Эндоплазматическая сеть и рибосомы. Виды эндоплазматической сети.
Добавил пользователь Евгений Кузнецов Обновлено: 28.01.2026
Эндоплазматическая сеть (ЭПС) - органелла, обеспечивающая синтез углеводов, липидов и белков, а также начальные посттрансляционные изменения последних. Она имеет одномембранное строение и состоит из системы уплощенных, удлиненных, трубчатых и везикулярных образований. Название органеллы обусловлено характером связи этих элементов друг с другом, образующих в цитоплазме непрерывную трехмерную сеть, элементы которой лишь на отдельных срезах могут иметь вид изолированных структур. Мембрана ЭПС тоньше, чем плазмолемма и содержит более высокую концентрацию белка, что связано с наличием в ней многочисленных ферментных систем. Степень развития ЭПС и особенности ее строения варьируют в различных клетках и зависят от их функции. Выделяют 3 разновидности ЭПС: гранулярную ЭПС (грЭПС) и гладкую, или агранулярную ЭПС (аЭПС), которые связаны друг с другом в области перехода, называемой переходной (транзиторной) ЭПС.
Гранулярная ЭПС обеспечивает (1) биосинтез всех мембранных белков и белков, предназначенных для экспорта из клетки, и (2) начальное гликозилирование и посттрансляционные изменения белковых молекул. Гранулярная ЭПС образована уплощенными мембранными цистернами и трубочками, на наружной поверхности которых располагаются рибосомы и полисомы, придающие мембранам зернистый (гранулярный) вид, что и отражено в названии органеллы. Мембраны грЭПС содержат особые белки, которые обеспечивают (1) связывание рибосом и (2) уплощение цистерн. Полость грЭПС содержит рыхлый материал умеренной плотности (продукты синтеза) и сообщается с перинуклеарным пространством . Благодаря грЭПС происходит отделение (сегрегация) вновь синтезированных белковых молекул от гиалоплазмы.
Агранулярная (гладкая) ЭПС представляет собой трехмерную замкнутую сеть мембранных анастомозирующих трубочек, канальцев, цистерн и пузырьков диаметром 20-100 нм, на поверхности которых рибосомы отсутствуют , что определило ее название. Соответственно, на мембранах аЭПС отсутствуют рецепторы, связывающие субъединицы рибосом (рибофорины). Предполагают, что аЭПС образуется в результате формирования выростов грЭПС, мембрана которых утрачивает рибосомы.
Функции аЭПС включают: (1) синтез липидов, в том числе мембранных (2) синтез гликогена, (3) синтез холестерина, (4) детоксикацию эндогенных и экзогенных веществ, (5) накопление ионов Са 2+ , (6) восстановление кариолеммы в телофазе митоза (эта функция оспаривается авторами, считающими, что кариолемма восстанавливается за счет мембранных пузырьков, на которые она ранее распалась). Помимо указанных основных функций, в некоторых типах клеток аЭПС выполняет ряд дополнительных - например, в мегакариоцитах (гигантских клетках костного мозга) ее элементы образуют демаркационные каналы, разделяющие формирующиеся тромбоциты.
Переходная (транзиторная) ЭПС - участок перехода грЭПС в аЭПС у формирующейся поверхности комплекса Гольджи. В области переходной ЭПС трубочки распадаются на отдельные фрагменты, образующие окаймленные транспортные пузырьки, которые переносят материал из ЭПС в комплекс Гольджи.
Эндоплазматическая сеть
Эндоплазматическая сеть (ЭПС) была открыта К.Р. Портером в 1945 г. Эта мембранная органелла общего назначения представляет собой совокупность вакуолей, плоских мембранных мешков или трубчатых образований, создающих как бы мембранную сеть внутри цитоплазмы. Различают два типа — гранулярную (шероховатую) и гладкую эндоплазматическую сеть.
Гранулярная эндоплазматическая сеть (reticulum endoplasmaticum granulosum) представлена замкнутыми мембранами, которые образуют уплощенные мешки, цистерны, трубочки. Ширина полостей цистерн значительно варьирует в зависимости от функциональной активности клетки. Наименьшая ширина их — около 20 нм, но они могут достигать диаметра в несколько микрометров. Отличительной чертой мембран гранулярной ЭПС является то, что они со стороны гиалоплазмы покрыты рибосомами (рис. 8).
Гранулярная эндоплазматическая сеть бывает представлена редкими разрозненными цистернами или их локальными скоплениями. Первый тип гранулярной эндоплазматической сети, характерен для малоспециализированных ктеток или для клеток с низкой метаболической активностью. Скопления эндоплазматической сети являются принадлежностью клеток, активно синтезирующих секреторные белки. Так, в клетках печени и некоторых нервных клетках гранулярная эндоплазматическая сеть собрана в отдельные зоны. В клетках поджелудочной железы гранулярная эндоплазматическая сеть в виде плотно упакованных друг около друга мембранных цистерн занимает базальную и околоядерную зоны клетки.
Рибосомы, связанные с мембранами эндоплазматической сети, участвуют в синтезе белков, выводимых из данной клетки («экспортируемые» белки). Кроме того, гранулярная эндоплазматическая сеть принимает участие в синтезе белков — ферментов, необходимых для организации внутриклеточного метаболизма, а также используемых для внутриклеточного пищеварения.
Белки, накапливающиеся в полостях эндоплазматической сети, могут, минуя гиалоплазму, транспортироваться в вакуоли комплекса Гольджи, где они часто модифицируются и входят в состав либо лизосом, либо секреторных гранул, содержимое которых остается изолированным от гиалоплазмы мембраной. В ряде случаев внутри самих канальцев или вакуолей гранулярной эндоплазматической сети может происходить модификация белков, например связывание их с сахарами (глюкозилирование), или конденсация синтезированных белков с образованием крупных агрегатов — секреторных гранул.
В гранулярной эндоплазматической сети происходит синтез мембранных интегральных белков (см. часть 2), которые встраиваются в толщу мембраны.
Итак, роль гранулярной эндоплазматической сети заключается в синтезе на ее рибосомах экспортируемых белков, в их изоляции от содержимого гиалоплазмы внутри мембранных полостей, в транспорте этих белков в другие участки клетки, в химической модификации таких белков и в их локальной конденсации, а также в синтезе структурных компонентов клеточных мембран.
Агранулярная (гладкая) эндоплазматическая сеть (reticulum endoplasmaticum nongranulosum) также представлена мембранами, образующими мелкие вакуоли и трубки, канальцы, которые могут ветвиться, сливаться друг с другом. В отличие от гранулярной эндоплазматической сети на мембранах гладкой эндоплазматической сети нет рибосом. Диаметр вакуолей и канальцев гладкой эндоплазматической сети обычно около 50—100 нм.
Гладкая эндоплазматическая сеть возникает и развивается за счет гранулярной эндоплазматической сети (при освобождении ее от рибосом).
Деятельность гладкой эндоплазматической сети связана с метаболизмом липидов и некоторых внутриклеточных полисахаридов. Гладкая эндоплазматическая сеть участвует в заключительных этапах синтеза липидов. Она сильно развита в клетках, секретирующих такие категории липидов, как стероиды, например, в клетках коркового вещества надпочечников, в сустентоцитах семенников.
Тесная топографическая связь гладкой эндоплазматической сети с отложениями гликогена (запасной внутриклеточный полисахарид животных) в гиалоплазме различных клеток (клетки печени, мышечные волокна) указывает на ее возможное участие в метаболизме углеводов.
В поперечнополосатых мышечных волокнах гладкая эндоплазматическая сеть способна депонировать ионы кальция, необходимые для функции мышечной ткани.
Очень важна роль гладкой эндоплазматической сети в дезактивации различных вредных для организма веществ за счет их окисления с помощью ряда специальных ферментов. Особенно четко она проявляется в клетках печени. Так, при ряде отравлений в клетках печени появляются ацидофильные зоны (не содержащие РНК), сплошь занятые гладким эндоплазматическим ретикулумом.
7. Эндоплазматическая сеть, строение, виды эпс. Строен и функц рибосом.
Впервые эндоплазматический ретикулум был обнаружен американским учёным К. Портером в 1945 году посредством электронной микроскопии.
Эндоплазматический ретикулум (ЭПР) (лат. reticulum — сеточка) или эндоплазматическая сеть (ЭПС) — внутриклеточный органоид эукариотической клетки, представляющий собой разветвленную систему соединённых между собой каналов и полостей, ограниченных одинарной мембраной, поверхность которой составляет более 50% площади всех клеточных мембран. Мембрана ЭПС тоньше чем плазмалемма и содержит более высокую концентрации. белка. Непосредственным продолжение ЭПС является наружная ядерная мембрана.
На поверхности мембран ЭПС происходит большая часть реакций метаболизма, протекающих в клетке. ЭПС разделяет цитоплазму на отдельные отсеки. по каналам ЭПС происходит упорядоченный обмен веществами и энергией между различными компонентами клетки.
ЭПС - генератор мембран для плазмолеммы, ап гольджи и лизосом.
Гранулярная или шероховатая эпс.
наружная обращеная к цитоплазме, сторона грЭПС покрыта рибосомами (котор имеют вид мелк гранул; поступают из ядра благодаря связи мембраны с наруж мембр ядра).
грЭПС - образ уплощенными мембранными цистернами и трубочками на наружной поверхности которых располог рибосомы и полисомы, придающие мембране зернист вид.
Мембраны содерж белки (которые обеспеч связывание рибосом, уплощение цистерн).
Полость грЭПС сообщ с перенуклеарн пространство. Благодаря грЭПС происход отделение вновь синтезированных белковых молекул от гиалоплазмы.
грЭПС хорошо развита в клетках, специализирующихся на белковом синтезе.
ФУНКЦИИ: 1)биосинтез всех мембранных белков, предназначенных для экспорта из клетки.
2) в грЭПС происход посттрансляционный процессинг белков. (созревание белка). белки приобрет характер для них третичную или четвертичную структуру. потом транспортир в комплекс гольджи - > потом в другие органойды.
3) гЭПС выполняет ф-ю пространственного разделения ферментных систем. резделени клетки с помощью мембран на отдел отсеки - компарменты.
4) обеспеч транспорт синтезируемых веществ в аппарат гольджи.
Гладкая или агранулярная ЭПС.
не имеет рибосом. Сост из сильно ветвящихся канальцев и мелких вакуолей диаметром 20-100 нм. гЭПС - трёхмерная замкнутая сеть мембранных анастамозирующих турбочек, канальцев, цистерн и пузырьков диаметром 20-100 нм, на поверхности которых рибосомы отсутсвт.
На цитоплазмотической поверхности гЭПС синтезируется большая часть липидов клетки, которые вход в состав всех её мембран. Часть синтезир на гЭПС белков и липидов встраивается в неё, но увеличения общей площади мембраны при этом не происход. на гЭПС соверш синтез и распад многих углеводов, включ полисахариды, образ стеройдные гормоны.
В Гэпс накаплив многие ядовит в-ва, подлежащ удален из клетки.
гЭПС наиболее развита в клетках с интенсивным жировыми углеводным обменом.
ФУНКЦИИ: 1) синтез липидов; (на мембранах) 2) синтез гликогена (в клетках печени)
3) синтез холестерина и других стеройдов 4) детоксикация эндогенных и экзогенных в-в. (в клетках печени) 5) накопление ионов Са. гЭПС в Миш клетках играет роль депо ионов кальция, необходимых для мыш сокращ. 6) компартментализация (эпс раздел клетку на отдел отсеки) 7) транспорт синтезируемых веществ 8) в мегакариоцитах элементы гЭПС образуют демаркационные каналы, разделяющие формирующие тромбоциты. 9) восстановление кариолеммы в телофазе митоза.
Рибосомы впервые были описаны как уплотненные частицы, или гранулы, клеточным биологом румынского происхождения Джорджем Паладе в середине 1950-х годов [1] . В 1974 г. Паладе, Клод и Кристиан Де Дюв получили Нобелевскую премию по физиологии и медицине «за открытия, касающиеся структурной и функциональной организации клетки». Термин "рибосома" был предложен Ричардом Робертсом в 1958 вместо "рибонуклеобелковая частица микросомальной фракции" [
Рибосома — важнейший органоид живой клетки сферической или слегка овальной формы, диаметром 100-200. В эукариотических клетках рибосомы располагаются на мембранах эндоплазматического ретикулума, хотя могут быть локализованы и в неприкрепленной форме в цитоплазме. Синтез рибосом у эукариот происходит в специальной внутриядерной структуре - ядрышке. Рибосомы представляют собой нуклеопротеид. Рибосомная РНК составляет около 70 % всей РНК клетки.
Рибосома- место синтеза белка. Каждая рибосома сост из 2х частей (субъединиц) - большой и малой. Построены они из равных частей (по массе) белка ирнк. РНК входящ в сост рибосом наз рибосомальной. рРНК синтез в ядрышке.
Основным методом выделения рибосом является осаждение центрифугированием. Этот метод позволяет выделить два основных типа рибосом, которые называются 70S-рибосомами и 8OS-рибосомами. (S — сведбсрг — единица, характеризующая скорость осаждения в центрифуге; чем больше число S. тем выше скорость осаждения). 70S - рибосомы обнаруживаются у прокариот и в хлоропластах и митохондриях эукариот. 8OS-рибосомы, несколько более крупные, находятся в цитоплазме эукариот. В процессе синтеза белка рибосомы движутся вдоль мРНК. Процесс идет более эффективно, если вдоль мРНК движется не одна, а несколько рибосом. Такие цепи рибосом на мРНК называют полирибосомами, или полисомами.
Вся внутренняя зона цитоплазмы заполнена многочисленными мелкими каналами и полостями, стенки которых представляют собой мембраны, сходные по своей структуре с плазматической мембраной. Эти каналы ветвятся, соединяются друг с другом и образуют сеть, получившую название эндоплазматической сети.
Эндоплазматическая сеть неоднородна по своему строению. Известны два ее типа - гранулярная и гладкая. На мембранах каналов и полостей гранулярной сети располагается множество мелких округлых телец - рибосом, которые придают мембранам шероховатый вид. Мембраны гладкой эндоплазматической сети не несут рибосом на своей поверхности.
Эндоплазматическая сеть выполняет много разнообразных функций. Основная функция гранулярной эндоплазматической сети - участие в синтезе белка, который осуществляется в рибосомах.
На мембранах гладкой эндоплазматической сети происходит синтез липидов и углеводов. Все эти продукты синтеза накапливаются н каналах и полостях, а затем транспортируются к различным органоидам клетки, где потребляются или накапливаются в цитоплазме в качестве клеточных включений. Эндоплазматическая сеть связывает между собой основные органоиды клетки.
ЯДРО (клеточное ядро), в биологии — обязательная часть клетки у многих одноклеточных и всех многоклеточных организмов. Размеры от 1 мкм (у некоторых простейших) до 1 мм (в яйцах некоторых рыб и земноводных). Все организмы нашей биосферы как одноклеточные, так и многоклеточные, подразделяются на эукариот— их клетки содержат ядро, и прокариот, клетки которых не имеют морфологически оформленного ядра. Термин «ядро» (лат. nucleus) впервые применил Р. Броун в 1833 году, когда описывал шарообразные структуры, наблюдаемые им в клетках растений.
Внутреннее пространство клеточного ядра отделено от цитоплазмы ядерной оболочкой, состоящей из двух мембран. Мембраны оболочки ядра сходны по строению с другими мембранными компонентами клетки и построены по тому же принципу: это тонкие липопротеидные пленки, состоящие из двойного слоя липидных молекул, в который встроены молекулы белков. Пространство между внутренней и внешней ядерными мембранами называется перинуклеарным. На поверхности внешней ядерной мембраны обычно располагается большое количество рибосом, и иногда удается наблюдать непосредственный переход этой мембраны в систему каналов гранулярной эндоплазматической сети клетки. Внутренняя ядерная мембрана связана с тонким волокнистым белковым слоем — ядерной ламиной, состоящей из белков-ламинов. Густая сеть фибрилл ядерной ламины способна обеспечить целостность ядра, даже после растворения липидных мембран оболочки ядра в эксперименте. С внутренней стороны к ламине крепятся петли хроматина, заполняющего ядро.
Ядерная оболочка имеет отверстия диаметром около 90 нм, образующиеся засчет слияния внешней и внутренней ядерных мембран. Такие отверстия в оболочке ядра окружены сложными белковыми структурами, получившими название комплекса ядерной поры. Восемь белковых субъединиц, входящих в состав ядерной поры, располагаются вокруг перфорации ядерной оболочки в виде колец, диаметром около120 нм, наблюдаемых в электронный микроскоп с обеих сторон ядерной оболочки. Белковые субъединицы комплекса поры имеют выросты, направленные к центру поры, где иногда видна «центральная гранула» диаметром 10-40 нм. Размер ядерных пор и их структура стандартны для всех клеток эукариот. Число ядерных пор зависит от метаболической активности клеток: чем выше уровень синтетических процессов в клетке, тем больше пор на единицу площади поверхности клеточного ядра. В процессе ядерно-цитоплазматического транспорта ядерные поры функционируют как некое молекулярное сито, пропуская ионы и мелкие молекулы (сахара, нуклеотиды, АТФ и др.) пассивно, по градиенту концентрации, и осуществляя активный избирательный транспорт крупных молекул белков и рибонуклеопротеидов, то есть комплексов рибонуклеиновых кислот (РНК) с белками. Так, например, белки, транспортируемые в ядро из цитоплазмы, где они синтезируются, должны иметь определенные последовательности примерно из 50 аминокислот, (т. наз. NLS последовательности), «узнаваемые» комплексом ядерной поры. В этом случае комплекс ядерной поры, затрачивая энергию в виде АТФ, активно транслоцирует белок из цитоплазмы в ядро.
Клеточное ядро является вместилищем практически всей генетической информации клетки, поэтому основное содержимое клеточного ядра — это хроматин: комплекс дезоксирибонуклеиновой кислоты (ДНК) и различных белков. В ядре и, особенно, в митотических хромосомах, ДНК хроматина многократно свернута, упакована особым образом для достижения высокой степени компактизации. Ведь все длинные нити ДНК, общая длина которых составляет, например, у человека около 164 см, необходимо уложить в клеточное ядро, диаметр которого всего несколько микрометров. Эта задача решается последовательной упаковкой ДНК в хроматине с помощью специальных белков. Основная масса белков хроматина — это белки гистоны, входящие в состав глобулярных субъединиц хроматина, называемых нуклеосомами. Всего существует 5 видов белков гистонов. Нуклеосома представляет собой цилиндрическую частицу, состоящую из 8 молекул гистонов, диаметром около 10 нм, на которую «намотано» чуть менее двух витков нити молекулы ДНК. В электронном микроскопе такой искусственно деконденсированный хроматин выглядит как «бусины на нитке». В живом ядре клетки нуклеосомы плотно объединены между собой с помощью еще одного линкерного гистонового белка, образуя так называемую элементарную хроматиновую фибриллу, диаметром 30 нм. Другие белки, негистоновой природы, входящие в состав хроматина обеспечивают дальнейшую компактизацию, т. е. укладку, фибрилл хроматина, которая достигает своих максимальнах значений при делении клетки в митотических или мейотических хромосомах. В ядре клетки хроматин присутствует как в виде плотного конденсированного хроматина, в котором 30 нм элементарные фибриллы упакованы плотно, так и в виде гомогенного диффузного хроматина. Количественное соотношение этих двух видов хроматина зависит от характера метаболической активности клетки, степени ее дифференцированности. Так, например, ядра эритроцитов птиц, в которых не происходит активных процессов репликации и транскрипции, содержат практически только плотный конденсированный хроматин. Некоторая часть хроматина сохраняет свое компактное, конденсированное состояние в течение всего клеточного цикла — такой хроматин называется гетерохроматином и отличается от эухроматина рядом свойств.
Репликация и транскрипция
Клетки эукариот содержат обычно несколько хромосом (от двух до нескольких сотен), которые теряют в ядре (в интерфазе, т. е. между митотическоми делениями) клетки свою компактную форму, разрыхляются и заполняют объем ядра в виде хроматина. Несмотря на деконденсированное состояние, каждая хромосома занимает в ядре строго определенное положение и связана с ядерной оболочкой посредством ламины. Строго закреплены на внутренней поверхности оболочки ядра такие структуры хромосом, как центромеры и теломеры. На определенной стадии жизненного цикла клетки, в синтетическом периоде, происходит репликация, т. е. удвоение всей ДНК ядра, и хроматина становится в два раза больше. Белки, необходимые для этого процесса, поступают, конечно, из цитоплазмы через ядерные поры. Таким образом, клетка готовится к предстоящему клеточному делению — митозу, когда общее количество ДНК в ядре вернется к первоначальному уровню.
Реализация генетической информации, заключенной в ДНК в виде генов, начинается с транскрипции, т. е. с синтеза информационных РНК (и-РНК) — точных копий генов, по которым затем будут строиться в цитоплазме на рибосомах белки. Этот процесс проходит в различных точках в обьеме ядра, морфологически ничем не отличающихся от окружающего хроматина. Чаще всего удается наблюдать транскрипцию диффузного, т.е. деконденсированного хроматина.
Кроме хроматина, составляющего хромосомы, в ядрах эукариот обычно содержится одно или несколько ядрышек. Это плотные структуры, не имеющие собственной оболочки и представляющие собой скопления молекул другого типа РНК — рибосомной РНК (р-РНК) в комплексе с белками. Такие комплексы называют рибонуклеопротеидами (РНП). Ядрышки имеют стандартную морфологию и образуются в ядре после деления клетки вокруг постояннодействующих точек активного синтеза рибосомной РНК. Гены рибосомной РНК, в отличие от большинства других генов, кодирующих белки, содержатся в геноме в виде многочисленных копий. Эти копии, расположенные в молекуле ДНК тандемно, т. е. друг за другом, располагаются в определенных районах нескольких хромосом генома. Такие районы хромосом называют ядрышковыми организаторами. Морфологически в ядрышке с помощью электронного микроскопа можно выделить следующие 3 зоны: гомогенные компактные фибриллярные центры, содержащие ДНК ядрышковых организаторов; плотный фибриллярный компонент вокруг них, где идет транскрипция генов рибосомной РНК и массивный гранулярный компонент ядрышка, состоящий из частиц РНП — будущих рибосом. Эти гранулы РНП, образующиеся в ядрышке, транспортируются в цитоплазму и образуют рибосомы, осуществляющие синтез всех белков клетки. Третий основной тип клеточных РНК — мелкие транспортные РНК — транскрибируются в различных участках ядра и выходят в цитоплазму через ядерные поры. Там они, как известно, обеспечивают транспортировку аминокислот к рибосомам в процессе синтеза белков.
Ядерный белковый матрикс
Для осуществления процессов репликации, транскрипции, а также поддержания определенного положения хромосом в обьеме ядра существуют каркасные белковые структуры, называемае ядерным белковым матриксом. Такой матрикс состоит, по крайней мере из трех морфологических компонентов: периферического фиброзного слоя- ламины; внутреннего, или интерхроматинового матрикса ядра и матрикса ядрышка. Наблюдения показывают, что компоненты ядерного матрикса — это не жесткие застывшие структуры, они динамичны и могут сильно видоизменяться в зависимости от функциональных особенностей ядер. Показано, что белковый матрикс имеет множество точек прочного связывания с ДНК ядра, которая, в свою очередь, имеет специальные последовательности нуклеотидов, необходимые для этого.
Строение и функции эндоплазматической сети: гладкая и шероховатая ЭПС
Прежде чем перейти к строению и функциям ЭПС, дадим ее определение.
Что такое ЭПС в биологии?
Эндоплазматическая сеть, а также ЭПС или эндоплазматический ретикулум — сложная ультрамикроскопическая разветвленная и взаимосвязанная система мембран, относительно равномерно пронизывающая цитоплазматическую массу всех эукариотических клеток.
Что такое ЭПС теперь понятно. Вот как выглядит эндоплазматическая сеть на рисунке:
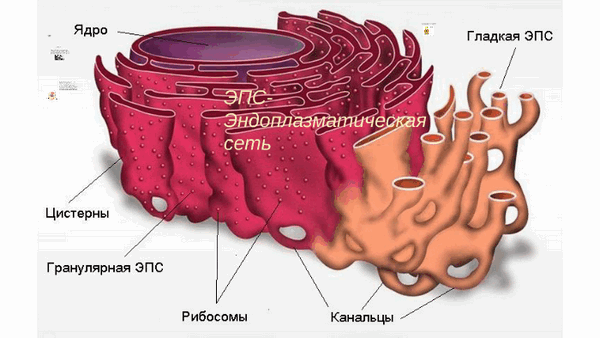
На рисунке ЭПС видно, из чего она состоит. Также рисунок ЭПС демонстрирует два вида ЭПС, о которых подробнее будет написано ниже.
Описание строения и функций ЭПС нужно начинать с того, что ЭПС — это мембранная органелла, которая включает в себя плоские мембранные мешочки: цистерны, каналы и трубочки. За счет такого строения ЭПС способствует существенному увеличению площади внутренней клеточной поверхности и делению клетки на секции. Строение эндоплазматической сети предполагает, что внутри клетки находится матрикс, представляющий собой умеренно плотный и рыхлый материал, то есть, продукт синтеза.
В каждой из секций клетки содержится различное количество химических веществ. По этой причине химические реакции в незначительном объеме клетки могут происходить одновременно или в определенной последовательности.
Особенность строения эндоплазматической сети — это ее открытие в перинуклеарное пространство, которое представляет собой полость, находящуюся между двух мембран кариолемы.
Еще один важный момент, касающийся строения ЭПС, заключается в том, что ее мембрана состоит из белков, липидов (в большей степени из фосфолипидов) и ферментов (аденозинтрифосфатаза, ферменты синтеза мембранных липидов).
Есть 2 вида ЭПС:
- Гладкая ЭПС или агранулярная аЭС. Гладкая эндоплазматическая сеть представлена трубочками: они анастамозируют между собой, у них нет на поверхности рибосом.
- Шероховатая ЭПС или гранулярная ЭПС,грЭС. Шероховатый эндоплазматический ретикулум состоит из цистерн, соединенных между собой и покрытых рибосомами.
В некоторых случаях выделяют переходящую или транзиторную эндоплазматическую сеть (тЭС). Она размещается в месте перехода одного вида ЭС в другой.
Гранулярная эндоплазматическая сеть характерная для всех клеток за исключением сперматозоидов. Степень развития этой сети зависит от специализации клетки.
Эндоплазматическая сеть в клетках эпителиальных железистых (печени — ее клетки синтезируют альбумины сыворотки крови, поджелудочной железы — ее клетки вырабатывают пищеварительные ферменты), фибробластах (клетки соединительной ткани — продуцируют белок коллаген), плазматических клетках (производят иммуноглобулины) развита очень сильно.
Агранулярная ЭС характерна для клеток надпочечников (они синтезируют стероидные гормоны), клеток мышц (они участвуют в обмене кальция) и клеток фундальных желез желудка (они работают над выделением ионов хлора).
Еще одни вид мембран цитоплазматической сети — разветвленные мембранные трубочки. Внутри них находится множество специфических ферментов, а также везикулы, которые представляют собой небольшие пузырьки, окруженные мембраной, чаще всего находящиеся около трубочек и цистерн. Их роль — обеспечение переноса синтезируемых веществ.
Это что касается особенностей строения эндоплазматической сети.
Теперь перейдем к функциям ЭПС.
Функции эндоплазматической сети
Говоря о строении и функциях эндоплазматической сети, важно напомнить следующее.
Эндоплазматический ретикулум — это аппарат синтеза и транспорта цитоплазматических веществ (в некоторой степени), за счет которого клетка может выполнять достаточно сложные функции.
К функциям ЭПС обоих видов относится все, что связано с синтезом и транспортом веществ. Что такое эндоплазматическая сеть в этом случае? Ретикулум — это универсальная транспортная система. Поэтому неудивительно, что выделяют определенные функции эндоплазматического ретикулума.
Общих функций у эндоплазматической сети обоих видов немало.
Благодаря своему содержимому (матриксу) и мембранам обе ЭПС в клетке выполняют общие функции.
Функции гладкой ЭПС и функции шероховатой ЭПС:
- разделительная или структурирующая. Благодаря ей происходит упорядоченное распределение цитоплазмы: она не смешивается. Кроме того, разделительная функция эндоплазматической сети препятствует попаданию случайных веществ в органеллу;
- трансмембранного транспорта. За счет этой функции необходимые вещества переносятся сквозь стенку мембраны;
- синтеза липидов мембраны при участии ферментов. Эти ферменты находятся в самой мембране и обеспечивают процесс репродукции эндоплазматической сети;
- обеспечение проведения импульсов возбуждения. Это возможно за счет разницы возникающих между двумя поверхностями мембран ЭС потенциалов.
Какие функции выполняет эндоплазматическая сеть в растительной клетке? В растительной клетке эндоплазматическая сеть выполняет функцию синтеза провакуолей, которые обеспечивают жизнь растительной клетки.
У каждого вида ЭПС есть свои специфические функции, которые зависят от строения и функций эндоплазматической сети в целом.
Функции гладкой ЭПС (агранулярной)
Гладкий эндоплазматический ретикулум помимо тех функций, что были перечислены выше, выполняет еще кое какие специфические функции:
- депо кальция. В скелетных мышцах, яйцеклетках, нейронах, в сердце и других клетках есть механизмы, благодаря которым изменяется концентрация ионов кальция. В поперечнополосатой мышечной ткани есть специальная ЭПС — саркоплазматический ретикулум. Он представляет собой резервуар кальций-ионов, а его мембраны имеют мощные кальциевые помпы, за счет которых в цитоплазму выбрасывается много кальция, а также происходит мгновенная транспортировка кальция в полости каналов сети;
- синтез липидов. Это вещества вроде холестерина и стероидных гормонов. Основные места синтеза стероидных гормонов — эндокринные клетки половых желез и надпочечников, клетки почек и печени. Синтез липидов осуществляют клетки кишечника: после этого липиды выводятся в лимфу и кровь;
- детоксикация. Заключается в обезвреживании экзогенных и эндогенных токсинов. К примеру, почечные клетки (гепатоциты) содержат ферменты оксидазы, разрушающие фенобарбитал;
- синтез гликогена. Он происходит в клетках печени с участием ферментов органлелл.
Функции шероховатой ЭПС (гранулярной)
Для гранулярной эндоплазматической сети характерны следующие функции:
- синтез белков. Эта функция ЭПС отличается некоторыми особенностями. В частности, начало синтеза происходит на свободных полисомах: в последующем они связываются с мембранами ЭС;
- осуществление синтеза всех белков клеточной мембраны, за исключением гидрофобных белков, а также белков внутренних мембран митохондрий и хлоропластов; специфических белков внутренней фазы мембранных органелл, секреторных белков, транспортируемых по клетке и поступающих во внеклеточное пространство;
- посттрансляционная модификация белков. Здесь имеются в виду процессы гидроксилирования, сульфатирования, фосфориллирования. Стоит также выделить процесс гликозилирования, осуществляющийся под влиянием фермента гликозилтрансферазы, связанного с мембраной. Гликозилирование осуществляется до момента секреции или транспортировки веществ к некоторым участкам клетки, к примеру, к лизосомам, плазмолемме, комплексу Гольджи;
- транспорт веществ, осуществляемый по внутримембранной части сети. Белки, прошедшие синтез, перемещаются по промежуткам эдндоплазматической сети в направлении комплекса Гольджи (он выводит вещества из клетки);
- участие в образовании комплекса Гольджи.
Множество функций ЭПС имеет отношение к транспорту белков, синтез которых осуществляется в рибосомах (они расположены на поверхности ЭПС). Белки после синтеза перемещаются внутрь сети, затем скручиваются и получают, таким образом, третичную структуру.
В процессе транспортировки к цистернам белок существенно изменяется. В некоторых случаях, к примеру, происходит его фосфориллирование или превращение в гликопротеин. Привычный путь для белка пролегает через зернистую ЭПС в аппарат Гольджи. Отсюда у него есть три варианта: выйти наружу клетки, поступать к другим органеллам той же клетки (к лизосомам) или отложиться как запасные гранулы.
Зернистая и незернистая эндоплазматическая сетка участвуют в клетках печени в детоксикации ядовитых веществ, которые после этого успешно выводятся из клетки.
У эндоплазматической сетки, как и у внешней плазматической мембраны, наблюдается избирательная проницаемость. В результате концентрация веществ внутри и снаружи каналов сетки получается неодинаковой. Этот момент важен для функции клетки.
Эндоплазматическая сетка мышечных клеток содержит больше ионов кальция, чем ее цитоплазма. Ионы кальция, покидая каналы эндоплазматической сетки, запускают процесс, связанный с сокращением мышечных волокон.
Ферменты самой сети синтезируют липидные компоненты мембран ЭПС, а белковые компоненты поступают из рибосом, которые находятся на ее мембранах. Гладкая ЭПС не обладает собственными факторам синтеза белка. Принято считать, что образование этой органеллы происходит как результат потери гранулярной ЭПС рибосом.
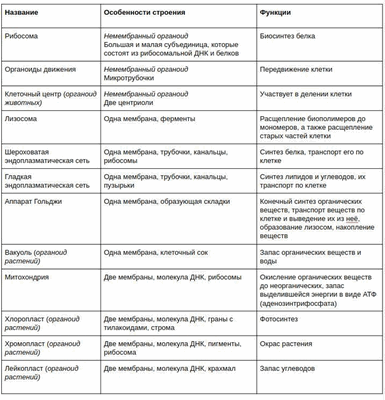
Строение и функции эндоплазматической сети в таблице (и других органоидов клетки):
Читайте также:
- Ингибиторы синтеза ранних белков вирусов. Ингибиторы синтеза нуклеиновых кислот вирусов.
- Вульгарная пузырчатка у ребенка
- Артериальные коллатерали локтевой области. Коллатеральное кровообращение в локтевой области. Анастомозы в области локтевого сустава.
- Советы при повреждении прямой кишки
- Расстройство адаптации: случай успешного лечения подростка с сахарным диабетом