Фотосистемы. Биохимия фотосинтеза. Источник кислорода.
Добавил пользователь Morpheus Обновлено: 28.01.2026
Мы знаем, что растение потребляет воду и солнечный свет, а в результате производит кислород. Но как это происходит? Чтобы понять, нужно рассмотреть процесс с точки зрения не только биологии, но и химии и физики. Каждая наука раскрывает свою сторону процесса, но, только объединившись, они дают целостную картину фотосинтеза.
Процесс фотосинтеза включает 2 фазы: световую, которая проходит на мембранах тилакоидов в присутствии света и темновую, которая не зависит от света и протекает в строме хлоропласта. В фотосинтезе участвуют многочисленные пигменты (особенно хлорофилл). В результате перемещения электронов по цепочке переносчиков запасается энергия в макроэргических связях АТФ, которая в последствии затрачивается на синтез углеводов в цикле Кальвина.
В виде побочного продукта фотосинтеза выделяется кислород, который используется многими организмами для дыхания. Благодаря ему, наша планета защищена от губительного воздействия ультрафиолетовых лучей. В противном случае, УФ- излучение способно вызвать у человека ожоги кожи и роговицы, генетические мутации. Фотосинтез обеспечивает питанием огромное количество живых организмов.
Состоит из черешка и листовой пластинки; выполняет функцию газообмена, транспирации, гуттации, фотосинтеза.
Процесс потребления организмом питательных веществ для поддержания нормального функционирования физиологических процессов
Пигменты, участвующие в фотосинтезе
Фотосинтез может осуществляться только с помощью определенных веществ — пигментов. Фотосинтетические пигменты высших растений делятся на две группы: хлорофиллы и каротиноиды.
Локализация
Пигменты локализованы в мембранах хлоропластов. В хлоропласте содержится около 400 молекул хлорофилла. Хлоропласты обычно располагаются в клетке так, чтобы их мембраны находились под прямым углом к источнику света, что гарантирует максимальное поглощение света (они могут перемещаться в клетке, в зависимости от того, как падает свет).
Хлорофилл
У растений в фотосинтезе участвует пигмент хлорофилл, который содержится в хлоропластах на мембранах тилакоидов. Хлорофилл придает хлоропластам и всему растению зеленую окраску. Хлорофилл обладает уникальным свойством: он умеет поглощать энергию солнечного света, переходя в возбужденное состояние.
Хлорофиллы поглощают главным образом красную и сине-фиолетовую часть спектра. Зеленую часть спектра они отражают и потому придают растениям характерную зеленую окраску, если только ее не маскируют другие пигменты.
Пигменты содержатся не только в клетках высших растений, но и в клетках водорослей. В клетках водорослей отдела Зеленых преобладает пигмент хлорофилл. Именно поэтому данный тип водорослей окрашен в различные оттенки зеленого.
В красных водорослях очень много пигмента фикоэритрина, характеризующегося красным цветом. Этот пигмент и придает данному отделу этих растений соответствующий цвет.
В бурых водорослях присутствует пигмент фукоксантин - бурого цвета.
То же самое можно сказать о водорослях других цветов - желто-зеленых, сине-зеленых. В каждом случае цвет определяется каким-то пигментом или их сочетанием.
Каротиноиды поглощают солнечный свет (особенно в коротковолновой — сине-фиолетовой — части спектра) и поглощенную энергию передают хлорофиллу, а также защищают хлорофилл от избытка света и от окисления кислородом, выделяющимся при фотосинтезе.
Каротиноиды постоянно присутствуют в листьях, но незаметны из-за присутствия хлорофилла. Зато осенью, когда хлорофилл разрушается, каротиноиды становятся хорошо видны. Именно они придают листьям желтую и красную окраску.
Универсальность молекулярного состава организмов
Биологические молекулы часто бывают универсальны и встречаются у совершенно разных организмов. Такая структура как молекула хлорофилла встречается у растений, а также у некоторых бактерий, но близкая ей по химическому составу молекула есть и у нас с вами. Это гемоглобин - железосодержащий белок крови животных. Различаются эти молекулы только центральными атомами железа, от которых, главным образом, и зависят основные функции этих веществ.
В зависимости от пигментного состава, водоросли могут поглощать солнечную энергию на разной глубине. Таким образом, увидев водоросль на картинке или в магазине, вы сможете сказать примерную глубину, на которой эта водоросль могла расти. Например, зеленые водоросли распространены на глубине до 30 м, поскольку более активно поглощают красный свет.
Красные водоросли, живущие на глубине от 50 до 200 м, осуществляют фотосинтез, используя слабый голубоватый зеленый свет, который проникает через толщу воды.
Световая фаза
Хлорофиллы объединяются в фотосистемы. Каждая фотосистема состоит из светособирающего комплекса, реакционного центра и переносчиков электронов. В них происходит возбуждение электрона, переход его по цепочке переносчиков, синтез АТФ и выделение О2 как побочного продукта . Световая фаза протекает на мембранах тилакоидов в хлоропластах исключительно при участии света.
Возбуждение хлорофиллов и перемещение электрона
Есть два типа фотосистем. Различаются они тем, что принимают разную длину волны света. Фотосистема II поглощает волны 680 нм за счет хлорофилла P680, а фотосистема I - свет длиной 700 нм за счет хлорофилла P700.
Молекулы хлорофилла фотосистем поглощают квант света. Один электрон каждой из них переходит на более высокий энергетический уровень (возбуждается) и перемещается далее по цепи переносчиков.
Молекулы хлорофилла, потерявшие электроны, становятся нестабильными и стремятся заполнить образовавшуюся «дыру». Фотосистема I восполняет потерю электронов через систему переносчиков электронов от фотосистемы II. Фотосистема II забирает электрон у воды в процессе фотолиза.
Фотолиз воды и образование кислорода
Одновременно с работой фотосистем происходит процесс распада воды под действием солнечного света. Этот процесс называется фотолиз.
В результате фотолиза образуются свободные электроны, кислород, ионы водорода. Электроны восполняют фотосистему II. Кислород выделяется в атмосферу. Ионы водорода накапливается в полости тилакоидов.
В полости тилакоида накапливается большой избыток ионов водорода, что приводит к созданию на мембране тилакоида градиента концентрации этих ионов.
Синтез молекул АТФ и образование НАДФ*2Н
Электрон проходит по цепи переносчиков (белков мембраны хлоропласта). В процессе этого перехода от одного переносчика к другому, электрон выделяет энергию, которая впоследствии тратится на синтез АТФ из АДФ и неорганического фосфата (Фн).
Градиент используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфата. Происходит перенос ионов водорода через мембрану восстановленным переносчиком НАДФ (никотинамидадениндинуклеотид-фосфатом) с образованием НАДФ*Н.
Далее этот водород, который присоединил к себе НАДФ, тратится на синтез углеводов в цикле Кальвина в темновой фазе фотосинтеза.
Таким образом, энергия света запасается в световой фазе фотосинтеза в виде двух типов молекул: восстановленного переносчика НАДФ*Н и макроэргического соединения АТФ.
Открытие фотосистем
Роберт Эмерсон путем опытов с 1942 по 1957 год анализировал влияние длины световой волны на квантовый выход фотосинтеза у одноклеточной водоросли хлореллы, то есть количество кислорода, выделившегося в процессе фотосинтеза в расчете на 1 квант поглощенной энергии.
Эмерсон установил, что у хлореллы наиболее эффективным для фотосинтеза был красный свет с длиной волны от 650 до 680 нм и синий свет с длиной волны от 400 до - 460 нм. Именно этот свет поглощается хлорофиллом. Он также вычислил, что фотосинтетическая эффективность красного света была на 36% выше, чем синего.
В следующих опытах было показано, что если клетки освещать красным светом с длиной волны от 650 до 680 нм, то квантовый выход достаточно высок.
Однако при дальнейшем увеличении длины волны света свыше 685 нм, квантовый выход фотосинтеза резко падает.
Если же хлореллу освещать и коротковолновым (650 нм) и длинноволновым (700 нм) красным светом, суммарный эффект будет больше, чем при действии каждого луча в отдельности. Это явление получило название эффекта усиления Эмерсона и дало возможность Эмерсону предположить, что в растениях существуют две фотосинтезирующие системы, которые должны работать согласованно.
Предположение Эмерсона легло в основу современной модели Z-фотосинтеза и дало толчок к исследованию физических, химических и функциональных свойств фотосистемы I и фотосистемы II.
Фотосинтез
Фотосинтез — это преобразование энергии света в энергию химических связей органических соединений.
Фотосинтез характерен для растений, в том числе всех водорослей, ряда прокариот, в том числе цианобактерий, некоторых одноклеточных эукариот.
В большинстве случаев при фотосинтезе в качестве побочного продукта образуется кислород (O2). Однако это не всегда так, поскольку существует несколько разных путей фотосинтеза. В случае выделения кислорода его источником является вода, от которой на нужды фотосинтеза отщепляются атомы водорода.
Фотосинтез состоит из множества реакций, в которых участвуют различные пигменты, ферменты, коферменты и др. Основными пигментами являются хлорофиллы, кроме них — каротиноиды и фикобилины.
В природе распространены два пути фотосинтеза растений: C3 и С4. У других организмов есть своя специфика реакций. Все, что объединяет эти разные процессы под термином «фотосинтез», - во всех них в общей сложности происходит преобразование энергии фотонов в химическую связь. Для сравнения: при хемосинтезе происходит преобразование энергии химической связи одних соединений (неорганических) в другие — органические.
