Двигательные рефлексы глаз. Взаимосвязь мозжечка и вестибулярного аппарата
Добавил пользователь Alex Обновлено: 28.01.2026
ЛОР-болезни:
Популярные разделы сайта:
Вестибулоглазодвигательные связи. Механизмы нистагма
Вестибулярные ядра имеют тесную связь с глазодвигательными ядрами. Главная связь осуществляется через систему перекрещенных и неперекрещеппых связей, формирующих задний продольный пучок [Клосовский Б. Н., Gehuchten P., Muskens L.]. От медиального ядра идут коллатерали к ядру отводящего нерва той же стороны. Основные связи вестибулярных ядер с III нервом перекрещенные, они более многочисленны, чем неперекрещенные.
Указывают, что не существует строго функционального-разграничения вестибулярных ядер, так как повреждение одного из них вызывает одновременное повреждение другого вследствие их близкого расположения. По этому вопросу даже в специальной зарубежной анатомической литературе приводятся очень скуднее и не совсем определенные даные [Бродал А., Вальберг Ф., Помпеано О.]. В то же время Не отражены важные сведения, полученные советскими учеными при разработке данного вопроса. Так, классические исследования Б. Н. Клосовского, Б. Н. Клосовского и Е. Н. Космарской внесли большой вклад в изучение вестибулоокуломоторных путей, функции вестибулярных ядер, механизма нистагма и произвольных движений глаз.
Чтобы избежать повреждения вестибулярных ядер при исследовании их функции, изолированно перерезали пути, идущие от вестибулярных ядер к ядру глазодвигательного нерва, таким образом впервые удалось достоверно выяснить функции медиального и верхнего ядер. Эти исследования показали, что горизонтальный нистагм осуществляется в медиальных ядрах, откуда импульсы идут к глазодвигательным ядрам по перекрещенным путям. Вертикальный нистагм осуществляется в верхних ядрах, откуда импульсы идут к глазодвигательным ядрам по неперекрещенным путям. Каждый из видов нистагма (горизонтальный, вертикальный, ротаторный) имеет свои проводящие пути.
Анатомический путь для горизонтального нистагма следующий: нейроэпителий ампулы горизонтального (латерального) полукружного капала, вестибулярный узел, вестибулярный нерв, медиальное ядро, перекрещенная вестибулоокуломотор-ная система той же стороны, ядро глазодвигательного нерва противоположной стороны, прямой нейрон от III нерва к внутренней прямой мышце глаза. Реципрокное торможение совершается через комиссуральные волокна между медиальными ядрами.
Анатомический путь для вертикального нистагма: нейроэпителий ампул вертикального канала, вестибулярный узел, вестибулярный нерв, верхнее ядро, неперекрещенный вестибулоглазодвигательный путь, ядро глазодвигательного нерва, перекрещенные и прямые пути к верхней и нижней мышцам глаза. Связь между обоими верхними ядрами осуществляется за счет комиссуры Фузе.
Пути для ротаторного нистагма еще не выявлены, но и для этого вида нистагма должны существовать особые пути в ЦНС.
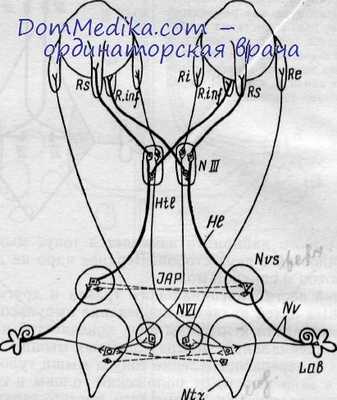
Пути вестибулярных импульсов при горизонтальном (тонкие линии) и при вертикальном (толстые линии) нистагме (по Б. Н. Клосовскому).
Lab — лабиринт; Nv — вестибулярный нерв; Ntr — медиальное ядро; Nvs — верхнее ядро; NVI — ядро VI нерва; NIII — ядро III нерва; HI — неперекрещенный (гомолатеральный) пучок; Htl — перекрещенный гетеролатеральный пучок; Re — наружная прямая мышца глаза; Ri — внутренняя прямая мышца глаза; Rs — верхняя прямая мышца глаза; R. inf — нижняя прямая мышца глаза; JАР — комиссу-ральные волокна между верхними вестибулярными ядрами.
А. Бродал, Ф. Вальберг, О. Помпеано, R. W. Bacon, V. Honubia считают, что вестибулоокуломоторный путь осуществляется как через медиальные продольные пучки, так и через сетчатое образование.
Некоторые авторы указывают на существование в мозге кролика мезодиэнцефалической области, электростимуляцией которой можно вызвать центральный нистагм. А. Е. Курашвили, В. Е. Корюкин при стимуляции передней подбугорной области в эксперименте наблюдали контралатеральный нистагм. По данным других авторов, стимуляция промежуточного мозга не дает эффекта, в то время как раздражение среднего мозга в области глазодвигательного ядра, медиального продольного пучка и примыкающего к этим структурам сетчатого образования вызывало максимальные ответы в вестибулярных ядрах.
По нашим данным, диэнцефально-гипоталамическое поражение обычно протекает с выраженным торможением экспериментального нистагма при повышении вегетативных реакций. Спонтанного нистагма при изолированном диэнцефальном поражении мы никогда не наблюдали.
На основании экспериментальных данных Б. Н. Клосовский установил, что корковые импульсы для сочетанных движений глаз идут через вестибулярные ядра, а оттуда в восходящем направлении через задний продольный пучок. Связь коры с вестибулярными ядрами осуществляется перекрещенными путями, волокна от коры подходят, вероятно, к центру быстрой фазы нистагма. Вестибулярные ядра и их связи с глазодвигательными ядрами являются подкорковыми центрами для координированных сочетанных движений глаз в горизонтальной и вертикальной плоскостях.
На основании экспериментальных данных мы полагаем, что вестибулярные ядра, принимающие участие в вестибулоглазодвигательных реакциях, функционально тесно связаны с кортикальным производным центром взора (поле 8) и со зрительной корой затылочной доли мозга. Эфферентные кортикальные влияния на эти же вестибулярные ядра оказывают височно-теменно-лобные отделы мозга обоих полушарий, так как при поражении этих областей четко выявляется асимметрия вестибулярного экспериментального нистагма по стороне независимо от раздражаемого лабиринта.
Посредством вестибулоглазодвигательпых связей, проходящих в заднем продольном пучке, вестибулярные импульсы влияют на глазные мышцы. Этим же путем происходят рефлекторные сочетания отклонения глаз, в результате которых направление взгляда не меняется при изменении положения головы. По этому же пути осуществляются нистагм и компенсирующая установка глаз. Задний продольный пучок координирует деятельность вестибулярного аппарата, глазных мышц, мозжечка и спинного мозга, то есть он соединяет центры, важные для сохранения равновесия тела и ориентировки в пространстве. Механизм нистагма тесно связан с механизмом движения глаз.
В клинической практике изучение патологии вестибулоглазодвигательпых реакций (нистагма) дает наиболее информативные и объективные данные из всего многообразного комлекса вестибулярных реакций.
Мозжечок и вестибулярная система (анатомия и физиология)
Мозжечковая атаксия развивается не только при поражении самого мозжечка, но и его проводников; как афферентных, так и эфферентных. На мозжечковую атаксию закрывание глаз не влияет, глубокая чувствительность остается сохранной.
Мозжечок расположен в задней черепной ямке над продолговатым мозгом и варолиевым мостом. Кверху от него находятся затылочные доли, между ними и мозжечком натянут мозжечковый намет - tentorium. Мозжечок состоит из среднего отдела или червя и двух полушарий. Поверхностным слоем мозжечка является кора - серое вещество. Кроме того, в белом веществе имеются еще скопления серого вещества - ядра мозжечка. Наиболее важными из них являются nucl. dentatus (зубчатое ядро), nucl. fastigi - ядро шатра.
С другими отделами центральной нервной системы мозжечок связан тремя парами ножек:
1) нижние ножки - corpora restiformia (веревчатые тела) (соединяют мозжечок с продолговатым мозгом);
2) средние ножки - brachia pontis (соединяют мозжечок с варолиевым мостом);
3) верхние ножки - brachia conjunktiva (соединяют мозжечок со средним мозгом на уровне четверохолмия).
Ножки мозжечка состоят из волокон, которые приносят импульсы к мозжечку или отводят от него.
Червь мозжечка является филогенетически более древним отделом связан с вестибулярным аппаратом.
Полушария мозжечка являются более новым образованием, развиваются параллельно с развитием коры головного мозга.
Мозжечок выполняет следующие функции:
ü автоматическая координация движений;
ü регуляция мышечного тонуса;
ü обеспечение равновесия тела.
В осуществлении произвольных движений главная роль мозжечка состоит в согласовании быстрых (фазических) и медленных (тонических) компонентов двигательного акта. Это возможно благодаря связям мозжечка с мышцами и корой головного мозга.
Мозжечок получает афферентные импульсы от всех рецепторов, раздражающихся во время движения - это проприорецепторы, вестибулярные, зрительные, слуховые, т.е. получает информацию о состоянии двигательного аппарата.
Сам мозжечок оказывает влияние на красное ядро и ретикулярную формацию ствола, которые посылают импульсы к γ-мотонейронам спинного мозга, регулирующим мышечный тонус. Импульсы от мозжечка поступают в кору двигательной зоны. Основная функция мозжечка осуществляется на подкорковом уровне: мозговой ствол, спинной мозг.
Эфферентные импульсы от ядер мозжечка регулируют проприоцептивные рефлексы на растяжение. При мышечном сокращении происходит возбуждение проприорецепторов (мышечных веретен) в мышцах агонистах и антагонистах, но рефлекса не возникает, так как мозжечок оказывает тормозное влияние. При поражении мозжечка это влияние утрачивается, и в конечностях при выполнении движений появляются колебательные движения (особенно при выполнении целенаправленных действий за счет возбуждения агонистов и антагонистов). Многие мозжечковые расстройства связаны с нарушением реципрокной иннервации антагонистов: при выполнении любого движения мышцы агонисты и антагонисты находятся в противоположном состоянии. Если нейроны сгибателей возбуждаются, то нейроны разгибателей тормозятся. Механизм состоит в реципрокном торможении спинномозговых двигательных центров. Этот механизм осуществляется сегментарным аппаратом спинного мозга при участии мозжечковых систем.
Вестибуло-зрительное взаимодействие и невесомость
С древнейших времён человечеству известно состояние укачивания - болезни движения (БД). В дневниках Аристотеля сохранились описания о неприятных ощущениях у воинов, передвигающихся на слонах в армии А. Македонского. Вначале это состояние связывали с длительным качанием головы и туловища, но вскоре эмпирическим путём человечество пришло к выводу, что причиной укачивания является ухо. С развитием транспорта, особенно мореплавания и авиации, стало очевидным, что причиной развития БД является вестибулярный аппарат. Резкое угнетение функции или отсутствие вестибулярного аппарата делает людей нечувствительными к БД. Подкреплением точки зрения о ведущей роли вестибулярной системы в развитии БД является резистентность к ней лиц с нефункционирующими лабиринтами в условиях параболического полета и при воздействии кориолисовых и прецессионных ускорений. Глухонемые люди не укачиваются. Установлено, что животные невосприимчивы к воздействию ускорений при разрушении вестибулярного аппарата и вестибулярных путей в центральной нервной системе (ЦНС).
Установлено, что вестибулярный аппарат, реагируя на ускорение, изменение положения тела и гравитацию, обеспечивает:
- ориентацию в пространстве;
- управление равновесием;
- стабилизацию изображения на сетчатке глаз;
- сохранение неподвижного изображения на сетчатке глаз при движении головы/тела.
Однако, для осуществления ориентации в пространстве, точности зрительного слежения (статических и динамических фиксационных саккад, удержания взора, плавного слежения) и координации тела необходимы как адекватное функционирование отдельно взятых сенсорных систем, так и их слаженное взаимодействие (конвергенция) в интегративных структурах ЦНС. Это касается прежде всего, зрительной, вестибулярной и двигательной систем, которые являются компонентами единой перцепто-моторной системы «организм-среда» и функционально неразделимы.
В условиях невесомости статолиты теряют вес и перестают давить на мембрану, и привычная для наземных условий информация об изменении положения головы и о линейном перемещении организма в ЦНС не поступает вообще или поступает искажённая. Таким образом, в условиях невесомости, информация, поступающая с вестибулярного аппарата не совпадает с информацией, поступающей от других сенсорных систем, при этом привычные сенсорные связи нарушаются и мозг в начале полета не может правильно интерпретировать поступающие в него сигналы, что приводит к развитию сенсорного конфликта, первым уровнем которого является отолито-каналовый конфликт, затем вестибуло-зрительный, далее - межсенсорная дезинтеграция.
Понимая эти особенности функционирования отолитов в невесомости и последующей межсенсорной дезинтеграции, многие исследователи предвидели развитие сенсорных нарушений в условиях невесомости в виде вестибулярных дисфункций. Полёт Германа Титова и последующие космические полёты подтвердили предположения исследователей.
В начальные периоды адаптации к условиям невесомости и реадаптации к земной гравитации практически всеми космонавтами отмечается ряд специфических, атипичных сенсорных реакций, в частности ориентационные иллюзии, головокружение, затруднение при фиксации и прослеживании зрительных объектов в поле зрения. Данное состояние ощущается как дискомфортное, особенно при наличии дополнительной вегетативной симптоматики.
Феноменологическая схожесть симптомов, развивающихся в невесомости и при реадаптации к земной гравитации, с земными формами болезни движения определило название - КОСМИЧЕСКАЯ ФОРМА БОЛЕЗНИ ДВИЖЕНИЯ (КБД).
КБД - это состояние, когда нормальная физиологическая адаптация переходит в фазу декомпенсации на уровне интегративных механизмов ЦНС, т.е. когда адаптационные реакции космического адаптационного синдрома (КАС) достигают клинического уровня и сопровождаются ухудшением профессиональной работоспособности.
Многие исследователи развитие КБД рассматривают с позиций интралабиринтных и экстралабиринтных механизмов. На наш взгляд такое деление неправомерно, так как вестибулярный аппарат в силу обширности функциональных связей вестибулярной системы с другими сенсорными системами, облигатно участвует во всех целостных реакциях организма.
Целью проведённых работ являлось:
- определение характера взаимодействия отолито-окуломоторной и купуло-окуломоторной подсистем при реадаптации к земным условиям после длительного пребывания в условиях невесомости;
- исследование влияния невесомости на характеристики всех форм зрительного слежения (плавное слежение, фиксационные саккады, удержание взора);
- определение роли и значения вестибулярной системы в обеспечении следящей функции глаз;
- уточнение взаимосвязи между показателями следящей функции глаз и состоянием вестибулярной функции;
- определение степени вовлеченности разных уровней иннервационных механизмов двигательного аппарата глаз в развитии нарушений зрительного слежения после космических полетов.
Оценка вестибулярной функции и следящих движений глаз была проведена с использованием компьютерных стимуляционных программ на АПК «СЕНСОМОТОР», «ОКУЛОСТИМ», «ВИРТУАЛ» (последний - на борту РС МКС в ходе космического полета), основу которых составляют:
- Персональный компьютер с двумя мониторами: один для предъявления зрительной стимуляции обследуемому космонавту, второй - для контроля врачом-экспериментатором за физиологическими реакциями обследуемого.
- Окулотахогониограф - устройство с 3-х осевыми акселлерометрическими датчиками и датчиками угловой скорости, регистрирующими скорость и угол поворота головой, а также усилитель электроокулограмм (ЭОГ), работающий на постоянном токе. Точность определения поворота глаз составляет 0.5-1°.
- Джойстик для регистрации начала, направления, интенсивности и окончания головокружения и других координационных иллюзий.
- Программно-математическое обеспечение - комплекс компьютерных тестов, обеспечивающих стимуляцию (раздельную, избирательную, сочетанную и полимодальную, строго дозируемую и строго направленную) зрительного и вестибулярного сенсорных входов, регистрацию физиологических реакций, хранение и обработку регистрируемых сигналов.
Обработка и анализ записей движений глаз и головы осуществлялась с помощью программного обеспечения, разработанного специалистами лаборатории «ВЕСТИБУЛЯРНАЯ ФИЗИОЛОГИЯ» ГНЦ РФ - ИМБП РАН (Сагалович С.В., Стефанков Д.В., Азаров К.А., Наумов И.А.).
На Рис. 1 представлено проведение послеполетного обследования космонавтов с использованием АПК «СЕНСОМОТОР» и «ОКУЛОСТИМ»
Рис. 1. Послеполетное обследование космонавтов с использованием АПК «СЕНСОМОТОР» и «ОКУЛОСТИМ»
Параллельно с методом ЭОГ был применен метод видеоокулографии (ВОГ). Движения глаз регистрировали с помощью ВОГ-комплексов VNG Ulmer (производства фирм Synapsys и Heinemann Medizintechnik GmbH) и Chronos Vision ETD (производства фирмы Chronos Vision GmbH) (Рис. 2).
Для регистрации горизонтальных, вертикальных и торсионных движений глаз на голову испытуемого надевались очки (шлем), снабженные инфракрасными видеокамерами с частотой развертки 50 Гц (в случае комплекса VNG Ulmer) и 100-400 Гц (в случае комплекса Chronos Vision ETD). Диапазон регистрируемых движений глаз составлял до 55° по горизонтали и до 35° по вертикали, точность распознавания положения глаза - 0.25° (VNG Ulmer) и 17/12/2018 21-суточная иммерсия 2018
В 2018 г. сотрудники лаборатории приняли участие в 21-суточном эксперименте «сухая» горизонтальная иммерсия с участием 6-ти испытателей в этом году и продолжением со следующими — в 2019. В ходе эксперименте кроме исследования зрительно-мануального слежения и состояния вестибулярной функции, сотрудник лаборатории Дмитрий Глухих принял непосредственное участие в качестве испытателя.
С 10 по 12 декабря в г. Москва прошла XVII Конференцию по космической биологии и авиакосмической медицине c международным участием, посвященная 100-летию со дня рождения академика О.Г. Газенко. Сотрудники лаборатории Наумов И.А. и Глухих Д.О. успешно выступили с докладами, посвященными исследованию вестибуло-зрительного взаимодействия в условиях измененной гравитации и зрительно-мануального слежения и вестибулярной функции в условиях […]
Физиологические основы поддержания равновесия
Причиной головокружения в большинстве случаев служит нарушение согласованной деятельности различных сенсорных систем - вестибулярной, зрительной, проприоцептивной (информация о положении тела в пространстве, получаемая от рецепторов, расположенных главным образом в мышцах и сухожилиях). Кроме того, важной, а иногда и доминирующей причиной возникновения головокружения является дисфункция центральных структур, участвующих в поддержании равновесия тела, главным образом, ядер мозжечка.
Вестибулярная система
Вестибулярная система состоит из:
- лабиринта,
- вестибулярной части преддверно-улиткового нерва,
- вестибулярных ядер в стволе головного мозга, а также их связей с другими отделами ЦНС (центральной нервной системы).
Правильная работа вестибулярной системы позволяет человеку четко ориентироваться в трехмерном пространстве, а именно:
- воспринимать положение тела относительно вектора силы тяжести (статический компонент);
- ощущать направление и скорость движения тела при его угловых и линейных перемещениях (динамический компонент).
Лабиринт располагается в каменистой части височной кости и включает:
- отолитовый аппарат, который представлен двумя сообщающимися камерами (саккулус и утрикулус);
- системой трех полукружных каналов, располагающихся во взаимоперпендикулярных плоскостях.
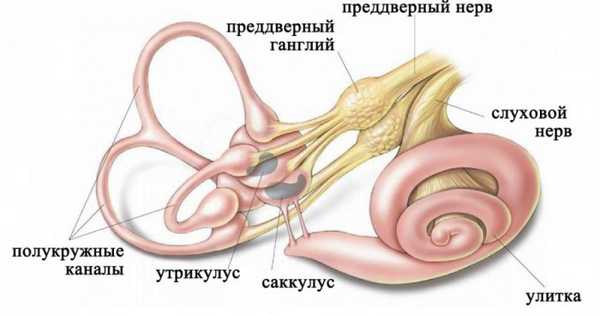
Строение лабиринта
В каждой камере отолитового аппарата и в каждом полукружном канале имеется скопление рецепторных клеток - макула, которая покрыта желатинообразной массой - купулой. В отолитовом аппарате купула покрывает волосковые клетки наподобие подушки и содержит отложения кристаллов кальцита (отолиты), которые придают купуле дополнительный вес.
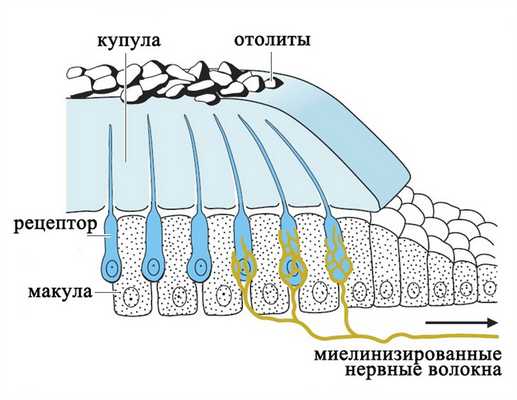
Отолитовый аппарат
В полукружных каналах желатинообразная масса не содержит отолитов и полностью перекрывает просвет канала.
Рецепторы вестибулярной системы представлены волосковыми клетками, которые несут на апикальной поверхности от 60 до 80 тонких выростов цитоплазмы (стереоцилий) и одну ресничку (киноцилию).
Восприятие положения тела относительно силы гравитации
При вертикальном положении головы макула утрикулуса располагается горизонтально. Когда голова наклоняется в сторону, утяжеленная отолитами желатинообразная мембрана под действием силы тяжести соскальзывает в сторону наклона. Это скольжение приводит к изгибанию стереоцилей волосковых клеток. Наклон стереоцилей сопровождается (в зависимости от направления) повышением или снижением частоты нервных импульсов в чувствительных нейронах вестибулярного ганглия. Макула саккулуса располагается вертикально и действует таким же образом.
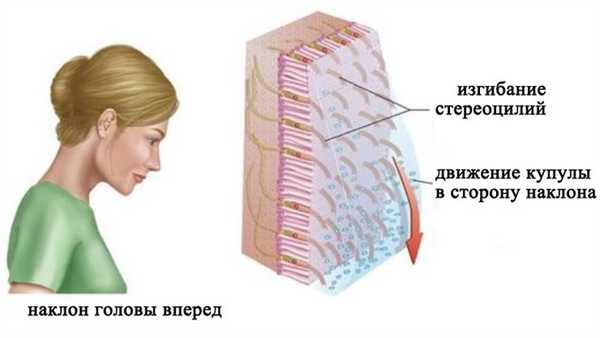
Восприятие положения тела относительно силы гравитации
Восприятие линейных ускорений
При резком линейном ускорении тела купула саккулуса или утрикулуса за счет сил инерции смещается в направлении, противоположном направлению движения, что также приводит к изменению электрической активности рецепторов.
Восприятие углового ускорения
Три полукружных канала расположены в трех разных плоскостях. Каждый из трех каналов действует как замкнутая трубка, заполненная лимфой. В расширенной части канала его внутренняя стенка выстлана волосковыми клетками, а расположенная над ними купула полностью перекрывает просвет канала. При повороте головы полукружные каналы поворачиваются вместе с ней, а эндолимфа в силу своей инерции в первый момент остается на месте. В результате этого возникает разность давлений по обе сторону купулы, и она прогибается в направлении, противоположном движению. Это вызывает деформацию стереоцилий и последующее изменение активности нейронов.
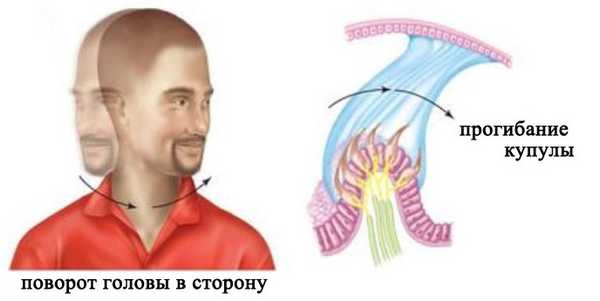
Восприятие углового ускорения
При вращении головы только в горизонтальной, сагитальной или фронтальной плоскости активируются рецепторы одного из соответствующих каналов. При сложном вращении головы активируются рецепторы всех трех каналов. Информация от них поступает в головной мозг и на основе ее конвергенции и анализа модулируется истинная картина перемещения головы.
Центральный отдел вестибулярной системы
Аксоны чувствительных нейронов, тела которых располагаются в вестибулярном ганглии, следуют в продолговатый мозг и оканчиваются в четырех парных вестибулярных ядрах. Приходящие в эти ядра импульсы от рецепторов дают точную информацию о положении в пространстве исключительно головы (но не всего тела!), поскольку она может быть наклонена или повернута относительно туловища. Для восприятия положения тела в пространстве необходим также учет угла наклона и поворота головы относительно туловища, поэтому вестибулярные ядра получают дополнительные стимулы от проприорецепторов мышц шеи.
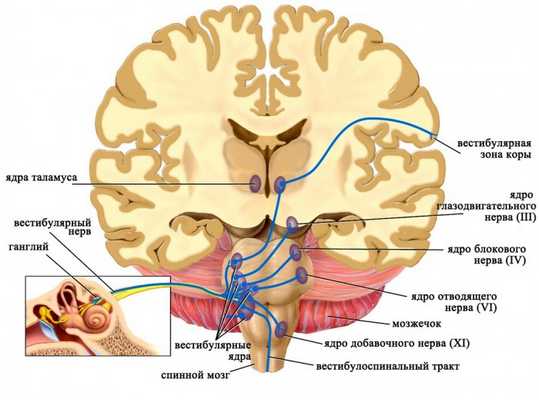
Ядра вестибулярного нерва и их связи
Далее от вестибулярных ядер афферентная импульсация направляется к нейронам специфических ядер таламуса, а отростки последних достигают постцентральной извилины коры больших полушарий головного мозга
Проприоцептивная система
Благодаря проприоцепции, мы ощущаем положение конечностей, движение и степень мышечного напряжения в них. Это дает человеку чувство “опоры”, т.е. осознание, что стопы опираются на какую-либо поверхность, удерживая вес тела. Рецепторный аппарат проприоцептивной чувствительности, расположен в мышцах, сухожилиях, фасциях, капсулах суставов, а также в коже.
Необходимо отметить, что важную роль в поддержании равновесия тела играют рецепторы глубокой чувствительности, расположенные не только в конечностях, но и в структурах шеи, главным образом, в глубоких мышцах. Информация, получаемая головным мозгом от этих рецепторов, необходима для пространственной ориентации человека, поддержании его позы, а также координинации движения головы и туловища.
Зрительная система
Эффективное поддержание равновесия требует четкого контроля со стороны зрительной системы (в соответствие с принципом обратной связи). При этом контроль над движениями мышц глазного яблока является чрезвычайно сложным процессом. Существует 3 основных системы контроля взора:
- Система саккадических движений глазных яблок;
- Система плавных (следящих) движений глазных яблок;
- Вестибуло-окулярная система.
В пределах головного мозга эти системы контролируются определенными анатомическими зонами, которые являются в значительной степени изолированными, и обеспечивают две главные функции:
- зафиксировать предмет рассматривания в периферии визуальной области, поворачивая к нему глаза;
- удержать изображение предмета рассматривания устойчивым на ямке сетчатки.
Система саккадических движений глазных яблок
Когда объект интереса появляется в периферии визуальной области, происходит быстрый поворот глазных яблок в его сторону, так, что изображение объекта проецируется на сетчатку в области желтого пятна. Тот же самый двигательный ответ глазных яблок может быть вызван внезапным звуком или болезненным стимулом. Такое быстрое движение глаз называется саккадическим, от французского слова, означающего резкое движение парусника при ветре или дергание головы лошади от потягивания узды. В целом, система саккадических движений глазных яблок обеспечивает обнаружение зрительной цели и выведение ее на наиболее чувствительную часть сетчатой оболочки. Саккады возникают, например, в процессе чтения, при этом глаза человека обычно совершают несколько саккадических движений на каждой строке. Кроме того, они появляются, когда человек рассматривает какой-либо объект (картину, скульптуру и пр.), но в этом случае саккады совершаются в разных направлениях (вверх, вниз, в стороны и под углом) последовательно от одной точки объекта к другой.
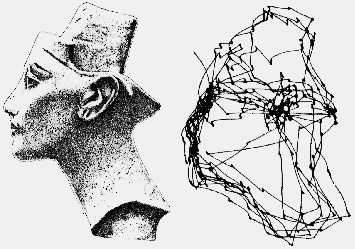
Классическое изображение, описывающее саккадические движение глазных яблок
при рассматривании объекта
Система плавных (следящих) движений глазных яблок
Когда объект рассматривания перемещается, саккадическая система может первоначально зафиксировать его, но скоро теряет, поскольку изображение ускользает из области желтого пятна (сетчатое скольжение). Плавные (следящие) движения глаз необходимы для длительной фиксации движущегося объекта и слежения за ним. После того как визуальная цель выбрана, система работает вне волевого контроля.
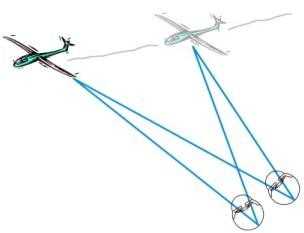
Схематическое изображение функционирования системы
плавных (следящих) движений глаз
Вестибуло-окулярная система
В то время как система следящих движений глазных яблок фиксирует изображение перемещающегося объекта рассматривания на желтом пятне, существует другая система, которая позволяет стабилизировать изображение неподвижного объекта рассматривания на сетчатке во время движения головы. Это основная функция вестибуло-окулярной системы. Благодаря ее наличию у человека во время движения на транспорте по неровной дороге или ходьбе не возникает проблем с четким рассматриванием отдаленного объекта. В том случае, когда по какой-либо причине вестибуло-окулярная система не работает возникает феномен, называющийся “осциллопсия” - “дергание” визуальной картинки при движении.
Мозжечок
Основная функция мозжечка заключается в получении информации о положении тела в пространстве от всех органов чувств и регуляции на ее основе мышечного тонуса и движений для поддержания равновесия и выполнения точных действий.
Для больных с повреждением мозжечка характерна астазия-абазия - нарушение способности к сохранению равновесия тела при стоянии и ходьбе. Больные ходят, широко расставив ноги - так называемая туловищная атаксия (“пьяная походка”).
Ходьба на пятках и носках невозможна. Атаксия в данном случае развивается вследствие неспособности головного мозга координировать деятельность мышц в процессе преодоления силы тяжести. Также выявляются глазодвигательные расстройства. Они проявляются нарушением фиксации взора на неподвижных или двигающихся объектах, в результате чего возникают рывковые движения глаз при слежении. Также характерен вертикальный нистагм, бьющий вверх или вниз.
Физиология равновесия.1.1.Функции вестибулярного анализатора .Организация вестибулярного аппарата.
Вестибулярная сенсорная система состоит из следующих отделов:
периферический отдел включает два образования, содержащие механорецепторы вестибулярной системы — преддверие (мешочек и маточка) и полукружные каналы;
корковый отдел представляют четвертые нейроны, часть которых представлена в проекционном (первичном) поле вестибулярной системы в височной области коры, а другая часть — находится в непосредственной близости к пирамидным нейронам моторной области коры и в постцентральной извилине. Точная локализация коркового отдела вестибулярной сенсорной системы у человека в настоящее время не установлена.
Строение и функции вестибулярного анализатора
Вестибулярный анализатор имеет важное значение в регуляции положения тела в пространстве и его движений. Периферический отдел вестибулярного анализатора является частью внутреннего уха и состоит из полукружных каналов, размещенных в трех взаимно перпендикулярных плоскостях, и из статоцистных органов - двух мешочков - овального (маточки) и круглого, который расположен ближе к улитке.
Вестибулярный орган (орган равновесия, орган гравитации) состоит из трех полукружных каналов и преддверия.Полукружные каналы расположены в трех взаимно перпендикулярных плоскостях: верхний—во фронтальной,задний—в сагиттальной инаружный—в горизонтальной. Преддверие состоит из двух мешочков—круглого (саккулюс), расположенного ближе к улитке, иовального (утрикулюс)расположенного ближе к полукружным каналам.
Полукружные каналы своими устьями открываются в преддверие и сообщаются с ним пятью отверстиями (колена двух каналов: верхнего и заднего — соединены вместе). Один конец каждого канала имеет расширение, которое называется ампулой. Все эти структуры состоят из тонких перепонок и образуют перепончатый лабиринт, внутри которого находится эндолимфа.Вокруг перепончатого лабиринта и между ним и костным его футляром имеетсяперилимфа,которая переходит в перилимфу органа слуха. В каждом мешочке преддверия имеются небольшие возвышения, называемые пятнами, а в ампулах полукружных каналов — гребешками. Они состоят из нейроэпителиальных клеток, имеющих на свободной поверхности волоски (реснички), которые разделяются на две группы: тонкие (их много) — стереоцилии и один более толстый и длинный на периферии пучка—киноцилия
Волосковые клеткипредставляют собой рецепторы вестибулярного анализатора иявляются вторичными. Рецепторные клетки преддверия покрыты желеобразной массой, состоящей в основном из мукополисахаридов; благодаря содержанию в ней значительного количества кристаллов карбоната кальция она получила названиеотолитовой мембраны. В ампулах полукружных каналов желеобразная масса не содержит солей кальция и называетсялистовидной мембраной (купула). Волоски рецепторных клеток пронизывают эти мембраны.
Возбуждение волосковых клеток происходит вследствие скольжения мембраны по волоскам, изгибания волосков (стереоцилии) в сторону киноцилий. Это связано с тем, что механическое управление ионными каналами мембраны волоска с помощью микрофиламентов, зависит от направления сгиба волоска:отклонение в одну сторону приводит к открыванию каналов и деполяризации волосковой клетки, а отклонение в противоположном направлении вызывает закрытие каналов и гиперполяризацию рецептора.
В волосковых клетках преддверия и ампулы при их сгибании генерируется рецепторный потенциал, который усиливает выделение ацетилхолина и через синапсы активирует окончания волокон вестибулярного нерва.
проводниковый отдел. К рецепторам подходят периферическиеволокна биполярных нейронов вестибулярного ганглия, расположенного во вутреннем слуховом проходе (первый нейрон). Аксоны этих нейронов в составе вестибулярного нерва направляются квестибулярым ядрам продолговатогомозга (второй нейрон). Вестибулярные ядра продолговатого мозга (верхнее—ядро Бехтерева,медиальное—ядро Швальбе,латеральное—ядро Дейтерсаинижнее—ядро Роллера) получают дополнительную информацию по аф- ферентным нейронам от проприорецепторов; мышц или от суставных сочленений шейного отдела позвоночника. Эти ядра, где расположен второй нейрон вестибулярного анализатора, тесносвязаны с различными отделами центральной нервной системы.Благодаря этому обеспечиваются контроль и управление эффекторными реакциями соматического, вегетативного и сенсорного характера.
третий нейронрасположен вядрах зрительного бугра, откуда возбуждение направляется в кору большого полушария.
Центральный отделвестибулярного анализатора локализуется ввисочной области коры большого мозга, несколько кпереди от слуховой проекционной зоны (21—22-е поля по Бродману, четвертый нейрон).
Нервные волокна, выходящие из вестибулярных ядер, образуют связи с другими отделами центральной нервной важнейшими из них являются следующие: вестибулоспинальные, вестибуловегетативныеивестибулоглазодвигательные. Эти связи обеспечивают контроль и управление различными двигательными реакциями, а также являются основой для рефлексов обеспечивающих равновесие.
а. Вестибулоспинальный тракт, волокна которого в конечном итоге оказывают влияние в основном наγ-мотонейроны мышц-разгибателей, хотя часть волокон оканчивается и наα-мотонейронах.
б. Связи с мотонейронамишейного отдела спинного мозга, входящие главным образом в вестибулоспинальный тракт.
в.Связи с ядрамиглазодвигательного нерва, которые опосредуют движения глаз, вызываемые вестибулярной активностью; эти волокна проходят в составе медиального продольного пучка. Функцию статолитового аппарата можно тестировать, наблюдая глазодвигательные реакции при наклонах головы.
г. Тракты, направляющиеся ввестибулярные ядра противоположной стороны мозга, благодаря которым афферентация с обеих сторон тела может обрабатываться совместно.
д. Связи сретикулярной формацией, посредством которых обеспечивается воздействие на ретикулоспинальный тракт, являющийся еще одним (полисинаптическим) путем кα- иβ-мотонейронам.
е. Тракты, проходящие через таламус впостцентральную извилину коры головного мозга, обеспечивающие сознательную обработку вестибулярной информации и таким образом сознательную ориентацию в пространстве.
ж.Волокна, направляющиеся вгипоталамус, которые в основном участвуют в возникновении кинетозов (укачивания).
з. Связи смозжечком, в особенности сarchicerebellum( вторичные вестибулярные афференты).мозжечок, может получать некоторые первичные вестибулярные афференты (так называемый прямой сенсорный мозжечковый путь). И первичные, и вторичные вестибулярные афференты у млекопитающих оканчиваются в мозжечке мшистыми волокнами на клетках-зернах флокку-лонодулярной доли (относящейся кarchicerebellum) и частично клеткахuvulaиparaflocculus(paleocerebellum). Клетки-зерна оказывают возбуждающее действие на клетки Пуркине в этих областях, а аксоны последних проецируются опять-таки в вестибулярные ядра. Такая цепь осуществляет тонкую регулировку вестибулярных рефлексов. При нарушениях функции мозжечка вследствие различных заболеваний эти рефлексы утрачивают тормозной компонент, что проявляется в возникновении таких симптомов, как усиленный или спонтанно возникающий нистагм, утрата равновесия, проявляющаяся в неустойчивой походке, избыточной амплитуде движений, особенно при ходьбе («петушиный шаг»). Эти симптомы являются частью синдромамозжечковой атаксии.
Читайте также: