Дефицит минеральных веществ. Насекомоядные растения.
Добавил пользователь Евгений Кузнецов Обновлено: 28.01.2026
летающие к клеткам корней или же проникающие в них. Гаустории высасывают питательные вещества (прежде всего углеводы) из растения, и можно предполагать, что механизм этого явления также построен на базе Н + -помпы.
Высшие растения-паразиты, использующие готовые органические вещества, — это, как правило, высокоспециализированные однолетники или многолетники с редуцированными или полностью утраченными в ходе эволюции листьями, а часто и корнями. Имеются виды, совершенно лишенные хлорофилла и не способные к фотосинтезу. К ним относится, например, заразиха (Oriobanche), паразитирующая на корнях многих культурных растений. Ее семена прорастают лишь под влиянием корневых выделений растения-хозяина. Как только кончик зародышевого корня проростка соприкоснется с корнем хозяина, он преобразуется в гаусторию (присоску), начинающую выделять гидролазы, растворяющие клеточные стенки, и активно внедряющуюся в корень. В период роста и развития заразиха поглощает большое количество азотистых веществ, углеводов и минеральных элементов, особенно фосфора, а также воду из корней растений-хозяев. В растениях томатов, пораженных заразихой, содержание, например, белкового азота снижается в 3 раза, а Сахаров — в 16 раз. Другой пример корневого паразита — петров крест (Lathraea squamaria), паразитирующий на корнях деревьев и кустарников.
У вьющегося паразитного травянистого растения повилики (Cuscuta) нитевидные стебли с редуцированными листьями-чешуйками обвиваются вокруг стеблей растений-хозяев и присасываются к ним с помощью гаусторий. Гаустории повилики — преобразованные адвентивные (придаточные) корни. Они принимают форму диска, плотно прилегающего к коре растения-хозяина. Группа клеток из центральной части диска внедряется в коровую паренхиму растения-хозяина и достигает центрального цилиндра, откуда повилика получает воду, органические вещества и минеральные элементы. Проростки повилики, совершая ростовые вращательные движения, находят растение-хозяина, реагируя на градиент влажности и выделяемые им вещества (явление хемотропизма — см. 13.5.2).
К паразитным растениям относится и раффлезия, питающаяся соками корней тропических лиан. В тело жертвы она внедряется с помощью гаусторий, выделяющих целлюлазу и другие ферменты, разрушающие клеточные стенки. Всю свою жизнь раффлезия проводит в теле хозяина — под землей. Лишь ее цветки появляются на поверхности почвы. С помощью радиоактивной метки показано, что паразиты поглощают из тела хозяина в основном сахарозу, глутамииовую и аспарагиновую кислоты и их амиды.
В настоящее время известно свыше 400 видов покрытосеменных растений, которые ловят мелких насекомых и другие организмы, переваривают свою добычу и используют продукты ее разложения как дополнительный источник питания. Большинство из них встречается на бедных азотом болотистых почвах, есть эпифитные и водные ормы.
Ловчие механизмы. Листья насекомоядных растений трансформированы в специальные ловушки. Наряду с фотосинтезом они служат для поимки добычи. По способу ее ловли насекомоядные растения можно разделить на две большие группы.
При пассивном типе ловли добыча может а) прилипать к листьям, желёзки которых выделяют липкую слизь, содержащую кислые полисахариды (библис, росолист), или б) попадать в специальные ловушки в виде кувшинов, урн, трубочек, окрашенных в яркие цвета и выделяющие сладкий ароматный секрет (саррацения, гелиамфора, дарлингтония).
Для активного захвата насекомых используются: 1) приклеивание добычи липкой слизью и обволакивание ее листом или волосками (жирянка, росянка), 2) ловля по принципу капкана — с захлопыванием ловчих листьев над добычей (аль-дрованда, венерина мухоловка), 3) ловчие пузырьки, в которые насекомые втягиваются с водой благодаря поддерживаемому в них вакууму (пузырчатка).
Общим для всех типов ловчих приспособлений является привлечение насекомых с помощью полисахаридных слизей или ароматного секрета (нектара), выделяемых или самими ловчими аппаратами, или желёзками вблизи от ловушки. Быстрые движения ловчих органов, как правило, осуществляются путем изменений в них тургора и запускаются с помощью распространяющихся потенциалов действия в ответ на раздражение чувствительных волосков, вызванное движениями насекомого (см. 13.6.1).
Пищеварение. Попавшее в ловушку или приклеившееся к липкой поверхности листа насекомое переваривается под действием секрета многочисленных желёзок (рис. 7.1). Некоторые насекомоядные парализуют добычу алкалоидами, содержащимися в выделяемой слизи. Так, росянка выделяет алкалоид ко-нитин, парализующий насекомое. Липкая слизь, секретируемая стоячими, или стебельчатыми, желёзками насекомоядных растений, содержит много кислых полисахаридов, состоящих из ксилозы, маннозы, галактозы и глюкуроновой кислоты. Кислая реакция слизи обеспечивается и находящимися в ней органическими кислотами. Слизь, выделяемая стоячими желёзками, содержит ряд гидролаз, активных в кислой среде. Например, у жирянки в слизи содержится кислая амилаза.
Кислые слизистые выделения, азот- и фосфорсодержащие продукты распада стимулируют работу сидячих желёзок, которые начинают выделять кислоты (муравьиную, бензойную), а также протеазы и ряд других гидролаз. Довольно подробно изучена протеолитическая активность секрета у мухоловки. Большая часть протеаз из ее секрета относится к тиоловым протеазам, среди них — карбоксипептидаза, хитинолитическая активность. Кроме протеаз в секрете обнаружены кислая фос-фатаза и эстераза, а у некоторых видов насекомоядных — рибо-нуклеаза, липаза, а также пероксидаза.
Секреторные клетки насекомоядных растений имеют хорошо развитый ЭР и аппарат Гольджи, которые продуцируют большое количество секоета. Секре ия слизи, кислот и гидро-
Столбчатые (А) и сидячие (Б, В) железы листа росянки
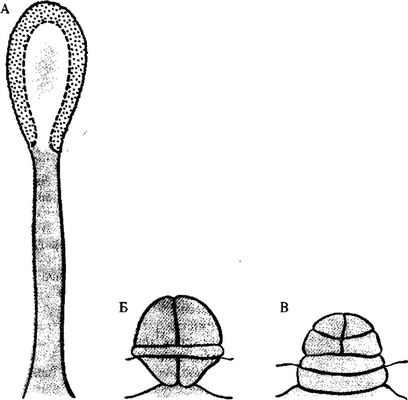
лаз носит активный характер и зависит от дыхания. Дыхательные яды — арсенит, цианид, а также хлороформ прекращают процесс секреции и продуцирование пузырьков с секретом в аппарате Гольджи.
Всасывание продуктов распада осуществляется теми же желёзками, соединенными с проводящей системой, и происходит довольно быстро, что было показано на примере мухоловки с помощью метиленовой сини. Через 5 мин краска проникала в цитоплазму всех клеток головки желёзок. Количественный ультраструктурный анализ показал доминирующую роль сим-пласта в транспорте продуктов пищеварения у мухоловки. Из всего сказанного следует, что процесс пищеварения у насекомоядных растений осуществляется принципиально так же, как в желудке животных. В том и другом случае секретируются кислоты (НС1 — в желудке, муравьиная кислота — у насекомоядных растений). Кислая реакция пищеварительного сока уже сама по себе способствует перевариванию животной пищи, так как в ее клетках активируются кислые лизосомные гидролазы. Выделение секреторными пищеварительными желёзками гидролаз (протеаз, фосфатаз, эстераз и др.), имеющих оптимум активности в кислой среде, создает благоприятные условия для переваривания пищи. На принципиальное сходство процесса кислого внеклеточного пищеварения у животных и растений впервые указал Дарвин в книге «Насекомоядные растения»
6. 2, 5. Приспособления растений к недостатку минеральных элементов
В ответ на дефицит нутриентов у растения интенсифицируется рост корней и увеличивается их количество. Это установлено при фосфорном и железном голодании растений фикуса и люпина. Как известно, корни обладают положительным гидро- и хемотропизмом. Если посадить растение в инертный субстрат и в определенное место положить кристалл питательной соли, то корни растения будут расти в направлении к этому кристаллу, что указывает на положительный хемотропизм. В то же время если вместо нутриента поместить токсичное вещество, корень будет расти в противоположном направлении. Это пример отрицательного хемотропизма.
На корнях в зоне поглощения образуется много корневых волосков. В мембранах клеток ризодермы увеличивается число систем транспорта ионов: ионных насосов, белков-переносчиков и каналов и повышается их активность, т. е. поглощение ионов регулируется и на уровне генной экспрессии. Соединения фосфора и железа хуже всего растворяются в щелочной среде. Именно на щелочных карбонатных почвах растения испытывают сильный дефицит фосфора и особенно железа. Поэтому выделение протонов ризодермой способствует переводу этих и других соединений в более доступные для растения формы. Помимо протонов ризодерма выделяет различные анионы, в основном анионы органических кислот. Последние вытесняют анионы минеральных кислот из почвенного поглощающего комплекса, способствуя закислению среды, а также принимают активное участие в хелатировании трудно растворимых или проч-носвязанных металлов. Кроме того, растворимость солей некоторых металлов зависит от степени окисления металла. Так, в почве содержатся главным образом плохо растворимые соли трехвалентного железа, тогда как соли двухвалентного железа лучше растворяются в воде. Растения поглощают железо на основе хелатирования и восстановления Fe3+-»Fe2+. Этот процесс хорошо изучен у цветковых растений. Существует две стратегии поглощения железа.
Стратегия I характерна для всех двудольных и однодольных, кроме злаков и, возможно, ситниковых и осок У растений при дефиците железа замедляются рост и утолщение корня, образуется очень много корневых волосков. Клетки ризодермы формируют лабиринтообразные инвагинации в клеточную стешу, как у клеток-спутников во флоэме или у секреторных клеток в солевых железках. Центральная вакуоль в этих клетках исчезает, формируется много маленьких вакуолей и увеличивается количество митохондрий. В мембранах клеток ризодермы возрастает количество белков-переносчиков, протонных помп, а также комплексов НАДФ-оксидоредуктазы. При поглощении железа активируется выделение протонов в ризосферу, что способствует увеличению растворимости железа. Стимулируется также секреция хелаторов, в первую очередь лимонной и яблочной кислот. Эти соединения хелатируют железо из почвенного поглощающего комплекса и проникают обратно в клеточную стенку. Далее на внешней стороне плазмалеммы Fe3+ восстанавливается НАДФ-редуктазой в Fe2+, поступает в клетку, а хелатор высвобождается обратно в почвенный раствор. Этот способ мобишзации железа достаточно эффективен, но неспецифичен, так как анионы органических кислот хелатируют не только железо, но и другие поглощаемые корнем металлы. К хе-латорам, выделяемым растениями и способствующим поглощению ионов железа, относится и рибофлавин. В форме хелатов двухвалентные ионы железа хорошо растворимы и поглощаются растением.
Стратегия II обнаружена у злаков. Корни злаков выделяют специализированные хелаторы— фитосидерофоры. Это непротеиногенные полиаминокислоты, которые синтезируются из метионина. Вначале образуется никотинамин, конденсация двух молекул никотинамина дает фигосидерофор — дезоксимугинеевую кислоту, из нее образуется му-гинеевая, или овсяная, кислота. Фитосидерофоры, хелатирующие железо из почвенных коллоидов, проявляют к нему достаточно высокое сродство, кроме него они образуют комплексы с Cu2+, Zn2+ и Со2+. С другими катионами: Са2+, Мg2+ А13+ и др., фитосидерофоры комплексов не формируют. После образования хелата он поглощается корнем. Переносчик проявляет активность преимущественно к комплексу Fe3+-фигосидерофор. Интенсивность поглощения комплексов с медью, цинком и кобальтом невелика. Попав в корень, трехвалентное железо в комплексе с фитосидерофором транспортируется в побег, где восстанавливается и включается в обмен. Синтез фитосидерофоров запускается только при недостатке железа. Стратегия II позволяет злакам достаточно легко, без затрат энергии на восстановление и в кратчайшие сроки ликвидировать дефицит железа в тканях.
Известен еще один способ решения проблемы дефицита элементов питания— секреция в ризосферу ферментов. Для минерального питания наиболее важны нитратредуктаза, участвующая в восстановлении нитрата, и фосфатаза, которая в кислой среде гидролизует нерастворимые полифосфаты почвы, переводя их в активное состояние.
В качестве приспособления к недостатку минеральных элементов следует рассматривать и создание запасов на «черный день», своеобразный вариант стратегии избежания стрессов. Азот откладывается в виде амидов и запасных белков. Кроме того, при сильном голодании возможна и мобилизация азота из вторичных азотсодержащих метаболитов, например алкалоидов. Фосфор запасается в виде конденсированных фосфатов, фитина, фосфо- и нуклеопротеидов. Фитин служит удобной запасной формой и для кальция и магния. Железо и микроэлементы «хранятся» обычно в комплексе с белками. Так, белок ферритин, имеющий много гидроксигрупп, часть из которых фосфорилирована, способен связывать до 6000 атомов железа. Ферритин представляет собой белковый цилиндр, внутри которого находится гидроксид железа. Это позволяет клетке накапливать большие концентрации металла, «не опасаясь» его токсического действия. Микроэлементы запасаются с помощью белков-металлотионеинов. Таким образом, эти белки участвуют не только в инактивации тяжелых металлов, но и в депонировании минеральных веществ.
Растения — автотрофные организмы, в ходе фотосинтеза и почвенного питания они поглощают неорганические вещества и синтезируют из них органические. Немногие растения (сапрофиты, паразиты и насекомоядные) относятся также к факультативным или облигатным гетеро-трофам, получающим органические вещества из внешней среды. Сапро-трофы (сапрофиты) питаются органическими веществами разлагающихся остатков растений и животных, паразиты — органическими веществами живых существ. Большинство высших растений имеет ряд приспособлений к использованию не только минеральных, но и органических субстратов, т. е. к гетеротрофному питанию, например, при прорастании семян, клубней, луковиц и др. При выращивании растений на органических удобрениях даже в стерильных условиях их продуктивность может быть выше, чем на эквивалентном по элементному составу минеральном субстрате. Объясняется это не только необходимостью энергетических трат на восстановление и включение веществ в органические соединения, но и другими причинами. Так, гуминовые кислоты — слабые поверхностно-активные вещества, т. е. детергенты, большей частью способны проникать через плазмалемму. По представлениям некоторых специалистов, гуминовые кислоты могут поступать почти целиком внутрь растительной клетки, перенося много различных элементов: металлы, органический азот, фосфор и серу. Кроме того, эти вещества, имеющие углеродный скелет, могут снова вовлекаться в обмен. В экспериментах по применению искусственных поверхностно-активных веществ повышается усвоение и органических, и минеральных удобрений. Растения могут поглощать мономерные органические вещества: сахара, некоторые аминокислоты, витамины, из почвы. В частности, витамин В12, который не синтезируется растениями, они поглощают из почвы, где его образуют бактерии.
В большинстве случаев высшие растения, оставаясь зелеными ав-тотрофными организмами, получают дополнительное азотное питание благодаря симбиотическим отношениям с трибами и/или бактериями, которые поселяются в ризосфере или непосредственно в корнях. И наконец, хищные или насекомоядные растения, обитающие обычно в условиях крайнего дефицита минеральных веществ: на болотах или в пресных водоемах, проблему дефицита азота и фосфора или органических соединений решают, поедая насекомых, ракообразных, инфузорий и др. Таким образом, проблема дефицита элементов минерального питания решается растениями самыми разнообразными способами: от усиления воздействия на почву до образования ассоциаций с другими организмами и даже перехода на гетеротрофный способ питания.
Экологические условия произрастания растений на болотах
Своеобразные едафични условия для роста растений образовались на болотах, где в качестве почвенного субстрата служит торф.
Торф (от араб, tubar - земля) - продукт неполного разложения отмерших болотных растений в условиях избыточного увлажнения и недостаточного доступа воздуха, который по мере наслоения значительной толще или захоронения наносами приобретает характер горной органогенного породы.
В регионах с влажным климатом, где осадки преобладают над испарением, часто проявляется процесс заболачивания, особенно там, где есть первичные условия - своеобразный рельеф и почвы. Избыток влаги, является основной чертой болота, вызывает появление особой болотной растительности и специфического почвенного процесса - торфоутворення. Типичными для Украинского Полесья является сфагновые болота. Водный режим этих болот обусловливается не только избытком влаги, а и своеобразием самых сфагновых мхов. Высокая влагоемкость свойственна как живой, так и мертвой сфагнума. В торфяных болотах низкий уровень бокового почвенного стока, значительное количество органики, имеющиеся токсичны для растений сероводород и метан. Еще одна особенность торфа - низкая теплопроводность, вследствие чего субстрат, где размещаются корни, имеет температуру на 2 - 4 ° С ниже, чем обычный грунт. Кроме того, постоянный прирост торфа поспособствовал тому, что в части растений выработалась способность образовывать новые корни по мере их захоронения.
Из-за накопления гуминовых кислот для болот характерна высокая кислотность субстрата как известно, способствует вымыванию минеральных веществ. Зольность торфа верховых болот составляет всего 2-4%. Некоторые болота вообще удовлетворяются только минеральными веществами, поступающими с осадками. В условиях дефицита элементов минерального питания растения характеризуются низкорослостью. Не только деревья, но и кустарники и кустарнички имеют хорошо развитую микоризу (береза, сосна, клюква, багульник, голубику, черника и т.д.). Симбиоз с грибами обеспечивает дополнительное количество элементов питания. Считается, что именно бедность сфагновых болот минеральными веществами объясняет появление на них насекомоядных растений - росянки круглолистной (Drosera rotundifolia), толстянки обычной (Pingvicula vulgaris ), волдырь обычного (Utricularia vulgaris). Таким образом, на болотах в условиях избыточной увлажненности и нехватки элементов минерального питания, формируются специфические фитоценоз.
Обмен минеральных веществ в наземных экосистемах
Поглощение питательных веществ растениями происходит за счет фотосинтеза, распада органических веществ, и, частично, за счет минералов, которые поступают из материнской породы. Между растительными группировками и грунтом существует постоянный круговорот минеральных веществ, в природных условиях имеет почти замкнутый характер (рис. 6.7). Поглощены и усвоены растениями химические элементы или непосредственно в виде растительных остатков, или через трофические цепи возвращаются в почву в составе органических веществ. Они образуют резерв питательных веществ. Органические остатки, в свою очередь, минерализуются и сорбируются почвенными коллоидами. Не все питательные вещества, необходимые для роста и сохранения растительных тканей, поступающих из почвы. Значительная их часть циркулирует в пределах самого растения. Минерализация и синтез органических веществ должны происходить синхронно, поэтому, скажем, при медленном распаде лесной подстилки чаще всего происходит и замедление скорости роста растений. В то же время, при интенсивной минерализации органических остатков, химические элементы быстро высвобождаются и снова включаются в круговорот.
Значительная производительность тропических лесов объясняется именно тем, что в таких экосистемах очень интенсивный круговорот минеральных веществ. Это обуславливает несколькими составляющим: высокой влажности воздуха и почвы, постоянно высокой температуре воздуха, интенсивной деятельности микроорганизмов, которые как раз и принимают основное участие в минерализации органики благодаря, опять же, благоприятным климатическим условиям. Именно последние являются причиной того, что круговорот минеральных веществ в экосистемах дождевых тропических лесов происходит непрерывно и практически с одинаковой скоростью
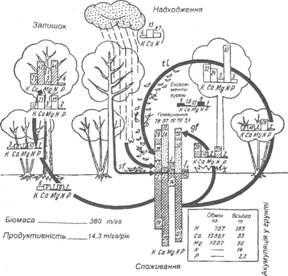
Рис. 6.7. Годовой круговорот основных химических элементов в смешанном дубово-грабово- буковом лесу в Бельгии (кг / га)
(По Р. Duvigneaud. 1968)
Остаток - в ежегодном приросте корней, древесины, коры однолетних побегов.
Возвращение - через лесную подстилку, наземную флору, вымывания, выщелачивания и сток по стволам.
Поступления - за счет осадков (случайные учтены).
В хвойных лесах умеренной зоны, особенно в отношении сухих и влажных условиях роста, происходит накопление органической массы в лесной подстилке и, таким образом, наблюдается изъятия (консервирования) части минеральных веществ из круговорота. Это объясняется наличием в осадках органических кислот и смолистых веществ, которые замедляют процессы минерализации, поскольку создаются неблагоприятные условия для жизнедеятельности микроорганизмов. Кроме того, благодаря наличию неблагоприятных сезонов и значительным колебаниям погодных условий в течение вегетационного периода, круговорот минеральных веществ несколько замедленный и периодический.
Любому растению необходимы не просто тепло, влажность, минеральные вещества или любые другие факторы, а их определенное количество или определенный режим. В 1840 немецкий химик Юстас фон Либих сформировал правило, известное как закон Либиха. В соответствии с законом, веществом, которое находится в минимуме, управляется урожай и определяется устойчивость последнего во времени. Либих сформулировал его по отношению к сельскохозяйственным культурам. Но оказалось, что эта закономерность имеет глобальный характер и может быть применена к любым группировок. Универсальное правило, сформулированное в 1909 Е. А. Мичерлихом, является уточнением закона Либиха:
Производительность биологической системы определяется всей совокупностью действующих экологических факторов.
Это одно из важнейших обобщений экологии. Выводом из закона совокупного действия факторов является то, что ограничение дозы (или отсутствие) любого вещества, необходимого для живого организма, приводит к одинаковому результату - замедлению роста и развития. Так, например, величина урожая определяется количеством в почве того элемента питания, необходимость растения в котором довольна меньше. Предложенное Е. А. Мичерлихом и Б. баул математическую формулировку закона совокупного действия стало первой попыткой математического описания явления взаимодействия экологических факторов. Именно эта работа стала базой для многофакторного анализа экосистем.
Хищные растения

Насекомоядные растения (используются также термины хищные растения и плотоядные растения) — представители растений, адаптировавшиеся для ловли и переваривания небольших животных, в основном насекомых. Таким образом они дополняют свое нормальное автотрофное питание (фотосинтез) одной из форм гетеротрофного питания.
Описание
Обычно такие растения обитают в обедненных азотом местах, а животных используют как дополнительный источник азота. Привлекая насекомых окраской, запахом или сладкими выделениями, растения ловят их тем или иным способом, а затем высвобождают в ловушку ферменты, которые переваривают пойманную жертву. Образующиеся в результате такого внеклеточного пищеварения продукты, в основном аминокислоты, всасываются и усваиваются.
У одних хищных растений (росянка, жирянка, росолист и др.) листья покрыты многочисленными желёзками, выделяющими липкую прозрачную жидкость, привлекающую насекомых и приклеивающую их к листу. При попадании насекомого на ловушку у растения усиливаются выделения из желёзок; при этом железистые волоски изгибаются по направлению к телу насекомого (росянка) или заворачиваются края ловчего листа, на котором оно находится (жирянка). У других растений ловчий аппарат представлен или пассивно улавливающими насекомых урнами (непентес, саррацения, дарлингтония и др.) или активно действующими ловушками (дионея, альдрованда, пузырчатка и др.).
Росянка — один из редких представителей подобных растений, произрастающих в Европе, поскольку большинство из них обитает в тропиках или субтропиках. Эти растения предпочитают влажные степи и болота, по большей части с кислыми почвами, с дефицитом минеральных веществ. Жирянка произрастает в лесотундре.
См. также
Ссылки
Wikimedia Foundation . 2010 .
Полезное
Смотреть что такое "Хищные растения" в других словарях:
хищные растения — Синонимы: плотоядные растения растения, восполняющие недостаток азота ловлей мелких беспозвоночных при помощи видоизмененных листьев, напр. росянка (Drosera), непентес (Nepenthes) … Анатомия и морфология растений
Растения — Многоо … Википедия
Растения-хищники — Непентес Насекомоядные растения (используются также термины хищные растения и плотоядные растения) представители растений, адаптировавшиеся для ловли и переваривания небольших животных, в основном насекомых. Таким образом они дополняют свое… … Википедия
Хищные животные — Хищник животное или другой организм (например, хищные растения), основной частью рациона которого являются живые животные Плотоядные животные, питающиеся преимущественно мясом, в том числе падалью Хищничество форма биологического… … Википедия
РАСТЕНИЯ НАСЕКОМОЯДНЫЕ — растения хищные, экологическая группа автотрофных растений (около 500 видов), способных улавливать насекомых и частично переваривать их с помощью протеолитических ферментов и органических кислот, в результате чего они восполняют недостаток азота… … Экологический словарь
Хищные грибы — Полифилетическая группа грибов Ловчая сеть грибка рода … Википедия
Насекомоядные растения — Полифилетическая группа растений Росянка анг … Википедия
Плотоядные растения — Непентес Насекомоядные растения (используются также термины хищные растения и плотоядные растения) представители растений, адаптировавшиеся для ловли и переваривания небольших животных, в основном насекомых. Таким образом они дополняют свое… … Википедия
плотоядные растения — см. хищные растения … Анатомия и морфология растений
Растение — ? Растения Многообразие растений Научная классификация Надцарство: Эукариоты Царство: Растения … Википедия
Ловить или не ловить

Насекомоядные растения обычно живут в местах, где мало элементов минерального питания. Показано, что они часто плохо растут и дают меньше семян, если лишить их возможности ловить жертвы. Но насекомые нужны им и для опыления цветков. Если слишком много насекомых будут привлечены цветками, растения поймают меньше жертв в свои ловушки и получат недостаточно минеральных веществ, а если почти все опылители будут пойманы, то может снизиться успешность размножения. Какие приспособления насекомоядных растений могут помогать им хотя бы частично разрешить это противоречие?
Подсказка 1
Подумайте, как растение может привлекать опылителей и жертв и в каких случаях оно может без них обойтись.
Подсказка 2
Поищите фотографии разных видов цветущих насекомоядных растений — это может помочь найти некоторые решения проблемы.
Решение
Попробуем логически вывести возможные способы разрешения этого конфликта, а заодно посмотрим, пользуются ли ими реальные хищные растения.
Пожалуй, самый радикальный способ — вообще отказаться от семенного размножения. Среди насекомоядных растений есть виды, способные к вегетативному размножению, но их не так уж много; и подавляющее большинство имеют цветки и дают семена. Так что такой радикализм в целом хищным растениям не присущ.
Второй по степени радикальности вариант — отказаться от перекрестного опыления, заменив его облигатным (или хотя бы факультативным) самоопылением или апомиксисом. Процент таких видов среди хищных растений довольно высок — видимо, в некоторых семействах он выше, чем среди «нехищных» родственников. Однако многие другие насекомоядные растения нуждаются в перекрестном опылении, и среди них есть даже такие виды, у которых самоопыление невозможно.
Третий вариант — перекрестное опыление ветром. Как ни странно, среди хищных растений этот вариант, кажется, не встречается. Почему-то насекомоядность возникла только у представителей энтомофильных семейств, и «отказаться» от связей с насекомыми им не удалось.
Даже удивительно, что не все хищные растения пошли этими легкими путями. Ведь при этом можно было бы дополнительно сэкономить энергию на производство нектара, запаха и привлекательных лепестков. Но факт остается фактом: очень многие хищные растения все-таки нуждаются в насекомых-опылителях. В таком случае было бы действительно глуповато переловить и переварить всех собственных опылителей. Как же этого избежать? Первая заповедь хищного растения — не делать ловушек из цветков!
Удержаться, конечно, сложно — ведь цветки так хорошо привлекают насекомых. И действительно, «цветочный» запах испускают ловушки по крайней мере одного вида непентесов (Nepenthes rafflesiana, см. The leaves of carnivorous plants emit flower scents to attract their prey) и одного вида австралийских росянок. В эти ловушки попадается много летающих насекомых, и для непентеса доказано, что их привлекает именно запах. А вот про настоящие цветки этих растений и про то, как они привлекают своих опылителей, увы, мало что известно. Иногда ловушки также выделяют нектар и имеют контрастные ультрафиолетовые «метки», характерные для цветков.

Ловушки хищных растений — это, как правило, видоизмененные листья или их части. А раз так — от цветков их можно изолировать. Вариантов такой изоляции два — в пространстве и во времени. Хищные растения используют оба варианта. Надежнее всего пространственная изоляция у растений, ловушки которых находятся под водой или под землей. Подводные ловушки пузырчатки (Utricularia) и альдрованды (Aldrovanda) ловят всякую мелкую водяную живность — от протистов и мелких рачков до мальков рыбок, — а цветки их находятся высоко над водой и опыляются насекомыми (рис. 1).
Подземные ловушки встречаются у пузырчатки (оказывается, больше половины видов этого самого крупного рода хищных растений — наземные или полуназемные!), а также у представителей рода Genlisea и не так давно открытого рода Philcoxia (см. Katherine Rowland, 2012. Hungry plant traps worms underground). Ловят они почвенных протистов и нематод, и опылителям этих растений тоже причинить вред не могут.
Сложнее обстоит дело с пространственной изоляцией у росянок (Drosera). При изучении большой выборки видов (более 50) выяснилось, что у низеньких росянок с распростертыми на земле ловчими листьями расстояние между листьями и цветками больше, чем у более высоких и имеющих ловчие листья на стеблях. Если бы это расстояние помогало защищать опылителей — следовало бы скорее ожидать обратной картины.

В другой работе при сравнении двух видов (с короткой и длинной цветоносной стрелкой) оказалось, что оба они ловят очень мало опылителей. Но если у вида с длинной стрелкой ее искусственно укоротить, то гораздо меньше насекомых посещают цветки. Значит, решили авторы работ, длинные цветоносы возникли не для «спасения» опылителей, а просто для того, чтобы поднять цветы повыше и сделать их позаметнее для насекомых (рис. 2).
Но в третьей работе в результате довольно сложных опытов было показано, что по крайней мере для некоторых видов росянок длина имеет значение: опылители предпочитали белый цвет (цветки) красному (ловушки) только при заметной разнице между ними по высоте (см. детали в статье Натальи Резник «Сохранить опылителей»).
Временная изоляция тоже встречается: у саррацений (Sarracenia) цветки не только сидят на длинных цветоножках, но и распускаются ранней весной, обычно до появления новых ловчих листьев.
Дальше могут включаться более сложные и тонкие механизмы. Можно с помощью разных ухищрений ловить одних насекомых, а «использовать» для опыления других. Немногочисленные пока работы показывают, что состав жертв и опылителей действительно сильно различается у многих хищных растений. Самый простой случай — когда в расположенные у земли ловушки попадают в основном ползающие или прыгающие насекомые, а опылением занимаются хорошие летуны. Можно ловить только маленьких и хилых комариков-хирономид или трипсов, а к опылению привлекать крупных и мощных пчел или мух, а то и птиц (цветки некоторых росянок птицы действительно посещают, но их участие в опылении не доказано). Такие крупные опылители могут не попадаться в мелкие ловушки или легко освобождаться из них.
Наконец, можно специально «указывать дорогу» — жертвам в ловушки, а опылителям к цветкам. Такими указателями у растений чаще всего служит цвет (включая ульрафиолетовые «метки») или запах. Запах цветков может привлекать одних насекомых (действительно полезных как опылителей), а запах ловушек — других. И такой случай (пока чуть ли не единственный) обнаружен у одного из видов росянок. У этого вида и цветки, и ловушки пахнут по-разному и, действительно, привлекают разных насекомых (см. статью Натальи Резник «Сохранить опылителей»). В той же работе про росянок, которую упоминает Наталья Резник, показано, что опылители выбирают белый цвет, предпочитая его красному, если между ними есть расстояние по вертикали (A. M. El-Sayed et al., 2016. Pollinator-prey conflicts in carnivorous plants: when flower and trap properties mean life or death). Жертвы тех же видов росянок таких предпочтений не выказали. В другой работе было доказано, что красные ловушки непентеса ловят насекомых эффективнее, чем зеленые. Видимо, не случайно ловчие листья многих растений имеют красную окраску, а у некоторых краснеют при дефиците азота. Цветки таких растений обычно белые, и не исключено, что их опылители «научились» выбирать «некрасные» посадочные площадки.
Послесловие
Конечно, решая такую задачу, составитель рассуждает, исходя из известных ему фактов, и таким образом ограничивает свою фантазию. Например, можно предположить, что у каких-то растений ловушки не просто непривлекательны для опылителей, а отпугивают их противным для них запахом или «опасным» цветом. Кстати говоря, известно, что в некоторых случаях красный цвет листьев насекомых не привлекает, а, скорее, отпугивает (см. Marco Archetti et al., 2009. Unravelling the evolution of autumn colours: an interdisciplinary approach).
А если пофантазировать, что растения могут использовать насекомых-помощников, отпугивающих от ловушек опылителей и заманивающих туда жертв? Слишком сложно? Вряд ли. В этом, как и в еще более сложных вариантах решения, нет ничего невероятного. По крайней мере, насекомые-помощники у разных хищных растений имеются. У одного из видов непентесов на ловушках и рядом с ними живут муравьи-кампонотусы, которые не отпугивают большинство насекомых (как часто делают муравьи-мутуалисты других растений), а поджидают добычу, разрывают ее на части и частично съедают, а частично «кормят» останками жертв растение-хозяина. На ловчих листьях другого хищного растения живут клопы, которые сами к ловчим листьям не липнут (как паук к своей паутине), а попавшихся жертв высасывают. Растение питается их экскрементами — как и в первом случае, у растения-хозяина слабоваты собственные пищеварительные ферменты. Так что, как выясняется, на компромисс хищным растениям приходится идти не только с опылителями, но и с другими насекомыми-помощниками. Может быть, и для нас это неплохо. Откажись растения от подобных компромиссов — и кто знает, не пришлось ли бы нам опасаться не только медведей и акул, но и триффидов (Triffid).
В реальности избежать компромиссов сложно, особенно если ты сочетаешь хищничество с растительным образом жизни. Конфликт «жертва — опылитель» у хищных растений (pollinator-prey conflict, PPC) — лишь один из типичных примеров адаптивного компромисса, или trade-off. Невозможность оптимизировать живой организм одновременно по всем параметрам неизбежно приводит к компромиссным решениям. Но существует ли PPC на самом деле? В своем обзоре Андреас Юргенс (см. список источников в конце послесловия) перечисляет условия, при которых PPC может существовать. Для этого успех размножения должен зависеть от опылителей, между составом опылителей и жертв есть перекрывание, а опылителей (или пыльцы) должно хотя бы иногда не хватать.
Установить, когда все эти условия выполняются, очень непросто. Хищные растения часто живут в достаточно труднодоступных и неудобных для наблюдения местах. (Когда-то много лет назад мои ученики вели круглосуточные наблюдения за несколькими видами росянок (по результатам этих наблюдений было опубликовано несколько статей. Сам я в этих наблюдениях не участвовал, а только возил наблюдателей на болото, но я хорошо представляю, что значит по нескольку часов кряду лежать на доске среди сплавины в окружении слепней и комаров.)
А без длительных наблюдений в природе невозможно определить ни состав опылителей, ни даже состав жертв (полупереваренных жертв с собранных растений в лаборатории редко когда удается определить точнее, чем до семейства).
Поэтому лишь в немногих случаях удается доказать, что лучшие растения-ловцы в пределах вида действительно могут пострадать от нехватки опылителей и дать меньше семян. Четко установить это, кажется, пока удалось только для одного из видов жирянок (Pinguicola).
Есть у РРС и дополнительные аспекты, в том числе близкие к «войне полов» (см. Sexual conflict) — они обсуждаются в том же обзоре Юргенса. Допустим, все насекомые прилетают на цветок с другого цветка такого же вида, все вымазаны в пыльце и оставляют ее на рыльце пестика. Если насекомое прилетает на цветок, принося пыльцу с другого растения, а потом попадает в ловушку, то выигрывает женская репродуктивная функция, а растение получает дополнительную пищу (само насекомое и пыльцу, в которой вымазана жертва). Дополнительная пища, возможно, позволит образовать больше семян. Но зато в этом случае пыльца (и мужские гаметы) на другое растение не попадут — мужская репродуктивная функция пострадает. А если насекомое измажется в пыльце и перелетит на цветок другого растения, то и женская функция (опыление своего пестика), и мужская (доставка своей пыльцы на чужой пестик) осуществятся, но зато растение не получит дополнительной пищи — а это плохо для женской репродукции.
Как этот очередной «конфликт интересов» разрешается в природе, никто даже не начинал изучать. Да и кто даст на это деньги? Уж тут-то наверняка это просто «удовлетворение собственного любопытства за казенный счет».
В целом это, конечно, так и есть (если не считать, что выращивание и продажа насекомоядных растений — модный и быстро развивающийся сегмент рынка комнатного цветоводства). Но если насекомоядные растения мало кого волнуют (хотя и они могут стать источником каких-нибудь ценных лекарств), то насекомые не могут нас не волновать. Значит, на изучение стимулов. управляющих их поведением, деньги могут быть потрачены и не впустую. А иногда хищные растения навевают ученым и вовсе неожиданные идеи и решения. Вот, например, росянка навеяла группе ученых из Университета штата Пенсильвания идею создания двуслойного синтетического биоматериала, который сначала отлавливает из крови, а затем убивает раковые клетки (Sh. Li et al., 2015. A Drosera-bioinspired hydrogel for catching and killing cancer cells). Может быть, и изучение РРС что-нибудь полезное может навеять — ведь никогда заранее не знаешь, что этим ученым придет в голову.
Спасибо П. А. Волковой за идею задачи.
Основные источники:
1) Carnivorous Plants. Physiology, ecology, and evolution // Oxford Univ. Press, 2017. Edited by Aaron Ellison and Lubomír Adamec. Adam T. Cross, Arthur R. Davis, Andreas Fleischmann, John D. Horner, Andreas Jürgens, David J. Merritt, Gillian L. Murza, and Shane R. Turner. Chapter 22: Reproductive biology and prey-pollinator conflicts.
2) Andreas Jürgens, Amber Sciligo, Taina Witt, Ashraf M. El-Sayed, D. Max Suckling. Pollinator-prey conflict in carnivorous plants // Biol Rev Camb Philos Soc. 2012. V. 87(3). P. 602—615.
3) Ashraf M. El-Sayed, John A. Byers, and David M. Suckling. Pollinator-prey conflicts in carnivorous plants: When flower and trap properties mean life or death // Sci Rep. 2016. V. 6: 21065. Published online 2016 Feb 18.
4) Наталья Резник. Сохранить опылителей // «Троицкий вариант — Наука» № 249, 13.03.2018, c. 16.
Читайте также:
- Диагностика менингиомы мостомозжечкового угла и внутреннего слухового прохода по КТ, МРТ
- Диагностика гнойных и септических осложнений ранений шеи, груди и живота
- Лучевые изменения брюшной полости при обструкции верхней полой вены (ВПВ)
- Трансплантация почки ребенку. Актуальность проблемы
- Гипоплазия и агенезия клеток Лейдига. Причины и диагностика