длине. Реализацию генетической информации осуществляют деспирализованные участки хромосом.
Классификация хроматина. Различают два вида хроматина:
1) эухроматин, локализующийся ближе к центру ядра, более светлый, более деспирилизованный, менее компакт-ный, более активен в функциональном отношении. Предполагается, что в нем сосредоточена та ДНК, которая в интерфазе генетически активна. Эухроматин соответствует сегментам хромосом, которые деспирализованы и от-крыты для транскрипции. Эти сегменты не окрашиваются и не видны в световой микроскоп.
2) гетерохроматин- плотно спирализованная часть хроматина. Гетерохроматин соответствует конденсированным, плотно скрученным сегментам хромосом (что делает их недоступными для транскрипции). Он интенсивно окра-шивается основными красителями, и в световом микроскопе имеет вид тёмных пятен, гранул. Гетерохроматин располагается ближе к оболочке ядра, более компактен, чем эухроматин и содержит “молчащие” гены, т.е. гены, которые в настоящий момент неактивны. Различают конститутивный и факультативный гетерохроматин. Консти-тутивный гетерохроматин никогда не переходит в эухроматин и является гетерохроматином во всех типах клеток. Факультативный гетерохроматин может превращаться в эухоматин в некоторых клетках или на разных стадиях онтогенеза организма. Примером скопления факультативного гетерохроматина является тельце Барра – инактиви-рованная Х-хромосома у самок млекопитающих, которая в интерфазе плотно скручена и неактивна. В большинст-ве клеток оно лежит у кариолеммы.
Таким образом, по морфологическим признакам ядра (по соотношению содержания эу- и гетерохромати-на) можно оценить активность процессов транскрипции, а, следовательно, синтетической функции клетки. При её повышении это соотношение изменяется в пользу эухроматина, при снижении – нарастает содержание гетеро-хроматина. При полном подавлении функций ядра (например, в поврежденных и гибнущих клетках, при орогове-нии эпителиальных клеток эпидермиса – кератиноцитов, при образовании ретикулоцитов крови) оно уменьшается в размерах, содержит только гетерохроматин и окрашивается основными красителями интенсивно и равномерно. Такое явление называется кариопикнозом (от греч. karyon – ядро и pyknosis – уплотнение).
Хроматин и хромосомы представляют собой дезоксирибонуклеопротеиды (ДНП), но хроматин – это рас-крученное, а хромосомы – скрученное состояние. Хромосом в интерфазном ядре нет, они хромосомы появляются при разрушении ядерной оболочки (во время деления).
Распределение гетерохроматина (топография его частиц в ядре) и соотношение содержания эу- и гетеро-хроматина характерны для клеток каждого типа, что позволяет осуществить их идентификацию как визуально, так и с помощью автоматических анализаторов изображения. Вместе с тем, имеются определенные общие закономер-ности распределения гетерохроматина в ядре: его скопления располагаются под кариолеммой, прерываясь в об-ласти пор (что обусловлено его связью с ламиной) и вокруг ядрышка (перинуклеолярный гетерохроматин), более мелкие глыбки разбросаны по всему ядру.
Строение хромосом
Хромосомы представляют собой наиболее упакованное состояние хроматина. Наиболее компактные хромо-сомы видны на стадии метафазы, при этом они состоят из двух хроматид, связанных в области центромеры.
Роль положительных и отрицательных обратных связей различна. Отрицательные обратные связи обеспечивают стабильность функций живых систем, их устойчивость к внешним воздействиям. Они являются основным механизмом энергетического и метаболического баланса в живых системах, контроля численности популяций, саморегуляции эволюционного процесса. Положительные обратные связи играют позитивную роль усилителей процессов жизнедеятельности. Особую роль они играют для роста и развития. Рассмотри их подробней.
Отрицательная обратная связь (ООС) - тип обратной связи, при котором входной сигнал системы изменяется таким образом, чтобы противодействовать изменению выходного сигнала.
Отрицательная обратная связь делает систему более устойчивой к случайному изменению параметров.
Отрицательная обратная связь широко используется живыми системами разных уровней организации - от клетки до экосистем - для поддержания гомеостаза. Например, в клетках на принципе отрицательной обратной связи основаны многие механизмы регуляции работы генов, а также регуляция работы ферментов (ингибирование конечным продуктом метаболического пути). В организме на этом же принципе основана система гипоталамо-гипофизарной регуляции функций, а также многие механизмы нервной регуляции, поддерживающие отдельные параметры гомеостаза (терморегуляция, поддержание постоянной концентрации диоксида углерода и глюкозы в крови и др.). В популяциях отрицательные обратные связи обеспечивают гомеостаз численности. Физиологический смысл отрицательной Обратной связи заключается в том, что увеличение регулируемой величины (например, активности органа) сверх некоего предела вызывает понижающее воздействие со стороны сопряжённой с нею подсистемы; резкое уменьшение регулируемой величины обусловливает противоположное воздействие.
Отрицательная обратная связь также поддерживает температуру тела человека около 37 С.
Человек и все живые существа, являющиеся саморегулирующимися гомеостатичными системами, живут главным образом благодаря негативной обратной связи.
Положительная обратная связь (ПОС) - тип обратной связи, при котором изменение выходного сигнала системы приводит к такому изменению входного сигнала, которое способствует дальнейшему отклонению выходного сигнала от первоначального значения.
Положительная обратная связь ускоряет реакцию системы на изменение входного сигнала, поэтому её используют в определённых ситуациях, когда требуется быстрая реакция в ответ на изменение внешних параметров. В то же время положительная обратная связь приводит к неустойчивости и возникновению качественно новых систем, называемых генераторы (производители).
Положительная обратная связь рассогласует систему, и, в конечном счёте, существующая система трансформируется в другую систему, которая оказывается более устойчивой (то есть в ней начинают действовать отрицательные обратные связи).
Действие механизма нелинейной положительной обратной связи ведёт к тому, что система начинает развиваться в режиме с обострением.
Положительная обратная связь играет важную роль в макроэволюции. В целом, в макроэволюции положительная обратная связь приводит к гиперболическому ускорению темпов развития, что создает эффект равномерного распределения событий по логарифмической шкале времени.
На уровне простейших микроорганизмов, в которых еще нет нервных клеток, канал обратной связи (восприятие - реагирование) существовал и существует непосредственно на границе, отделяющей внешнюю среду организма от внутренней среды. Исходя из общепринятой сейчас теории возникновения жизни, рассмотрим, как работала обратная связь на уровне добиологическом. Например: простейшие многомолекулярные системы - коацерваты, уже имели зачатки обратной связи - обмен веществ своей внутренней среды с внешней. Возможно, одним из определяющих моментов возникновения жизни было образование мембраны, обладающей зачатками избирательной проницаемости и отделяющей систему от внешней среды.
Более подробно действие обратных связей можно рассмотреть на примере роста численности популяции некоторых видов, к примеру, мелких рыбёшек, в зависимости от наличия пищи (планктона) и наличия рыб-хищников. Чем больше пищи, тем большее число потомков рыбёшек может прокормиться и затем дать новое потомство. При неограниченном количестве пищи и отсутствии хищников и болезней у рыбешек их численность могла бы неограниченно возрастать. Здесь имеет место положительная обратная связь, выражающаяся в том, что процесс роста популяции рыбешек ведет к еще большему (в геометрической прогрессии) её росту. В случае наличия рыб-хищников возникает еще одна обратная связь: численность хищников будет влиять на количество корма для них (на количество мелких рыбешек). Эта обратная связь будет отрицательной. В результате действия обратных связей численность в популяциях является волнообразной и колебания численности будут происходить вокруг некоторого среднего уровня.
У высокоорганизованных животных деятельность центральной нервной системы в норме всегда включает как необходимое условие наличие обратной связи. Так, любое действие животного, например погоня за добычей, сопровождается импульсами, поступающими от центральной нервной системы к мышцам (бег, схватывание добычи), и обратными сигналами от органов чувств (зрение, проприорецепторы и др.), позволяющими учитывать результаты усилий и корректировать их в связи с ходом событий.
Саморегуляция процессов жизнедеятельности также обусловлена обратной связи. Так, подъём артериального давления выше нормы воспринимается специальными рецепторами, которые сигнализируют об этом в вазомоторные центры нервной системы. Это приводит к возникновению центробежных импульсов, ведущих к снижению давления. Подобный процесс - пример отрицательной обратной связи, наиболее часто наблюдаемой в стабильных живых системах. Большинство регуляторных систем животных и растительных организмов работает по этому принципу. Положительная обратная связь преобладают в период эмбрионального развития.
Многие процессы в экологии, например, регуляция динамики популяций, также основаны на положительной и отрицательной О. с. Так, особый случай отрицательной обратной связи представляет собой рассмотренная итальянским математиком В. Вольтерра система хищник - жертва. Увеличение численности жертв способствует усиленному размножению хищников, а рост численности последних, напротив, - снижению численности жертв. Хотя таким образом равновесие и поддерживается в природе, но благодаря запозданию в размножении животных оно приобретает форму волн жизни - широких колебаний численности животных вокруг среднего уровня.
На молекулярном уровне по принципу О. с. регулируется огромное число ферментативных реакций, одновременно протекающих в живой клетке. Координация этой сложной взаимосвязанной системы осуществляется путём изменения активности ферментов (отрицательную обратную связь осуществляют ингибиторы, положительную - стимуляторы) или скорости их синтеза (обратные связи осуществляют эффекторы).
Комбинации положительных и отрицательных обратных связей обусловливают альтернативную смену физиологических состояний (например, сон - бодрствование). Изучение кривой развития патологических процессов неинфекционного характера (трофические язвы, гипертония, маниакально-депрессивный психоз, эпилепсия и т.д.) позволяет, исходя из результата, определить наиболее вероятный тип Обратной связи, лежащий в основе заболевания, и ограничить изучение его этиологии и патогенеза механизмами определённой категории. Живые объекты как наиболее совершенные саморегулирующиеся системы богаты различными типами обратной связи; изучение последних - весьма продуктивно для исследования биологических явлений и установления их специфичности.
Таким образом, общие характеристики обратных связей могут быть сформулированы следующим образом:
Отрицательные обратные связи способствуют восстановлению исходного состояния. Положительные - уводят организм все дальше от исходного состояния.
Самоорганизация на всех уровнях начинается на основе механизмов положительной обратной связи, на которые затем накладываются ограничения отрицательных обратных связей.
Иммунитет (лат. immunitas — освобождение, избавление от чего-либо) — невосприимчивость, сопротивляемость организма к инфекциям и инвазиям чужеродных организмов (в том числе — болезнетворных микроорганизмов), а также воздействию чужеродных веществ, обладающих антигенными свойствами. Иммунные реакции возникают и на собственные клетки организма, измененные в антигенном отношении [1] .
Обеспечивает гомеостаз организма на клеточном и молекулярном уровне организации [1] . Реализуется иммунной системой.
Биологический смысл иммунитета — обеспечение генетической целостности организма на протяжении его индивидуальной жизни [2] . Развитие иммунной системы обусловило возможность существования сложно организованных многоклеточных организмов [3] .
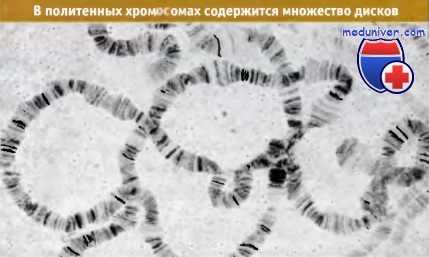
Значение пуфов (вздутий) политенных хромосом
Строение политенных хромосом
• Политенные хромосомы двукрылых характеризуются наличием серий полос-дисков, рисунок которых можно использовать в качестве цитологической карты
Интерфазные ядра в некоторых тканях личинок двукрылых мух содержат хромосомы во много раз большего размера, чем обычные хромосомы. У этих хромосом увеличены диаметр и длина. На рисунке ниже в качестве примера представлен хромосомный набор клеток слюнной железы D. melanogaster. Такие хромосомы называют политенными.
Каждая хромосома состоит из видимой серии полос-дисков (которые более правильно, но редко называют хромомерами). Существует примерно десятикратное различие в размере дисков. Наибольшая ширина составляет около 0,5 мкм, а наименьшая — 0,05 мкм. (Наименьшие диски видны только в электронном микроскопе.) В дисках находится большая часть массы ДНК, и они интенсивно прокрашиваются соответствующими красителями.
Участки между отдельными дисками окрашиваются светлее; их называют междисками. В хромосомном наборе D. melanogaster содержится около 5000 дисков.
Центромеры всех четырех хромосом D. melanogaster агрегируют с образованием хромоцентра, который в значительной степени состоит из гетерохроматина (а у самцов включает целиком Y-хромосому). С учетом этого можно считать, что около 75% гаплоидного набора ДНК организовано в диски и междиски. В растянутом состоянии эта ДНК представляет собой линейную структуру, длиной примерно 40 000 мкм. Длина всего хромосомного набора составляет примерно 2000 мкм, т. е. примерно в 100 раз превышает длину гаплоидного набора в митозе.
Это ясно показывает, в каком растянутом состоянии находится генетический материал в политенной хромосоме по сравнению с его обычными состояниями в интерфазном хроматине или в митотических хромосомах.
Политенные хромосомы D. melanogaster образуют серии дисков и междисков.
Какова структура этих гигантских хромосом? Каждая хромосома образуется в результате последовательных актов репликации конъюгировавшей пары гомологов. Реплики не разделяются, а остаются прикрепленными друг к другу по всей длине. В начале процесса каждая конъюгировавшая пара содержит количество ДНК, равное 2С (где С — это содержание ДНК в одной индивидуальной хромосоме). Затем число удвоений происходит до 9 раз; в результате максимальное содержание ДНК может достигнуть 1024С. Число удвоений варьирует в различных тканях личинки D. melanogaster.
Каждая политенная хромосома видна в виде большого числа параллельных нитей, сильно сконденсированных в дисках и слабее в междисках. Вероятно, каждая такая нить представлена единичной копией (С) гаплоидной хромосомы. Отсюда и происходит термин «политения». Степень политении определяется числом гаплоидных хромосом, содержащихся в гигантской хромосоме.
Для каждой линии Drosophila характерен свой набор дисков. Постоянство числа и линейного расположения дисков впервые было отмечено в 1930-х годах, когда стало понятно, что диски образуют цитологическую карту хромосом. В результате перестроек — таких как делеции, инверсии или дупликации, — происходит изменение линейного порядка дисков.
Линейный порядок дисков соответствует линейному расположению генов. Таким образом, генетические перестройки, как это видно по карте сцепления, можно привести в соответствие со структурными перестройками на цитологической карте. В конечном итоге, конкретную мутацию можно локализовать в определенном диске. Поскольку общее число генов у D. melanogaster превышает число дисков, в большинстве дисков, вероятно, локализуется множество генов.
Положение определенных генов на цитологической карте можно определить непосредственно с помощью метода гибридизации in situ. Общая схема метода представлена на рисунке ниже. Радиоактивная проба, представляющая некий ген (чаще всего меченный клон кДНК, полученный с иРНК), гибридизуется с денатурированной ДНК политенных хромосом in situ. Методом радиоавтографии можно определить местоположение одного или нескольких исследуемых генов по скоплению зерен серебра в области одного или нескольких дисков. Недавно вместо радиоактивных зондов стали использовать флуоресцентные.
Используя этот метод, оказывается возможным непосредственно выявить полосу, в которой локализована специфическая последовательность.
Индивидуальные диски, содержащие определенные гены, можно идентифицировать методом гибридизации in situ. При большом увеличении показана гибридизация in situ на дисках 87А и 87С с меченной ДНК,
выделенной из клеток, подвергнутых действию теплового шока.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
• Полосы-диски политенных хромосом, являющиеся сайтами генной экспрессии, образуют вздутия или «пуфы».
Одна из наиболее удивительных особенностей политенных хромосом — это возможность непосредственно наблюдать их активные участки. Некоторые диски иногда разбухают, образуя на хромосоме вздутия или пуфы; при этом происходит выпячивание хромосомного материала из осевой структуры. Пример очень больших пуфов (названных кольцами Бальбиани) изображен на рисунке ниже.
Какова природа пуфа? Пуф представляет собой участок, в котором хромосомные нити находятся в более релаксированном виде, чем когда они упакованы в диске. Эти нити остаются непрерывно связанными с нитями, находящимися в осевой части хромосомы. Обычно пуфы образуются из материала отдельных дисков, хотя некоторые особенно крупные пуфы, такие как кольца Бальбиани, разбухают так сильно, что заслоняют соседние диски.
Порядок расположения пуфов соответствует транскрипции генов. В процессе развития личинки пуфы появляются и исчезают в определенном тканеспецифическом порядке. Каждая ткань в любой момент развития характеризуется определенной картиной расположения пуфов.
Пуфы индуцируются гормоном экдизоном, который контролирует развитие Drosophila, некоторые пуфы индуцируются непосредственно гормонами, тогда как индукция других опосредована продуктами более ранних пуфов.
Пуфы — это сайты, в которых в данный момент происходит синтез РНК. Согласно общепринятой точке зрения, образование пуфов представляет собой увеличение размеров одного диска, связанное с необходимостью релаксации его структуры для синтеза РНК Поэтому образование пуфов рассматривают как следствие транскрипции. Пуф может образоваться при экспрессии одного активного гена.
Место, где образуется пуф, отличается от того места, на котором расположен обычный диск, присутствием в нем дополнительных белков, включая РНК-полимеразу II, а также других белков транскрипции.
Особенности хромосом типа ламповых щеток и политенных хромосом позволяют прийти к выводу: чтобы транскрипция могла осуществиться, генетический материал, обычно плотно упакованный, должен деконденсироваться. Однако еще остается вопрос, соответствует ли такое локальное расплетение на уровне целой хромосомы тем событиям, которые происходят на молекулярном уровне в общей массе обычного интерфазного хроматина.
Какую функциональную роль играют диски политенных хромосом, иными словами, соответствует ли каждый диск некой генетической единице? Можно было бы надеяться, что ответ на этот вопрос будет получен на основании результатов секвенирования генома дрозофилы, поскольку, сопоставляя области междисков с местонахождением определенных последовательностей ДНК, можно выяснить их локализацию. Однако пока не получено картины, позволяющей оценить функциональную роль дисков.

На хромосоме IV из слюнной железы С. tentans видны три кольца Бальбиани.
Читайте также:
- Отравление колониестимулирующими факторами и их побочные эффекты
- Перелом головчатой кости. Диагностика и лечение
- Осложнения вирусного гепатита. Начальная стадия печеночной энцефалопатии
- Обструкция протезированного клапана сердца. Регургитация через протезированный клапан сердца
- КТ, МРТ при болезни Кюммеля