Выделяют две фазы фотосинтеза — световую и темновую. Первая зависит от светового излучения (hν), которое необходимо для протекания реакций. Темновая фаза является светонезависимой.
У растений фотосинтез протекает в хлоропластах. В результате всех реакций образуются первичные органические вещества, из которых потом синтезируются углеводы, аминокислоты, жирные кислоты и др. Обычно суммарную реакцию фотосинтеза пишут в отношении глюкозы — наиболее распространенного продукта фотосинтеза:
Атомы кислорода, входящие в молекулу O2, берутся не из углекислого газа, а из воды. Углекислый газ - источник углерода, что более важно. Благодаря его связыванию у растений появляется возможность синтеза органики.
Представленная выше химическая реакция есть обобщенная и суммарная. Она далека от сути процесса. Так глюкоза не образуется из шести отдельных молекул углекислоты. Связывание CO2 происходит по одной молекуле, которая сначала присоединяется к уже существующему пятиуглеродному сахару.
Для прокариот характерны свои особенности фотосинтеза. Так у бактерий главный пигмент — бактериохлорофилл, и не выделяется кислород, так как водород берется не из воды, а часто из сероводорода или других веществ. У сине-зеленых водорослей основным пигментом является хлорофилл, и при фотосинтезе выделяется кислород.
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H2O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · H2 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе - солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C3-фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза - C4, также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO2. Темновая фаза протекает в строме хлоропласта.
Восстановление CO2 происходит за счет энергии АТФ и восстановительной силы НАДФ·H2, образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
Первая реакция темновой фазы - присоединение CO2 (карбоксилирование) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат) - РиБФ. Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско.
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) - первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ), включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · H2. ТФ — первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO2. Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ. Поскольку ТФ — это трехуглеродный сахар, а РиБФ — пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется. А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · H2, которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) — конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза - это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат, который превращается в глюкозу. В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фосфогликолат — это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
2 Гликолат (С2) → 2 Глиоксилат (С2) →2 Глицин (C2) - CO2 → Серин (C3) →Гидроксипируват (C3) → Глицерат (C3) → ФГК (C3)
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
C4-фотосинтез, или цикл Хэтча-Слэка
Если при C3-фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C4-пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
С4-фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
С4-растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
Растения, в которых темновая фаза фотосинтеза протекает по C4-пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой — обкладка проводящего пучка. Наружный слой — клетки мезофилла. Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C3-растений. То есть C4-путь дополняет, а не заменяет C3.
В мезофилле CO2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO2, чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание. Таким образом, преимущество C4-фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO2 в хлоропластах клеток обкладки уходит на обычный C3-путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C4-путь возник в эволюции позже C3 и во многом является приспособлением против фотодыхания.
Первичным источником энергии на Земле является энергия Солнца. Диапазон солнечного излучения, достигающего земной поверхности, называется видимым или белым светом; нижний предел длины волны его равен примерно 400 нм, а верхний — 700 нм. Фотосинтезирующие организмы (зеленые растения, водоросли, цианобактерии) обладают способностью улавливать кванты солнечного света и трансформировать их в полезную химическую энергию. Процесс фотосинтеза, заключительной реакцией которого является синтез углеводов из С02, может быть суммирован следующим стехиометрическим уравнением:
Таким образом, в результате фотосинтеза происходит:
- • восстановление световой энергией низкоэнергетической окисленной формы углерода (С02) в высокоэнергетическую восстановленную форму углерода в составе углеводов, которые затем используются нефотосинтезирующими организмами как источник энергии и углерода;
- • образование молекулярного кислорода; эта реакция представляет собой единственный источник кислорода на Земле.
Существуют две фазы процесса фотосинтеза — световая и темновая.
Световая фаза включает три процесса:
• начальной реакцией является фотохимический процесс окислительного расщепления воды — фотоокисления:
- • энергия высокоэнергетических электронов воды используется специализированной мембранной системой для фосфорилирования АДФ и образования АТФ в системе фотосинтетического фосфорилирования;
- • часть энергии электронов восстанавливает НАДФ* в реакции фотовосстановления:
Темповая фаза — это ферментативная утилизация и превращение С02 в углеводы:
Следовательно, НАДФН и АТФ, образующиеся в ходе световых реакций, являются метаболически используемыми восстанавливающими и энергетическими агентами в процессе фотосинтеза глюкозы из диоксида углерода в тем новой стадии.
Фотосинтез - это процесс образования в организмах органических веществ, на основе оксида углерода(1У) и водорода, продуцируемого фо- толитическим разложением воды. Фотосинтез осуществляется как в эукариотических организмах (высших зеленых растениях, водорослях), так и в прокариотах (фотосинтезирующих бактериях и сине-зеленых водорослях).
Суммарно процесс фотосинтеза можно выразить уравнением:
Чаще всего п=6, что соответствует образованию гексоз:
Некоторые фотосинтезирующие бактерии используют в качестве источника водорода вместо Н2О неорганические соединения (сероводород, газообразный водород и др.) или органические вещества (молочную кислоту, изопропиловый спирт).
С другой стороны, отдельные фотосинтезирующие организмы способны использовать вместо оксида углерода(1У) другие акцепторы молекулы водорода. Так, азотфиксирующие организмы используют для этих целей молекулярный азот (N2), который в процессе синтеза восстанавливается до аммиака.
Фотосинтез - это основной источник органических соединений и молекулярного кислорода на Земле. Энергия солнца аккумулируется в химических связях углеводов и в макроэргических связях АТФ, образующегося в процессе фотохимического фосфорилирования.
Фотосинтетический аппарат локализован в особых органеллах клетки - хлоропластах (хлорофилловых зернах). Хлоропласты высших растений представляют собой дискообразные тела (1-10 мк), наполненные пластинками, сгруппированными в граны. Гомогенная основа, лежащая между пластинками, называется матриксом (стромой).
Хлоропласты содержат от 58 до 75% воды. Сухое вещество хлоропласта состоит из белков (37—47%), липидов (29-36%;), углеводов (до 30%) и пигментов (7%). В хлоропластах имеются также ДНК и РНК.
В матриксе, в который по сути погружены граны хлоропластов, содержится целый ряд водорастворимых ферментов, обеспечивающих превращения продуктов фотосинтеза.
Фоточувствительным элементом хлоропласта являются пигменты, заключающие в себе хлорофиллы и каротиноиды (каротины и ксантофиллы). Их способность поглощать кванты света определенной длины волны в видимой области электромагнитного спектра обусловлена наличием в соответствующих молекулах мощных сопряженных систем (ароматических колец порфирина, полиеновых фрагментов и др.).
В зеленых растениях хлорофилла обычно значительно больше, чем каротиноидов - их соотношение иногда достигает значения 5:1. Пластиды, не имеющие хлорофилла и содержащие лишь каротиноиды
(в желтых плодах, корнях, цветах), называют хромопластами. Пластиды бесцветных тканей растений, например, клубней картофеля, в которых пигменты отсутствуют, называют лейкопластами. Между тремя видами пластид имеются переходные структуры.
Хлорофиллы являются комплексными соединениями - порфири- натами магния(II), обнаруживающими сходство с порфиринатами жс- леза(Н) (гемами). Известно несколько модификация порфиринатов магния (а, в, с, Сг, d, е), которые отличаются природой заместителей в кольце порфирина. Во всех растениях обнаруживается хлорофилл а:
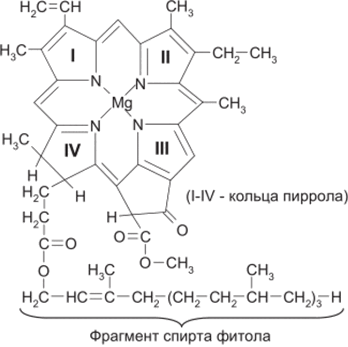
Хлорофилл а существует в растениях в виде различных форм, что связано с агрегацией его молекул и комплексообразованием с белками. Эти формы отличаются но положению максимума основной полосы поглощения в интервале 660-705 нм.
Остаток длинноцепного непредельного спирта - фитола - придает молекуле хлорофилла липофильные свойства, т.е. способность растворяться в жирах.
В качестве вспомогательного хлорофилла в высших растениях
и зеленых водорослях функционирует хлорофилл в, отличающийся
тем, что вместо метильного радикала во втором пиррольном кольце он
содержит формильную группу -с?°.
Поглощение хлорофиллом световой энергии обеспечивает комплекс антенны - две дополняющие друг друга фотосистемы (I и II), работающие на различных максимумах поглощения (700 и 680 нм).
Поглощенная световая энергия передается реакционному центру, содержащему хлорофилл а. Вместе они образуют так называемую фото- сннтетическую единицу.
Структура электронотранспортных систем фотосинтеза в хлоро- пластах подобна структуре дыхательной цепи митохондрий. В этих системах содержатся цитохромы в- и с-типа; хиноны (пластохиноны); медьсодержащий белок - пластоцианин; белок, содержащий негемовое железо - ферредоксин (Fe-S-белок); НАД-редуктаза и др.
Фотосинтез представляет собой цепь окислительновосстановительных реакций и состоит из двух стадий, или фаз - световой и темповой.
Световую фазу, связанную с использованием солнечной энергии, можно разделить на два этапа. Первый из них заключается в фотолизе (фотохимическом разложении) воды при участии хлорофилла с выделением молекулярного кислорода. Второй этап световой фазы, в котором принимают участие хлорофилл, цитохромы и другие переносчики электронов, сводится к переносу электронов от воды к НАДО и образованию АТФ.
Далее процесс фотосинтеза находит продолжение в темповой фазе. Здесь образовавшиеся 2 молекулы НАДФ-Нг и 3 молекулы АТФ используются для восстановления одной молекулы оксида углсрода(1У) до углевода. Восстановление одной молекулы ССЬ до углевода и выделение одной молекулы СЬ требует не менее восьми квантов световой энергии, т.е. квантовый выход фотосинтеза составляет 1/8 (12%).
Какие же продукты образуются в результате темнового этапа фотосинтеза - ассимиляции оксида углерода(1У)?
Характер биохимических превращений соединений углерода при фотосинтезе отражает цикл Кальвина (схема 7.11). С использованием меченного оксида углерода ( |4 СО,) было установлено, что первым продуктом фотосинтеза является фосфоглицериновая кислота.
ССЬ сочетается с фосфорилированной пентозой - рибулозо-1,5- дифосфатом и образующиеся при этом метаболиты Су. фосфоглицери- новый альдегид и фосфодиоксиацетон - приводят к образованию 1,6-дифосфатфруктозы. Последняя, в свою очередь, является источником биосинтеза сахарозы и полисахаридов (схема 7.11).
Суммарное уравнение цикла Кальвина:
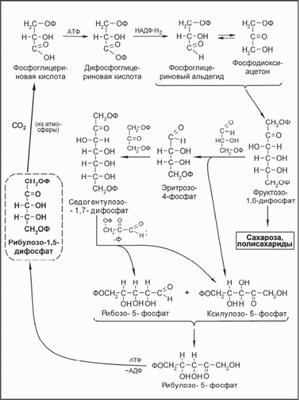
Схема 7.11. Биохимические превращения оксида углерода(1У) и рибулозо- 1,5-дифосфата в условиях фотосинтеза (цикл Кальвина).
Характерно, что при низких концентрациях ССЬ, приближающихся к атмосферным (0,03%), в хлоропласте иод действием рибулозо- дифосфаткарбоксилазы образуется фосфогликолевая кислота. Последняя выходит из хлоропласта и превращается в гликолевую кислоту, которая в особой субклеточной структуре - пероксисоме - окисляется с участием кислорода в глиоксиловую кислоту и претерпевает ряд превращений:
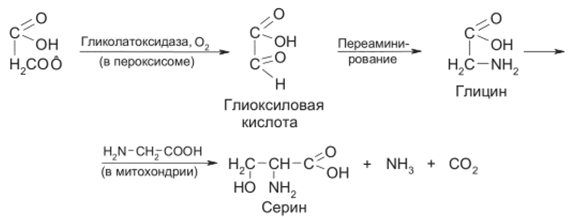
Освобождающийся аммиак связывается в глютамин
Как видно, пероксисомы и митохондрии принимают участие в процессе так называемого фотодыхания - стимулируемого светом поглощения кислорода и выделения СОг. В результате фотодыхания может значительно (до 30%) снижаться продуктивность растений.
Усвоение ССЬ в процессе фотосинтеза происходит путем карбок- силирования не только выше представленного рибулоздифосфата, но и других органических соединений, например, фосфоенолпирувата:
У некоторых фотосинтезирующих бактерий при участии восстановленного фсрредоксина оксид углсрода(1У) может ассимилироваться за счет реакций с ацетил- и сукцинил-КоА:
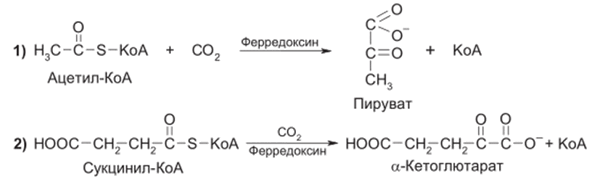
У пурпурных бактерий в условиях фотосинтеза оксид углеро- да(1У) может восстанавливаться сероводородом до формальдегида:
У бактерий этого типа (Chromatium) при коротких экспозициях света (~30 секунд) около 45% углерода ССЬ включается в аспаргино- вую кислоту через показанный выше биосинтез оксалоацетата, который далее подвергается переаминированию:
Биохимия фотосинтеза
Фотосинтез — важнейшая составляющая часть Жизни растений. В отличие ог животных, которые используют уже готовые источники пополнения биологических запасов органических веществ, растения создают их сами, используя простые неорганические соединения (С02, Н20), соединения металлов и солнечную энергию. Из молекул С02 строятся углеродные цепи углеводов и всех необходимых для жизни органических веществ. Источником водорода для всех этих соединений является вода. Создание молекул органических соединений из С02 и Н20, наиболее стабильных продуктов их полного окисления, совершается в сложнейшем биологическом аппарате зеленого листа растений, называемом аппаратом фотосинтеза, и происходит по простой суммарной схеме реакции фотосинтеза:
Эта реакция является обратной реакции окисления глюкозы, как источника энергии в клетках растений и животных. Фотосинтез не может проходить самопроизвольно, без наличия хлорофилла (см. рис. 1.1). Хлорофилл (MgXji) можно назвать фогобиокатализатором, так как он связывает воедино частицы неживой и живой природы и электромагнитное излучение и сам не изменяется в ходе фотохимического процесса.
По современным представлениям фотосинтез в зеленом листе — это сложнейший физический, химический и биологический процесс окислительно-восстановительного превращения Н20 и С02 в углеводы и другие органические соединения, инициируемый хлорофиллом (а) в фотосинтетическом аппарате. Фотосннтетический аппарат — это самонастраивающаяся, саморе- гулнруемая биологическая структура, возникающая в белково-липидных мембранах зеленого листа в результате идеальной пространственной «подгонки» (подстройки друг к другу) всех участников фотосинтеза: MgXa (а), С02, Н20, молекул акцепторов (окислителей) и доноров (восстановителей) электрона, а также ферментов, витаминов, источников энергии (АТФ) и других молекул и ионов за счет универсального и специфического взаимодействия, а также за счет экранирования тех или иных реакционных центров.
Первичным восстановителем в фотосинтезе является Н20, которая разлагается на хлорофилл—белковом комплексе, находящемся в возбужденном высокоэнергетическом состоянии, до 02 и 2Н + . Различают световую и темповую стадии фотосинтеза. Первая, световая стадия, включает фотофизиче- ские и фотохимические процессы. Главный фотофизический процесс происходит с молекулой гидрата хлорофилла (акваэкстракомплекса хлорофилла
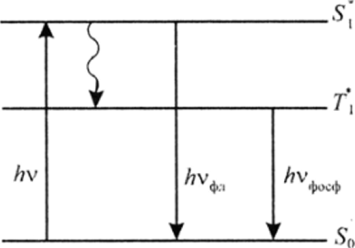
Рис. 28.5. Схема возбуждения и дезактивации хлорофилла:
/iv — квант поглощения; Ауф., — квант флюоресценции; ЛУфосФ — квант фосфоресценции; So — основное (синглетное) состояние, .Si* и Т — первые синглетное и триплетное возбужденное состояние
Мй(Н2 0)Хл) в электромагнитном поле. При этом, поглощая квант света в красной части спектра, хлорофилл возбуждается до синглетного (5,*) состояния (рис. 28.5). Время жизни этого состояния очень мало и составляет около 10 8 с.
Из нестабильного синглетного состояния хлорофилл, теряя часть энергии, быстро переходит в так называемое метасгабильное состояние с двумя неспаренными электронами в сопряженной л-системс. Это бирадикальное состояние, называемое триплетным 7*, с временем жизни тА = 0,2 с:
Из синглетного состояния хлорофилл может перейти снова в исходное состояние с испусканием кванта флуоресценции (/?уфл) или же после рассеяния части энергии в виде тепловой ( может перейти в бирадикальное
состояние 7J*. Это состояние, как предполагается является активным в фотосинтезе. В отличие от 5,* время его жизни велико, поэтому хлорофилл в состоянии Т* может вступать в самые разнообразные биохимические превращения.
В отсутствие реагентов триплетный хлорофилл испускает квант фосфоресценции (), т. е. обладает свечением после того, как убран источник облучения.
В фотосинтетическом аппарате молекула возбужденного хлорофилла может передавать энергию возбуждения другим молекулам хлорофилла, л- системам каротиноидов и фикобилинов (а также и получать энергию от их возбужденных светом молекул) или использовать ее для окисления молекулы Н20 с последующей передачей электрона от воды по цепи электронных акцепторов с постоянно снижающимся нормальным окислительно-восстано-
вительным потенциалом вплоть до передачи электрона акцептору С02 (восстановителю).
При этом не следует думать, что первичный электрон движется свободно, как по проводнику. Он химически связан и движется от соединения к соединению по очень сложной «электронно-транспортной цепи». Очень существенным фактом является то, что молекулы хлорофилла, начиная пусковую (первую) стадию фотосинтеза, взаимодействуют с окружающей средой (электромагнитное поле и скопление различных активаторов и реагентов) не в одиночку, а сгруппировавшись в фотосистемы I и II. Ориентировочно установлено, что фотосистема I, состоящая из 60—80 молекул хлорофилла (а), 15 молекул p-каротина, одной молекулы цитохрома / и двух молекул цитохрома /?6, 15 молекул пластохинона и ферментно-белкового окружения, является только фотосборщиком, т. е. она поглощает кванты света, запасает энергию солнца и передает ее затем в реакционный центр (РЦО первой фотосистемы. В этом РЦ] находятся 2—3 десятка молекул Mg)Gi (а) в форме P7(K, (700 нм — положение первой полосы поглощения X, этого типа ассоциатов хлорофилла) в белково-липидном окружении. Принято считать, что фотосистема 1 собирает кванты света в более длинноволновой части спектра (до 700 нм), а РЦ) получает энергию фотосистемы I и осуществляет темновую стадию фотосинтеза
Фотосистема II состоит из ассоциатов молекул фотосборщиков — 80 молекул хлорофилла, 50 молекул хлорофилла (b), 20 молекул ксантофилла (каротиноида, имеющего в отличие от Р-каротина еще одну 71-связь в цикле), белково-липидного окружения и реакционного центра PLfc, состоящего из 15 молекул хлорофилла (а), одного цитохрома Ь559 и ферментного окружения.
Эта фотосистема II собирает кванты света с более высокой энергией (более коротковолновые кванты до А., = 680 нм).
Считается, что только обе фотосистемы обеспечивают эффективный фотосинтез в любую светопогоду. Первая стадия фотосинтеза состоит из фото- физической и фотохимической компонент. Фотофизическая компонента детально изучена физиками-спектроскопистами на молекулах хлорофилла и его ассоциатах и на многочисленных их моделях — мсталлопорфиринах. Сложнее обстоит дело с фотохимической компонентой окисления воды, выделения кислорода и транспортировки электрона к акцепторам С02.
На механизм фотохимической стадии фотосинтеза существует множество различных взглядов, многие из которых являются механистическими, далекими от биохимии. По современным биохимическим представлениям ключевым соединением в фотосинтезе является хлорофилл. MgXn в отличие от всех других металлоаналогов хлорофилла — «металлохлорофиллов» МХл (М 2 * = Mn, Fe, Со, Ni, Си, Zn, Cd, Hg, Pd, Ag и др.) обладает оптимальным набором свойств, необходимых для фотосинтеза, и среди них одним из главных свойств — способностью прочно связывать одну молекулу воды и давать моногидрат (акваэкстракомплекс) хлорофилла:
Этот аквакомплекс достаточно прочен в ассоциатах хлорофилла и в водно-органическом окружении живой клетки зеленого листа. В этом комплексе молекула координированной воды уже частично окислена, так как элекгронная пара атома кислорода сильно смещена к Mg 2+ :
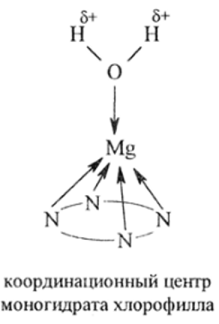
Таким образом, окисление воды триплетным хлорофиллом облегается ес координацией. Три десятилетия назад профессором В. М. Кутюриным была высказана и обоснована гипотеза, что воду окисляет 1/^Хл('л п или его
катион-радикалы 1ме формы MgXn*, Mg^ 2+ . Их окислительновосстановительный потенциал (Е° - +0,73 и +0,89 В соответственно) достаточен для окисления координированной воды. Особенно благоприятно влияет на окисление воды хлорофиллом возбуждение его электронов и донорноакцепторное окружение оболочки хлорофилла. В качестве акцепторов электрона могут выступать хиноны (пластохинон и др.), нуклеотиды, например, никотинамидадениндинуклеотидфосфат (НАДО):
Хлорофилл и его ароматическая тг-система являются хорошим резервуаром для электронов и положительных зарядов («дырок»). Впервые в 50-х годах XX столетия Л. А. Красновским было показано, что хлорофилл подвергается легкому обратимому фотовосстановлению даже такими слабыми восстановителями, как витамин С. При этом в отсутствие протонодоноров образуется первичная фотовосстановленная форма хлорофилла ( MgXa-):
Она в протонодонорной среде превращается во вторичную фотовосстанов- ленную форму хлорофилла:
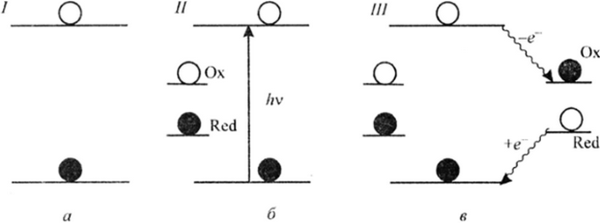
Рис. 28.6. Схема синхронного окислительно-восстановительного взаимодействия фотовозбужденного хлорофилла (по В. Б. Евстигнееву): а — основное состояние; б — возбужденное состояние; в — конечное состояние;
/—///— этапы взаимодействия хлорофилла с донором и акцептором электрона
В то же время В. Б. Евстигнеев обнаружил способность хлорофилла к легкому обратимому фотоокислению такими слабыми окислителями, как хиноны:Особенно важно, что возбужденный хлорофилл одновременно может выступать и как донор, и как акцептор электронов (рис. 28.6).
Из рис. 28.6 видно, что состояние II является одновременно хорошим восстановителем и хорошим окислителем, так как имеет вакансию (О) для электрона на ВЗМО и заполненную нестабильную ( ф ) НВМО. С вакантной орбиталью хлорофилла реагирует донор электрона (изображен темным кружком), а с заполненной орбиталью хлорофилла взаимодействует акцептор электрона (изображен светлым кружком). Из-за больших размеров л- системы хлорофилла и ее граничных орбиталей — верхней занятой (ВЗМО) и низшей вакантной (НВМО) — процесс переноса электрона от восстановителя (Red = Н20) к окислителю (Ох = хиноны, НАДФ и др.) может происходить с участием МО хлорофилла как самосогласованный тримолскулярный (а возможно — внутримолекулярный) процесс без видимого (явного) образования анион- и катион-радикальных форм как таковых. Этому благоприятствует то, что в зеленом листе существует комплекс: донор электрона — хлорофилл — акцепторы электрона, вмонтированный в белково-липидные слои мембран хлоропластов зеленого листа.
После всего сказанного фотохимическую стадию разложения воды можно представить в виде весьма вероятных гипотетических реакций:
По реакции (28.1) возникает активный электрон, носителем которого является A s , протон и г идроксил-радикал. Хлорофилл освобождается в неизменном виде. В качестве В могут выступать любые основные центры белка (фермента) из окружения хлорофилла. По реакции (28.2) происходит рекомбинация ОН* до пероксида водорода и его ферментативное разложение с выделением кислорода, поступающего в атмосферу.
Одновременно с реакцией (28.1) происходит фотофосфорилирование с превращением АДФ в АТФ с участием неорганического фосфата. Приведенная выше схема фотохимического разложения воды на хлорофилле находится в согласии с современными представлениями химии. Источником молекул Н20 в фотосинтезе может явиться присутствующий в фотосинтетическом аппарате аквакомплекс марганца [Мп(Н20)6(Н20)т], способный доставлять молекулы воды хлорофиллу, так как является одним из самых лабильных комплексов ионов 20.
Будущие исследования дадут точный ответ на вопрос о механизме световой стадии фотосинтеза, сопровождающейся выделением 02 из воды и включением электрона воды во второй гем новой процесс фотосинтеза. Несомненно одно, что хлорофиллу в этом процессе принадлежит главная роль. На тем новой стадии фотосинтеза проходят более сотни ферментативных химических реакций, каждая из которых вызывается собственным ферментом. На темновой стадии фотосинтеза важнейшую роль играют, кроме ферментов, такие биомолекулы, как НАДФ, АТФ, рибулозо-1,5-дифосфат и др. Химически неактивный (как говорят биохимики, «холодный») электрон воды превращается в химически активный («горячий») элекгрон восстановленной формы НАДФ:
При этом восстанавливается никотинамидная часть НАДФ. Далее НАДФН2 и АТФ участвуют в биохимических циклах темновых реакций, важнейшим из которых является углеводный цикл Кальвина (получившего Нобелевскую премию за этот цикл работ), который схематично может быть записан так:
Этот цикл очень сложен и состоит из таких главных стадий: 1. Стадия включения С02 рибулозо-1,5-дифосфатом:
Эта реакция относится к реакциям ферментативного деструктивного карбоксилирования, в результате чего разрывается связь С-С и внедряется молекула угольной кислоты. Эта стадия связана только с внутримолекулярным окислением-восстановлением и не требует «горячего» фотосинтетиче- ского электрона.
2. Включение в цикл четырех атомов водорода ( в расчете на две молекулы глицериновой кислоты), т. е. восстановление -СООН до -СНО с помощью НАДФН2:
3. Освобождение углевода после конденсации двух молекул фосфогли- церинового альдегида до рибулозо-5-монофосфата с отделением СН20 как структурной единицы глюкозы. Для образования глюкозы эта стадия должна повториться 6 раз:
Эта серия биохимических реакций цикла Кальвина, приводящая к синтезу глюкозы:
проходит в сложнейшем белковом (ферментном) окружении.
Далее глюкоза используется в реакции полимеризации до целлюлозы как структурного элемента растений, в реакциях синтеза многих других ор- ганических соединений в клетке растений.
Циклы восстановительного аминирования яблочной и щавелевоуксусной кислоты дают аминокислоты, необходимые для синтеза белков. Синтез липидов происходит по своей сложной биологической цепи. В заключение этого раздела следует отметить, что все современные сведения о фотосинтезе являются пока сугубо приближенными. Это вполне естественно потому, что биохимия фотосинтеза необычайно сложна.
Основной задачей при исследовании фотосинтеза на современном этапе являются: расшифровка природы всех участников электронно-транспортной цепи от Н20 до С02, установление строгой последовательности их расположения и характера донорно-акцепторных, ион-дипольных, координационных и других взаимодействий этих молекул в составе фотосинтетшюского аппарата, определение природы их связи с молекулами хлорофилла, белков и липидов мембран хлореллы. Все эти вопросы относятся к структуре фотосин- тетического аппарата, которая непосредственно определяет его функции.
Читайте также: