Изменения синапсов при развитии памяти. Нейроны и синапсы в формированиия памяти
Добавил пользователь Владимир З. Обновлено: 21.01.2026
Сообщают (Yamasaki М., Takeuchi Т., 2007; Гурская О. Я. и со-авт., 2015; Kim S., Kaang В., 2017; Albo Z., Graff J., 2018; Basu S., Lamprecht R., 2018; Nakahata Y., Yasuda R., 2018; Rodriguez-Iglesias N. et al., 2019), что в процессе консолидации долгосрочной памяти происходят существенные структурные изменения нейронов, касающиеся дендритных шипиков и синапсов. Так, отмечено, что при активации глутаматных и других синаптических рецепторов в процессе обучения создаются новые актиновые цитоскелетные каркасы, приводящие к изменениям морфологии шипиков. Это имеет важное значение для формирования долгосрочной памяти. Вновь образованный актиновый каркас цитоскелета сохраняется длительное время благодаря активной стабилизации уровня и активности белков, регулирующих сборку актиновых нитей в этих шипиках. При нарушении этих процессов следы памяти утрачиваются.
Как известно, шипики с большими постсинаптическими плотностями (PSD), имеют более высокий уровень АМРА-рецепторов, чем шипики с меньшими PSD. Чем больше площадь PSD, тем больше площадь головки шипика. Следовательно, шипики с большей головкой (грибовидной) экспрессируют больше глутаматных АМРА-рецепторов, чем шипики с меньшей головкой. Все это означает, что в процессе долгосрочной консолидации следов памяти должна возрастать площадь головок дендритных шипиков, что подтверждают исследования. Кроме того, при формировании долгосрочной памяти утолщается шейка головки дендритных шипиков, что повышает синаптические возможности.
Действительно, обучение ведет к изменению морфогенеза шипика. Например, выработка контекстного страха приводит к увеличению плотности дендритных шипиков в гиппокампе СА1 и передней поясной извилине. Закрепление слухового страха сопровождается уменьшением числа шипиков в слое V пирамидных нейронов в ассоциативных участках лобной коры у мыши, а исчезновение страха вызывает формирование шипиков в этом регионе мозга. Эти изменения стабильны, то есть сохраняются длительно — от нескольких дней до нескольких лет (особенно, если память поддерживается тренировками), и это коррелирует с длительностью хранения следов памяти.
Итак, сохранение структуры шипика необходимо для поддержания памяти, чему способствует сохранение актинового цитоскелета шипика.
Как известно, зрелые шипики содержат смесь разветвленных и линейных актиновых филаментов у основания шипика, в его шейке и в головке. Этот актиновый цитоскелет играет важную роль в стабильности шипика, его подвижности и морфологии. Форма и динамика зрелых шипиков регулируются двумя различными группами актиновых филаментов. Стабильный пул F-актина имеет скорость оборота, равную 1 мин, и находится у основания шипика, тогда как динамический пул F-актина имеет скорость оборота, равную 1 с. Изменения в структуре шипика зависят от полимеризации актина. Например, увеличение головки шипика под влиянием глутамата зависит от полимеризации актина.
Полимеризация, деполимеризация актинового цитоскелета, а также ветвление актина контролируется малыми ГТФ-азами (Rael, Cdc42 и Rho) и другими веществами, активность которых зависит от состояния глутаматных и других синаптических рецепторов. Показано, что процессы, происходящие с цитоскелетом, имеют важное значение для формирования памяти. Вмешательство в правильную полимеризацию актинового цитоскелета ухудшает формирование долгосрочной памяти. Например, удаление белка п-кофилина (n-cofilin), вызывающего деполимеризацию актиновых филаментов, приводит к угасанию памяти. Нарушение функции белков, регулирующих сборку актиновых нитей, в частности, волнового регуляторного комплекса, также нарушает процесс консолидации памяти, что установлено на крысах при их обучении в водном лабиринте Морриса.
Важно отметить, что среди регуляторов актинового цитоскелета шипиков находится мозговой нейротрофический фактор (BDNF), способный активировать малую ГТФ-азу, то есть белок Rac, поэтому недостаточная продукция BDNF может приводить к нарушению формирования актинового цитоскелета шипиков и тем самым к уменьшению эффективности долгосрочной консолидации. Недостаток продукции белка Агр2/3, необходимого для роста головки шипика, помимо нарушения морфологии шипика приводит к ослаблению способности к консолидации памяти, что установлено при исследовании рабочей памяти в Y-лабиринте, а также при исследовании способности к распознаванию нового объекта (эпизодическая память) и формированию долгосрочной памяти страха.
Многие из этих данных получены при использовании ингибиторов синтеза соответствующих белков, регулирующих формирование актинового циктоскелета, например, при использовании латрун-кулина А, который предотвращает включение G-актина в динамический F-actin, при использовании ингибиторов таких белков, как белок Rael, белок Cdc42, белок профилин, комплекс белков Агр2/3.
Каким же образом удается долгосрочно сохранять изменения актинового цитоскелета шипиков, возникшие после обучения? Для объяснения механизма стабилизации морфологии дендритных шипиков предложена гипотеза (Basu S., Lamprecht R., 2018), которая содержит ряд положений.
Первое положение: во время фазы поддержания памяти происходит спонтанная активация глутаматных АМРА-рецепторов, при которой генерируются миниатюрные потенциалы. Они помечают синапс, структуру которого требуется сохранять. Кроме того, эти миниатюрные потенциалы привлекают в синапс белки, участвующие в формировании актинового цитоскелета шипиков.
Но этого механизма недостаточно, так как обмен белков идет с огромной скоростью. Поэтому предлагается второе положение модели, согласно которому измененный при обучении актиновый цитоскелет служит каркасом, в котором новые актиновые мономеры и ядерные белки непрерывно пополняют сеть, сохраняя общую каркасную структуру нетронутой. Таким образом, быстрый оборот F-актина не влияет на структуры актиновых филаментов в шипи-ке, в связи с чем структура шипика остается стабильной. Зрелые шипики состоят из смеси разветвленных и линейных актиновых филаментов в основании, шейке и головке, которые определяют их структуру.
Третье положение утверждает, что при длительной депрессии (LTD) шипики могут сжиматься («усадка» шипиков), что способствует очищению нейронной сети, то есть торможению ненужных следов памяти. В этом случае генерация миниатюрных ПД прекращается, что уменьшает активность данного шипика. В какой степени гипотеза (Basu S., Lamprecht R., 2018) отражает реальные события — очевидно, покажут будущие исследования. Представление о важности миниатюрных потенциалов при активации АМРА-рецепторов для поддержания функции дендритного синапса поддерживают и другие авторы (Nakahata Y., Yasuda R., 2018).
Сообщают (Гурская О. Я. и соавт., 2015), что в формировании шипиков при обучении важную роль играет белок Wnt5a, как компонент сигнальной системы «Wnt — бета-катенин», о которой говорилось выше в разделе «Генетические аспекты долгосрочной консолидации памяти». Вероятнее всего, активируя транскрипционный фактор бета-катенин, белок Wnt5a инициирует появление новых шипиков и увеличивает размеры уже существующих шипиков, что повышает эффективность глутаматергических синапсов в гиппокампе и тем самым способствует консолидации долгосрочной памяти.
Сообщают (Albo Z., Graff, J. 2018), что дендритные шипики имеют различные формы: грибовидные, тонкие, короткие и разветвленные. При этом тонкие шипики предназначены для формирования краткосрочной памяти, а грибовидные шипики представляют собой субстраты долгосрочной памяти. Сообщается, что введение транскрипционного фактора MEF2, который тормозит шипикоге-нез, блокировало формирование долгосрочной памяти. Показано, что у крыс в медиальной префронтальной коре уже через 1 час после обучения возрастает доля грибоподобных шипиков по сравнению с тонкими шипиками. Это подтверждает представление о том, что долгосрочная консолидация памяти сопровождается морфологическими изменениями шипиков.
В то же время установлено, что в гиппокампе морфология шипиков, меняющаяся при выработке рефлекса, сохраняется сравнительно недолго — не более 15 дней после обучения. Эти авторы сообщают также, что при консолидации долгосрочной памяти в дендритных шипиках происходит полимеризация актина и повышение содержания кофилина, который способствует сборке цитоскелета шипиков и тем самым регулирует морфологию шипиков. Для того чтобы вновь сформированные шипики существовали длительное время, они должны быть помечены (отмаркированы), чтобы в них происходили процессы, постулируемые гипотезой (Basu S., Lamprecht R., 2018). По мнению ученых (Albo Z., Graff J., 2018), в реализации долгосрочного сохранения вновь созданных шипиков участвует и эпигенетический механизм, в том числе ацетилирование гистонов.
Сообщают (Kim S., Kaang В., 2017), что дендритные шипики обнаружены во многих областях мозга мыши, но число шипиков с возрастом снижается (а не возрастает). При этом скорость оборота шипиков в разных областях мозга различна. Показано, что у пирамидных нейронов области СА1 гиппокампа крыс при обучении меняются дендриты и образуются новые синаптические соединения, то есть дендритные шипики. Особенно это характерно при формировании памяти страха. Эти шипики могут сохраняться длительное время, что обусловлено, вероятно, эпигенетическими механизмами.
Полагают (Yamasaki М., Takeuchi Т., 2007), что при долгосрочной консолидации следов памяти повышается объем дендритных шипиков, которые метятся для того, чтобы постоянно поддерживать свою функцию. В этом помогает дофамин, активируя Dl/DS-рецепторы, что привлекает к шипику белки, участвующие в формировании ши-пика. Авторы также подчеркивают, что в геноме млекопитающих имеется 1755 генов, продукты которых имеют отношение к изменению дендритных шипиков. Все это говорит о необходимости продолжения исследований механизмов сохранения дендритных шипиков, сформированных при обучении.
Указывают (Rodriguez-Iglesias N. et al., 2019), что микроглия принимает активное участие в формировании шипиков дендритов нейронов, которые образуются в результате «взрослого» нейрогенеза в нейрогенных нишах, что позволяет этим нейронам встраиваться в нейронные сети гиппокампа без потери памяти на прошедшие события. Этому способствует прямое влияние микроглиоцитов на нейроны либо опосредованное, то есть за счет выделения БАВ, например, BDNF. При этом головки шипиков под влиянием микроглиоцитов могут образовывать филоподии, за счет которых шипики могут перемещаться вдоль дендрита, либо образовывать полисинап-тические бутоны. Часть БАВ, которые продуцируют микроглиоциты, могут повышать число дендритных шипиков нейронов (BDNF, интерлейкин-10), а часть — снижать их число (интерлейкин-1бета, ТЫРальфа). С участием микроглиоцитов происходит рост числа шипиков под влиянием физических упражнений, либо под влиянием когнитивных нагрузок. При старении, когда в мозге уменьшается число микроглиоцитов, уменьшается интенсивность взрослого нейрогенеза и способность нейронов образовывать новые дендритные шипики. Более детально о других аспектах участия микроглиоцитов и других глиальных клеток в процессах памяти сказано ниже.
Долговременная память. Роль синаптических процессов и нейропептидов в консолидации памяти
В долговременной памяти информация может сохраняться всю жизнь. Основой долговременной памяти являются структурные изменения в нейронах. В долговременную память информация начинает поступать через 45 минут от ее поступления в кратковременную память. Объем долговременной памяти практически безграничен. Она по своему механизму коренным образом отличается от кратковременной и промежуточной памяти, так как не нарушается при механических травмах мозга, электрошоке, наркозе и т.д. Долговременная память формируется на базе кратковременной и промежуточной памяти, при этом происходят устойчивые изменения в нейронах на синаптическом, молекулярном и клеточном уровнях.
Важную роль в механизмах долговременной памяти играют синаптические процессы. В ЦНС под влиянием обучения образуются новые синапсы и увеличивается их количество, повышается выделение в них медиаторов, разрастаются дендриты и увеличивается число шипиков на них, а также количество коллатералей аксонов. Прекращение обучения приводит к исчезновению этих явлений в течение месяца. В нейронах синтезируются специальные белки, стабилизирующие синаптические процессы. В результате этих изменений в структурах мозга создаются зоны облегченной синаптической проводимости, что и обусловливает формирование соответствующих энграмм памяти.
При конвергенции на одном нейроне синаптических входов от безразличного (в будущем условного) и безусловного раздражителя в процессе выработки условного рефлекса происходит следующее (рис. 110). Изначально возбуждение, поступающее в синапс (1) от безразличного раздражителя, не вызывает возникновения потенциала действия в нейроне, а генерирует только подпороговый ВПСП. Предполагается, что возбуждение, приходящее в синапс (2) от безусловного раздражителя, помимо генерации потенциала действия в нейроне-мишени ускоряет биохимические реакции в его цитоплазме (3), направленные на активацию генома ядра нервной клетки-мишени (4). Вследствие такой активации происходит изменение образования РНК и синтеза в рибосомах (5) нейрона специфического белка.Затем вновь синтезированный белок перемещается (6) в сторону постсинаптической мембраны того синапса, который входит в структуру временной нервной связи и подвергается подпороговой активации в результате поступления в него возбуждения от безразличного (в будущем условного) раздражителя. На последнем этапе белковая молекула встраивается в постсинаптическую мембрану (7) данного синапса. В результате возбудимость постсинаптической мембраны увеличивается, синапс из неэффективного превращается в эффективный, и при поступлении возбуждения от условного раздражителя нейрон начинает генерировать потенциал действия.Процесс обучения ухудшается при торможении синтеза РНК.
Большое значение в обеспечении механизмов долговременной памяти придается медиаторам. Основные медиаторные системы головного мозга - холинергическая и моноаминергическая, включающая норадренергическую, дофаминергическую и серотонинергическую - принимают самое непосредственное участие в обучении и формировании энграмм памяти. Так, экспериментально установлено, что уменьшение количества норадреналина замедляет обучение, вызывает амнезию и нарушает извлечение следов из памяти. Показано участие дофамина в механизмах памяти. Серотонин принимает участие в процессах консолидации памяти.
Нейропептиды и память.Большую роль в консолидации памяти играют нейропептиды, которые могут находиться в пресинаптических окончаниях в качестве сопутствующего медиатора. Например, вместе с ацетилхолином часто выделяется окончаниями аксонов вазоактивный интестинальный пептид (ВИП), энкефалин, люлиберин. Нейропептиды могут оказывать модулирующее действие на пре- и постсинаптическом уровне.
Сопутствующий пептид может значительно повысить чувствительность постсинаптических рецепторов к основному медиатору. Например, ВИП усиливает более чем в 10 000 раз чувствительность к ацетилхолину. Поскольку пептиды выделяются нейронами и в синаптическую щель и во внеклеточное пространство, то они могут оказывать влияние на целую группу нейронов и способствовать быстрому функциональному объединению нервных клеток в ходе обучения, в явлениях памяти.
Ставится вопрос о возможности переноса выработанных навыков от одного животного к другому с помощью образованных в ходе обучения и «отвечающих» за соответствующий навык специфических пептидов. Наибольшую популярность в связи с этим приобрели эксперименты Г. Унгара, который предпринял попытку выделить особое вещество - пептид «скотофобин» (от греч. «боязнь темноты»), образующийся в мозге при обучении животного бояться темноты. Этот пептид был синтезирован и использовался в различных лабораториях для проверки способности переноса химическим путем определенной информации от обученных животных к необученным. Однако многочисленные проверки не дали положительных результатов.
Идея существования биохимических факторов, способных к сохранению и переносу информации, большинством исследователей воспринимается критически. В настоящее время считается, что гипотеза молекулярного кодирования индивидуального опыта не имеет прямых фактических доказательств. Несмотря на то, что установлена существенная роль нуклеиновых кислот и белков в механизмах научения и памяти, предполагается, что принимающие участие в формировании новой ассоциативной связи РНК и белки специфичны лишь по отношению к функциональному изменению участвующих в процессе синапсов и неспецифичны по отношению к самой информации.
Значительный интерес для выяснения роли пептидов в процессах памяти представляют опыты с введением в мозг адренокортикотропного и меланоцитостимулирующего гормонов, вазопрессина, позволяющие судить об их способности улучшать запоминание. Обнаружено, что некоторые фрагменты АКТГ участвуют в механизмах консолидации памяти. Животные с врожденным нарушением синтеза вазопрессинане способны к выработке сложных условных оборонительных рефлексов. При дополнительном введении этим животным вазопрессина образование оборонительных навыков восстанавливается. Установлено, что у животных с дефицитом вазопрессина страдает не сам процесс обучения, а процесс запоминания информации, т.е. консолидация сформировавшихся энграмм. В отличие от вазопрессина гормон окситоцин нарушает сохранение выработанных навыков у животных. Существенно улучшают процессы памяти эндогенные опиатные пептиды - эндорфины и энкефалины.
Большинство исследователей считают, что нейропептиды участвуют в процессах обучения и памяти путем взаимодействия с медиаторами, через которые они оказывают свое регулирующее влияние.
Химические синапсы. Особенности проведения возбуждения через химические синапсы. Медиаторы.
Синапс - это специализированное структурное соединение между клетками, обеспечивающее взаимное влияние между ними. Через синапсы передаются возбуждающие и тормозные влияния между двумя возбудимыми клетками, осуществляется трофическое влияние, синапсы играют важную роль в реализации механизмов памяти.
Все синапсы классифицируются по следующим критериям:
1. По виду соединяемых клеток:
1. межнейронные - локализуются в ЦНС и вегетативных ганглиях;
2. нейроэффекторные - соединяют эфферентные нейроны соматической и вегетативной нервной системы с исполнительными клетками;
3. нейрорецепторные - осуществляют контакты во вторичных рецепторах между рецепторной клеткой и дендритом афферентного нейрона.
2. По эффекту: возбуждающие и тормозящие.
3. В зависимости от местоположения в ЦНС: аксосоматические, аксодендритные, аксоаксональные, дендросоматические и дедродендритные.
4. По способу передачи сигналов:
1. Химические - наиболее распространенные в ЦНС, в которых посредником (медиатором) передачи является химическое вещество. Химические синапсы по природе медиатора делят на холинэргические (медиатор - ацетилхолин), адренэргические( норадреналин), дофаминэргические (дофамин), ГАМК-эргические (γ-аминомасляная кислота) и т.д.
2. Электрические, в которых сигналы передаются электрическим током;
3.Смешанные синапсы - электрохимические.
Механизм синаптической передачи сигналов.
►Химические синапсы - это преобладающий тип синапсов в мозгу млекопитающих и человека. В химическом синапсе выделяют пресинаптическое окончание, синаптическую щель и постсинаптическую мембрану.
В пресинаптическом окончании находятся синаптические пузырьки - везикулы - диаметром до 200 нм, которые образуются либо в теле нейрона и с помощью аксонного транспорта доставляются в пресинаптическое окончание, либо синтезируются (или ресинтезируются) в самом пресинаптическом окончании. Везикулы содержат медиаторы, необходимые для передачи влияния одной клетки на другую. Для синтеза медиатора нужны ферменты, которые образуются в теле клетки на рибосомах и доставляются в пресинаптическое окончание аксонным транспортом. Кроме везикул с медиатором в пресинаптическом окончании имеются митохондрии, которые обеспечивают энергией процесс синаптической передачи. Эндоплазматическая сеть окончания содержит депонированныйСа + . Микротрубочки и микрофиламенты участвуют в передвижении везикул. Пресинаптическое окончание имеет пресинаптическую мембрану. Пресинаптической мембраной называют часть пресинаптического окончания, которая ограничивает синаптическую щель.
Синаптическая щель имеет ширину 20-50 нм. В ней содержится межклеточная жидкость и вещество мукополисахаридной природы в виде тяжей междупре- и постсинаптической мембранами. В синаптической щели также находятся ферменты, которые могут разрушать медиатор.
Постсинаптическая мембрана - утолщенная часть клеточной мембраны иннервируемой клетки, содержащая белковые рецепторы, имеющие ионные каналы и способные связывать молекулы медиатора. Постсинаптическую мембрану нервно-мышечного синапса называют также концевой пластинкой.
Химические синапсы. Особенности проведения возбуждения через химические синапсы. Медиаторы.
Химические - наиболее распространенные в ЦНС, в которых посредником (медиатором) передачи является химическое вещество. Химические синапсы по природе медиатора делят на холинэргические (медиатор - ацетилхолин), адренэргические( норадреналин), дофаминэргические (дофамин), ГАМК-эргические (γ-аминомасляная кислота) и т.д.
Химические синапсы - это преобладающий тип синапсов в мозгу млекопитающих и человека. В химическом синапсе выделяют пресинаптическое окончание, синаптическую щель и постсинаптическую мембрану.
В процессе передачи сигнала в химическом синапсе можно выделить следующие этапы (см. схему):
1. Потенциал действия поступает в пресинаптическое окончание.
2. После поступления ПД к пресинаптическому окончанию происходит деполяризация мембраны окончания, активируются потенциал-зависимые кальциевые каналы и в синаптическуютерминаль входит Са + .
3. Повышение концентрации ионов Са + активирует транспортную систему, что инициирует их экзоцитоз.
4. Содержимое везикул выделяется в синаптическую щель.
5. Молекулы медиатора, диффундируются в синаптической щели, связываются с рецепторами постсинаптической мембраны.
6. Рецепторы постсинаптической мембраны активируют ионные каналы.
7. В результате под действием медиатора происходит активация ионных каналов и переход по этим каналам ионов К + и Nа + по их градиентам концентрации. Движение ионов формирует постсинаптический потенциал, который по своим свойствам является локальным ответом.
8. Медиатор, находящийся в контакте с рецепторами постсинаптической мембраны и в синаптической щели, разрушается ферментами.
9. Продукты разрушения медиатора и не разрушенный медиатор всасываются преимущественно в пресинаптическое окончание, где осуществляется ресинтез медиатора и помещение его в везикулы.
На все эти процессы требуется определенное время, которое получило название синаптической задержки и составляет 0,2-0,5 мс. Синаптическая задержка пропорционально зависит от температуры.
Выделение молекул медиатора из пресинаптического окончания пропорционально количеству поступившего туда Са + в степени n = 4. Следовательно, химическое звено пресинаптического окончания работает как усилитель электрических сигналов.
Химическая передача осуществляет как возбуждающее, так и тормозное действие на постсинаптическую мембрану и зависит от медиатора и рецепторов постсинаптической мембраны. Из пресинаптического окончания выделяются следующие медиаторы:
1. Ацетилхолин - встречается в различных отделах ЦНС (кора большого мозга, ретикулярная формация ствола мозга, гипоталамус, спинной мозг). Известен в основном как возбуждающий медиатор (например, он является медиатором ά-мотонейронов спинного мозга). Тормозное влияние ацетилхолин оказывает в глубоких слоях коры большого мозга, стволе мозга и в хвостатом ядре.
2. Катехоламины(норадреналин, дофамин, серотонин, гистамин) в основном содержатся в нейронах ствола мозга, в меньших количествах в других отделах ЦНС. Например, амины обеспечивают возникновение процессов возбуждения и торможения в промежуточном мозге, черной субстанции, лимбической системе, полосатом теле.
1. Норадреналин. Норадренэргические нейроны сконцентрированы в основном в области голубого пятна (средний мозг). Норадреналин является тормозным медиатором клеток Пуркинье мозжечка и возбуждающим - в гипоталамусе, ядрах эпиталамуса. В ретикулярной формации ствола мозга и в гипоталамусе обнаружены ά - и β - адренорецепторы. Норадреналин регулирует настроение, эмоциональные реакции, обеспечивает поддержание бодрствования, участвует в механизмах формирования некоторых фаз сна, сновидений.
2. Дофамин. Дофаминэргические нейроны имеются в составе полосатого тела, в гипофизе. Дофамин участвует в формировании чувства удовольствия, регуляции эмоциональных реакций, поддержании бодрствования. Дофамин полосатого тела регулирует сложные мышечные движения.
3. Серотонин. Серотонин содержится главным образом в структурах, имеющих отношение к регуляции вегетативных функций. С помощью серотонина в нейронах ствола мозга передаются возбуждающие и тормозящие влияния, в коре - тормозящие. Серотонин ускоряет процессы обучения, формирование болевых ощущений, сенсорное восприятие, засыпание.
4. Гистаминв довольно высокой концентрации обнаруживается в гипофизе и срединном возвышении гипоталамуса. В остальных отделах содержание гистамина очень низко.
1. Аминокислоты. Кислые аминокислоты (глицин, γ - аминомасляная кислота - ГАМК) являются тормозными медиаторами в синапсах ЦНС. Глицин работает в спинном мозге, ГАМК - в коре больших полушарий, мозжечке, стволе мозга, спинном мозге. Нейтральные аминокислоты (ά - глутамат, ά - аспартат) передают возбуждение: глутаминовая кислота является основным возбуждающим медиатором. Рецепторы глутамата и аспарагиновой кислоты имеются на клетках спинного мозга, мозжечка, таламуса, гиппокампа, коры большого мозга.
2. Полипептиды. К ним относят энкефалины, эндорфины, ангиотензин, люлиберин, олигопептиды, субстанцию Р и пептид, вызывающий δ-сон.
1. Энкефалины и эндорфины - медиаторы нейронов, блокирующих болевуюимпульсацию. Они реализуют свое влияние посредством опиатных рецепторов, которые особенно плотно располагаются на клетках лимбической системы, черной субстанции, ядрах промежуточного мозга и голубого пятна спинного мозга. Энкефалины и эндорфины дают антиболевые реакции, повышение устойчивости к стрессу и сон.
2. Пептид, вызывающий δ-сонтакже дает антиболевые реакции, повышение устойчивости к стрессу и сон.
3. Ангиотензин участвует в передаче информации о потребности организма в воде, повышает артериальное давление, тормозит синтез катехоламинов, стимулирует секрецию гормонов, информирует ЦНС об осмотическом давлении крови.
4. Люлиберинучаствует в передаче информации о потребности организма в половой активности.
5. Олигопептиды - медиаторы настроения, полового поведения, передачи ноцицептивного возбуждения от периферии в ЦНС, формирования болевых ощущений.
6. Субстанция Р- является медиатором нейронов, передающих болевую информацию. Особенно много этого полипептида содержится в дорсальных корешках спинного мозга.
Кроме выше перечисленных медиаторов, существуют химические вещества, циркулирующие в крови, которые оказывают модулирующее действие на активность синапсов. К ним относятся простагландины и нейрогормоны. Простагландины влияют на секрецию медиатора и работу аденилатциклаз. Гипоталамические гормоны, регулирующие функцию гипофиза, также выполняют медиаторную функцию.
Эффект действия медиатора зависитв основном от свойств ионных каналов постсинаптической мембраны и вторых посредников. Например, ацетилхолин в коре большого мозга может вызвать и возбуждение и торможение, в синапсах сердца - торможение, в синапсах гладкой мускулатуры желудочно-кишечного тракта - возбуждение. Катехоламины стимулируют сердечную деятельность, но тормозят сокращения желудка и кишечника.
Механизмы долговременной памяти: синаптическая теория памяти
Перевод значимой информации из кратко- в долговременную память называется консолидацией энграммы. Этот процесс сопровождается образованием структурно-химических изменений в нервной системе. Существует две группы гипотез о способе фиксации информации в долговременной памяти: синаптическая теория и биохимические теории памяти.
Свое название синаптическая теория получила из-за того, что главное внимание в ней уделяется роли синапса в фиксации следа памяти. Она утверждает, что при прохождении импульса через определенную группу нейронов возникают стойкие изменения синаптической проводимости в пределах определенного нейронного ансамбля.
Г. Линч и М. Бодри (1984) предложили следующую гипотезу. Повторная импульсация в нейроне, связанная с процессом запоминания, предположительно, сопровождается увеличением концентрации кальция в постсинаптической мембране, что приводит к расщеплению одного из ее белков. В результате этого освобождаются замаскированные и ранее неактивные белковые рецепторы (глутаматрецепторы). За счет увеличения числа этих рецепторов возникает состояние повышенной проводимости синапса, которое может сохраняться до 5-6 суток.
Эти процессы тесно связаны с увеличением диаметра и усилением активности так называемого аксошипикового синапса - наиболее пластичного контакта между нейронами. Одновременно с этим образуются новые шипики на дендритах, а также увеличиваются число и величина синапсов. Таким образом, экспериментально показаны морфологические изменения, сопровождающие формирование следа памяти.
Медиаторам - химическим посредникам в синаптической передаче информации - придается большое значение в обеспечении механизмов долговременной памяти. Основные медиаторные системы головного мозга - холинэргическая и моноаминэргическая (включает норадреноэргическую, дофаминэргическую и серотонинэргическую) - принимают самое непосредственное участие в обучении и формировании энграмм памяти. Так, экспериментально установлено, что уменьшение количества норадреналина замедляет обучение, вызывает амнезию и нарушает извлечение следов из памяти.
Показано, что под влиянием обучения увеличивается количество холинорецепторов, т.е. рецепторов, расположенных на теле нейрона и отвечающих за обнаружение медиатора ацетилхолина. В процессе образования условного рефлекса повышается чувствительность соответствующих нейронов к ацетилхолину, что облегчает обучение, ускоряет запоминание и способствует более быстрому извлечению следа из памяти. В то же время вещества, препятствующие действию ацетилхолина, нарушают обучение и воспроизведение, вызывая амнезию (потерю памяти).
Важно подчеркнуть, что холинэргическая система испытывает на себе модулирующее влияние со стороны моноаминоэргической системы. Под действием этих влияний может изменяться активность холинэргических синапсов и запускаться цепь биохимических внутриклеточных процессов, приводящих к более эффективному образованию энграмм.
Биохимические теории памяти
В этой группе гипотез процессы памяти связывают со стойким изменением химических процессов в нервных клетках, в результате этих изменений облегчается проведение импульса через синапс.
Концепции биохимического кодирования индивидуального опыта в памяти, опираются на две группы фактов: 1) образование в мозге при обучении новых биохимических факторов (например, «пептидов памяти»); 2) возможность передачи приобретенной информации необученному мозгу с помощью этих факторов. Исследовались следующие биохимические процессы, связанные с памятью.
1. Усиление синтеза и изменение структуры нуклеиновых кислот (ДНК и РНК).
2. Усиление синтеза некоторых белков, например, содержащихся только в нервной ткани белков S-100 и 14-3-2.
3. Носителями долговременной памяти могут быть пептиды - вещества, состоящие из аминокислот, но более простые, чем белки.
4. Иммунохимические процессы - на основе связывания веществ-антигенов, вырабатываемых клеткой, с антигенами, вырабатываемыми глиальными (опорными и питающими) клетками.
Однако идея существования биохимических факторов, способных к сохранению и переносу информации, большинством исследователей воспринимается критически. В настоящее время считается, что гипотеза молекулярного кодирования индивидуального опыта не имеет прямых фактических доказательств. Несмотря на то, что установлена существенная роль нуклеиновых кислот и белков в механизмах научения и памяти, предполагается, что принимающие участие в формировании новой ассоциативной связи РНК и белки специфичны лишь по отношению к функциональному изменению участвующих в процессе синапсов и неспецифичны по отношению к самой информации.
Лекция 5. Физиологический подход к определению мышления, интеллекта и сознания
Понятие мышления и интеллекта, классификация видов мышления. Психофизиологический подход к измерению интеллекта. Структуры мозга, обеспечивающие мышление и интеллект. Факторы, определяющие развитие интеллекта. Физиологический подход к определению сознания. Мозговые центры и сознание. Физиологические теории сознания. Прожекторная теория.
1. Понятие мышления и интеллекта, классификация видов мышления
Мышление- опосредованное и обобщенное отражение действительности, в основе которого лежит произвольное оперирование образами и понятиями и которое дает знание о наиболее существенных свойствах, связях и отношениях объективного мира.
Мышление человека связано с процессами формирования понятий и решения задач с использованием этих понятий на основе определенных правил.
Иначе говоря, мышление - процесс познавательной деятельности, при котором субъект оперирует различными видами обобщений, включая образы, понятия и категории.
Уровень мышления, как способность к проникновению в суть возникающих проблем и выработки наиболее адекватных решений входит в понятие интеллекта.
Интеллект - общая познавательная способность, определяющая готовность к усвоению и использованию знаний и опыта, а также к разумному поведению в проблемных ситуациях. Критерий интеллекта - способность к отражению отношений и связей между предметами и ситуациями.
Существуют многочисленные классификации мышления. В зависимости от того, какие правила использует человек в процессе познания мира, выделяют мифологическое и логическое мышление. При логическом мышлении отдельные высказывания формируют цепочку для создания умозаключения, расчленяя объект анализом, направляя мысль от частного к общему (индукция) или от общего к частному (дедукция). Логическое мышление требует обязательной проверки результата мышления фактом и строится на основе фактов. В противоположность ему мифологическое мышление игнорирует факт, не расчленяет объект на признаки, нечувствительно к числу и особым образом относится к слову, поскольку для него сказать означает уже сделать (Лотман, Успенский, 1973).
Другой тип классификации определяет отношение мышления к практической деятельности и вычленяет соответственно теоретическое и практическое мышления. В свою очередь теоретическое делится на понятийное и образное в зависимости оттого, к чему обращается человек в процессе решения проблемы - к образу пли понятию. Практическое мышление также подразделяется на наглядно-образное и наглядно-действенное в зависимости от того, в какой мере непосредственное действие участвует в процессе решения задачи. Несмотря на существование такого деления, на практике эти виды мышления участвуют в обработке информации одновременно.
Связи между нейронами. Синапсы
Для передачи и переработки информации нейроны взаимодействуют друг с другом и с клетками исполнительных органов. Это взаимодействие осуществляется с помощью особых контактов — синапсов.
В синапсе различают пресипаптическую мембрану окончания нервного волокна, синаптическую щель (пространство между мембранами контактирующих клеток) и постсинаптическую мембрану, относящуюся к следующему в цепи нейрону или исполнительной клетке — мышечной, железистой (рис. 4.13). Число синапсов на одном нейроне может достигать нескольких тысяч (рис. 4.14). В зависимости от характера влияния на клетку различают возбуждающие и тормозные синапсы (рис. 4.15).
В настоящее время известны два способа передачи сигналов через синапс: химический и электрический. Нейроны мозга человека связаны между собой в основном химическими синапсами. Вещества, передающие сигналы через синап-
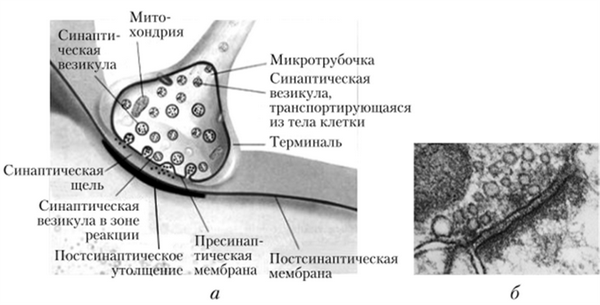
Рис. 4.13. Межнейрональные синапсы
а — схема; б — электронная микрофотография
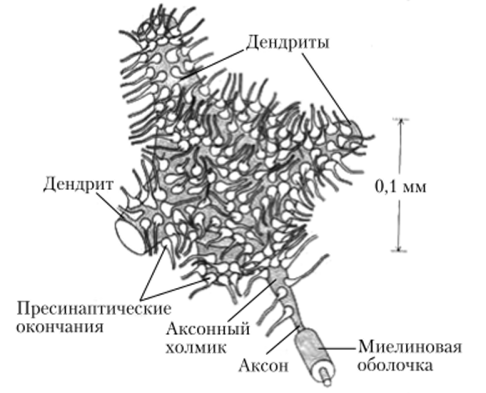
Рис. 4.14. Синапсы на теле и отростках могонейрона спинного мозга
тический контакт, называются медиаторами. К ним относятся ацетилхолин, адреналин, серотонин, гистамин, нейропептиды, некоторые аминокислоты (глутаминовая, аспаргино- вая, глицин, гамма-аминомасляная и др.).
Механизм передачи сигнала через синапс с участием медиатора показан на рис. 4.16. Медиатор находится в пузырьках пресинаптических окончаний. Когда нервные импульсы достигают окончания, пузырьки сливаются с пресинаптической
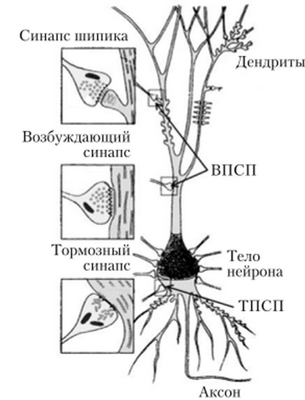
Рис. 4.15. Возбуждающий и тормозный синапсы нейронов головного мозга
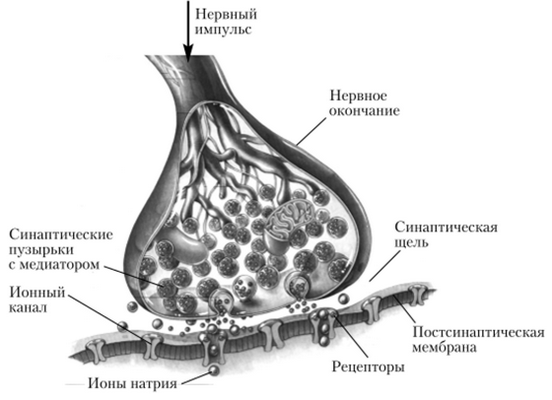
Рис. 4.16. Синаптическая передача
мембраной и медиатор выделяется в синаптическую щель. Он связывается с клеточными рецепторами поверхности постсинаптической мембраны следующей клетки, что ведет к изменению ее мембранного потенциала. Изменение заряда мембраны называется постсинаптическим потенциалом (ПСП). Он может быть возбуждающим (ВПСП) или тормозным (ТПСП).
Дендриты и сома нейрона принимают множество возбуждающих и тормозных синаптических сигналов, которые суммируются и создают суммарный постсинаптический потенциал мембраны. Этот потенциал на выходе из тела клетки, в мембране аксонного холмика, преобразуется в импульсные разряды, которые могут передаваться на большие расстояния. Таким образом, в синапсах химический сигнал преобразуется в электрический.
Долгое время существовала общепринятая точка зрения о том, что информация через синапс передается только в одном направлении: от пресинаптической мембраны к постсинаптической. Современные данные показали, что возможна передача и в противоположном направлении: от постсинаптической мембраны нейрона к пресинаптической мембране нервного окончания.
Были определены информационные молекулы, осуществляющие эту передачу: это полипептиды (фактор роста нерва), а также подвижные мелкие молекулы, среди которых — молекулы монооксида азота (N0). Показано также, что электрические процессы от постсинаптической мембраны могут пассивно распространяться по соседним дендритам на небольшие расстояния в обратную сторону, к пресинаптическому участку, оказывая влияние на синтез и выделение медиатора в синапсе. Передача информации в двух направлениях характерна также для электрических синапсов.
Передача информации через синапс в обоих направлениях важна для работы нейронных сетей. Она обеспечивает взаимодействие близко расположенных нейронов, участвует в образовании новых синапсов и их созревании.
Образование новых синапсов лежит в основе очень важного свойства нервной системы — пластичности, от которого зависят развитие мозга ребенка, процессы научения и памяти, восстановление нервных связей, утраченных при заболеваниях. Для этих процессов необходима обратная (ретроградная) передача сигнала — от постсинаптической к пре- синаптической мембране синапса. Только в таком случае образуется новый синапс.
Читайте также: