Механизм транспорта белков через аппарат Гольджи
Добавил пользователь Алексей Ф. Обновлено: 22.01.2026
В 1898 г. итальянский ученый К. Гольджи выявил в нервных клетках сетчатые образования, которые он назвал “внутренним сетчатым аппаратом” (рис. 174). Сетчатые структуры (аппарат Гольджи) встречаются во всех клетках любых эукариотных организмов. Обычно аппарат Гольджи располагается около ядра, вблизи клеточного центра (центриоли).
Тонкое строение аппарата Гольджи. Аппарат Гольджи состоит из мембранных структур, собранных вместе в небольшой зоне (рис. 176, 177). Отдельная зона скопления этих мембран называется диктиосомой (рис. 178). В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены в виде стопки плоские мембранные мешки, или цистерны, между которыми располагаются тонкие прослойки гиалоплазмы. Каждая отдельная цистерна имеет диаметр около 1 мкм и переменную толщину; в центре ее мембраны могут быть сближены (25 нм), а на периферии иметь расширения, ампулы, ширина которых непостоянна. Количество таких мешков в стопке обычно не превышает 5-10. У некоторых одноклеточных их число может достигать 20 штук. Кроме плотно расположенных плоских цистерн в зоне АГ наблюдается множество вакуолей. Мелкие вакуоли встречаются главным образом в периферических участках зоны АГ; иногда видно, как они отшнуровываются от ампулярных расширений на краях плоских цистерн. Принято различать в зоне диктиосомы проксимальный или формирующийся, цис-участок, и дистальный или зрелый, транс-участок (рис. 178). Между ними располагается средний или промежуточный участок АГ.
Во время деления клеток сетчатые формы АГ распадаются до диктиосом, которые пассивно и случайно распределяются по дочерним клеткам. При росте клеток общее количество диктиосом увеличивается.
В секретирующих клетках обычно АГ поляризован: его проксимальная часть обращена к цитоплазме и ядру, а дистальная - к поверхности клетки. В проксимальном участке к стопкам сближенных цистерн примыкает сетевидная или губкообразная система мембранных полостей. Считается, что эта система представляет собой зону перехода элементов ЭР в зону аппарата Гольджи (рис. 179).
В средней части диктиосомы периферия каждой цистерны также сопровождается массой мелких вакуолей около 50 нм в диаметре.
В дистальном или транс-участке диктиосом к последней мембранной плоской цистерне примыкает участок, состоящий из трубчатых элементов и массы мелких вакуолей, часто имеющих фибриллярную опушенность по поверхности со стороны цитоплазмы - это опушенные или окаймленные пузырьки такого же типа, как и окаймленные пузырьки при пиноцитозе. Это - так называемая транс-сеть аппарата Гольджи (TGN), где происходит разделение и сортировка секретируемых продуктов. Еще дистальнее располагается группа более крупных вакуолей - это уже продукт слияния мелких вакуолей и образования секреторных вакуолей.
С помощью мегавольтного электронного микроскопа было установлено, что в клетках отдельные диктиосомы могут быть связаны друг с другом системой вакуолей и цистерн и образовывать рыхлую трехмерную сеть, выявляемую в световом микроскопе. В случае диффузной формы АГ каждый отдельный его участок представлен диктиосомой. У клеток растений преобладает диффузный тип организации АГ, обычно в среднем на клетку приходится около 20 диктиосом. В клетках животных часто с зоной мембран аппарата Гольджи ассоциированы центриоли; между радиально отходящих от них пучков микротрубочек лежат группы стопок мембран и вакуолей, которые концентрически окружают клеточный центр. Эта связь, свидетельствует об участии микротрубочек в движении вакуолей.
Секреторная функция аппарата Гольджи. Основные функции АГ заключаются в накоплении продуктов, синтезированных в ЭР, обеспечение их химических перестроек, созревания.
В цистернах АГ происходит синтез полисахаридов, их взаимосвязь с белками. и образование мукопротеидов. Но главной функцией аппарата Гольджи является выведение готовых секретов за пределы клетки. Кроме того, АГ является источником клеточных лизосом.
Синтезированный на рибосомах экспортируемый белок отделяется и накапливается внутри цистерн ЭР, по которым он транспортируется к зоне мембран АГ. Здесь от гладких участков ЭР отщепляются мелкие вакуоли, содержащие синтезированный белок, которые поступают в зону вакуолей в проксимальной части диктиосомы. В этом месте вакуоли сливаются друг с другом и с плоскими цис-цистернами диктиосомы. Таким образом происходит перенесение белкового продукта уже внутри полостей цистерн АГ.
По мере модификации белки в цистернах аппарата Гольджи, с помощью мелких вакуолей переносятся от цистерн к цистерне в дистальную часть диктиосомы, пока не достигнут трубчатой мембранной сети в транс-участке диктиосомы. В этом участке происходит отщепление мелких пузырьков, содержащих уже зрелый продукт. Цитоплазматическая поверхность таких пузырьков бывает сходна с поверхностью окаймленных пузырьков, которые наблюдаются при рецепторном пиноцитозе. Отделившиеся мелкие пузырьки сливаются друг с другом, образуют секреторные вакуоли. После этого секреторные вакуоли начинают двигаться к поверхности клетки, плазматическая мембрана и мембраны вакуолей сливаются, и, таким образом, содержимое вакуолей оказывается за пределами клетки. Морфологически этот процесс экструзии (выбрасывания) напоминает пиноцитоз, только с обратной последовательностью стадий. Он носит название экзоцитоз.
В аппарате Гольджи происходит не только передвижение продуктов из одной полости в другую, но и происходит модификация белков, которая заканчивается адресацией продуктов, либо к лизосомам, плазматической мембране или к секреторным вакуолям.
Модификация белков в аппарате Гольджи. В цис-зону аппарата Гольджи синтезированные в ЭР белки попадают после первичного гликозилирования и редукции нескольких сахаридных остатков. После чего все белки получают одинаковые олигосахаридные цепи, состоящие из двух молекул N-ацетилглюкозамина, шести молекул маннозы (рис. 182). В цис-цистернах происходит вторичная модификация олигосахаридных цепей и их сортировка на два класса. В результате сортировки получается один класс фосфорилирующихся олигосахаридов (богатые маннозой) для гидролитических ферментов, предназначенных для лизосом, и другой класс олигосахаридов для белков, направляемых в секреторные гранулы или к плазматической мембране
Превращения олигосахаридов осуществляются с помощью ферментов - гликозилтрансфераз, входящих в состав мембран цистерн аппарата Гольджи. Так как каждая зона в диктиосомах имеет свой набор ферментов гликозилирования, то гликопротеиды как бы по эстафете переносятся из одного мембранного отсека (“этажа” в стопке цистерн диктиосомы) в другой и в каждом подвергаются специфическому воздействию ферментов. Так в цис-участке происходит фосфорилирование манноз в лизосомных ферментах и образуется особая маннозо-6-группировка, характерная для всех гидролитических ферментов, которые потом попадут в лизосомы.
В средней части диктиосом протекает вторичное гликозилирование секреторных белков: дополнительное удаление маннозы и присоединение N-ацетилглюкозамина. В транс-участке к олигосахаридной цепи присоединяются галактоза и сиаловые кислоты (рис. 183).
В ряде специализированных клеток в аппарате Гольджи происходит синтез собственно полисахаридов.
В аппарате Гольджи растительных клеток синтезируются полисахариды матрикса клеточной стенки (гемицеллюлозы, пектины). Диктиосомы растительных клеток участвуют в синтезе и выделении слизей и муцинов, в состав которых входят также полисахариды. Синтез же основного каркасного полисахарида растительных клеточных стенок, целлюлозы, происходит на поверхности плазматической мембраны.
В аппарате Гольджи клеток животных синтезируются длинные неразветвленные полисахаридные цепи глюкозаминогликанов. Глюкозаминогликаны ковалентно связываются с белками и образуют протеогликаны (мукопротеины). Такие полисахаридные цепи модифицируются в аппарате Гольджи и связываются с белками, которые в виде протеогликанов секретируются клетками. В аппарате Гольджи происходит также сульфатирование глюкозаминогликанов и некоторых белков.
Сортировка белков в аппарате Гольджи. В конечном итоге через аппарат Гольджи проходит три потока синтезированных клеткой нецитозольных белков: поток гидролитических ферментов для лизосом, поток выделяемых белков, которые накапливаются в секреторных вакуолях, и выделяются из клетки только по получении специальных сигналов, поток постоянно выделяемых секреторных белков. Следовательно, в клетке существует механизм пространственного разделения разных белков и их путей следования.
В цис- и средних зонах диктиосом все эти белки идут вместе без разделения, они только раздельно модифицируются в зависимости от их олигосахаридных маркеров.
Собственно разделение белков, их сортировка, происходит в транс-участке аппарата Гольджи. Принцип отбора лизосомных гидролаз происходит следующим образом (рис. 184).
Белки-предшественники лизосомных гидролаз имеют олигосахаридную, конкретнее маннозную группу. В цис-цистернах эти группировки фосфорилируются и вместе с другими белками переносятся в транс-участок. Мембраны транс-сети аппарата Гольджи содержат трансмембранный белок - рецептор (манноза-6-фосфатный рецептор или М-6-Ф-рецептор), который узнает фосфорилированные маннозные группировки олигосахаридной цепи лизосомных ферментов и связывается с ними. Следовательно М-6-Ф-рецепторы, являясь трансмембранными белками, связываясь с лизосомными гидролазами, отделяют их, отсортировывают, от других белков (например, секреторных, нелизосомных) и концентрируют их в окаймленных пузырьках. Оторвавшись от транс-сети эти пузырьки быстро теряют окаймление, сливаются с эндосомами, перенося таким образом свои лизосомные ферменты, связанные с мембранными рецепторами, в эту вакуоль. Внутри эндосом из-за активности протонного переносчика происходит закисление среды. Начиная с рН 6 лизосомные ферменты диссоциируют от М-6-Ф-рецепторов, активируются и начинают работать в полости эндолизосомы. Участки же мембран вместе с М-6-Ф-рецепторами возвращаются путем рециклизации мембранных пузырьков обратно в транс-сеть аппарата Гольджи.
Возможно, что часть белков, которая накапливается в секреторных вакуолях и выводится из клетки после поступления сигнала (например нервного или гормонального) проходит такую же процедуру отбора, сортировки на рецепторах транс-цистерн аппарата Гольджи. Секреторные белки также сначала попадают в мелкие вакуоли одетые клатрином, а затем сливаются друг с другом. В секреторных вакуолях белки накапливаются в виде плотных секреторных гранул, что приводит к повышению концентрации белка в этих вакуолях примерно в 200 раз, по сравнению с его концентрацией в аппарате Гольджи. По мере накопления белков в секреторных вакуолях и после получения клеткой соответствующего сигнала они путем экзоцитоза выбрасываются из клетки.
От аппарата Гольджи исходит и третий поток вакуолей, связанный с постоянной, конститутивной секрецией. Например, фибробласты выделяют большое количество гликопротеидов и муцинов, входящих в основное вещество соединительной ткани. Многие клетки постоянно выделяют белки, способствующие связыванию их с субстратами, постоянно идет поток мембранных пузырьков к поверхности клетки, несущие элементы гликокаликса и мембранных гликопротеидов. Этот поток выделяемых клеткой компонентов не подлежит сортировке в рецепторной транс-системе аппарата Гольджи. Первичные вакуоли этого потока также отщепляются от мембран и относятся по своей структуре к окаймленным вакуолям, содержащим клатрин (рис. 185).
Заканчивая рассмотрение строения и работы такой сложной мембранной органеллы, как аппарат Гольджи, необходимо подчеркнуть, что несмотря на кажущуюся морфологическую однородность его компонентов, вакуоли и цистерны, на самом деле, это не просто скопище пузырьков, а стройная, динамичная сложно организованная, поляризованная система.
В АГ происходит не только транспорт везикул от ЕР к плазматической мембране. Существует обратный перенос везикул. Так от вторичных лизосом отщепляются вакуоли и возвращаются вместе с рецепторными белками в транс-АГ зону, существует поток вакуолей от транс-зоны к цис-зоне АГ, а так же от цис-зоны к эндоплазматическому ретикулуму. В этих случаях вакуоли одеты белками COP I-комплекса. Считается, что таким путем возвращаются различные ферменты вторичного гликозилирования и рецепторные белки в составе мембран.
Особенности поведения транспортных везикул послужили основанием для гипотезы о существовании двух типов транспорта компонентов АГ (рис. 186).
По первому типу в АГ имеются стабильные мембранные компоненты, к которым от ЭР транспортными вакуолями эстафетно переносятся вещества. По другому типу АГ является производным ЭР: отщепившиеся от переходной зоны ЭР мембранные вакуоли сливаются друг с другом в новую цис-цистерну, которая затем продвигается через всю зону АГ и в конце распадается на транспортные везикулы. По этой модели ретроградные COP I везикулы возвращают постоянные белки АГ в более молодые цистерны.
Комплекс Гольджи
Аппарат Гольджи (комплекс Гольджи) — мембранная структура эукариотической клетки, органелла, в основном предназначенная для выведения веществ, синтезированных в эндоплазматическом ретикулуме. Комплекс Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1898 году.
Содержание
Строение
Комплекс Гольджи представляет собой стопку дискообразных мембранных мешочков (цистерн), несколько расширенных ближе к краям и связанную с ними систему пузырьков Гольджи. В растительных клетках обнаруживается ряд отдельных стопок (диктиосомы), в животных клетках часто содержится одна большая или несколько соединённых трубками стопок.
Транспорт веществ из эндоплазматической сети
Аппарат Гольджи асимметричен — цистерны, располагающиеся ближе к ядру клетки (цис-Гольджи) содержат наименее зрелые белки, к этим цистернам непрерывно присоединяются мембранные пузырьки — везикулы, отпочковывающиеся от гранулярного эндоплазматического ретикулума (ЭПР), на мембранах которого и происходит синтез белков рибосомами. Перемещение белков из эндоплазматической сети (ЭПС) в аппарат Гольджи происходит неизбирательно, однако не полностью или неправильно свернутые белки остаются при этом в ЭПС. Возвращение белков из аппарата Гольджи в ЭПС требует наличия специфической сигнальной последовательности (лизин-аспарагин-глутамин-лейцин) и происходит благодаря связыванию этих белков с мембранными рецепторами в цис-Гольджи.
Модификация белков в аппарате Гольджи
В цистернах Аппарата Гольджи созревают белки предназначенные для секреции, трансмембранные белки плазматической мембраны, белки лизосом и т. д. Созревающие белки последовательно перемещаются по цистернам органеллы, в которых происходит их модификации — гликозилирование и фосфорилирование. При О-гликозилировании к белкам присоединяются сложные сахара через атом кислорода. При фосфорилировании происходит присоединение к белкам остатка ортофосфорной кислоты.
Разные цистерны Аппарата Гольджи содержат разные резидентные каталитические ферменты и, следовательно, с созревающими белками в них последовательно происходят разные процессы. Понятно, что такой ступенчатый процесс должен как-то контролироваться. Действительно, созревающие белки «маркируются» специальными полисахаридными остатками (преимущественно маннозными), по-видимому, играющими роль своеобразного «знака качества».
Не до конца понятно, каким образом созревающие белки перемещаются по цистернам Аппарата Гольджи, в то время как резидентные белки остаются в большей или меньшей степени ассоциированы с одной цистерной. Существуют две взаимонеисключающие гипотезы, объясняющие этот механизм. Согласно первой (1), транспорт белков осуществляется при помощи таких же механизмов везикулярного транспорта, как и путь транспорта из ЭПР, причём резидентные белки не включаются в отпочковывающуюся везикулу. Согласно второй (2), происходит непрерывное передвижение (созревание) самих цистерн, их сборка из пузырьков с одного конца и разборка с другого конца органеллы, а резидентные белки перемещаются ретроградно (в обратном направлении) при помощи везикулярного транспорта.
Транспорт белков из аппарата Гольджи
В конце концов от транс-Гольджи отпочковываются пузырьки, содержащие полностью зрелые белки. Главная функция аппарата Гольджи - сортировка проходящих через него белков. В аппарате Гольджи происходит формирование "трехнаправленного белкового потока":
- созревание и транспорт белков плазматической мембраны;
- созревание и транспорт секретов;
- созревание и транспорт ферментов лизосом.
С помощью везикулярного транспорта прошедшие через апарат Гольджи белки доставляются "по адресу" в зависимсоти от полученных ими в аппарате Гольджи "меток". Механизмы этого процесса также не до конца понятны. Известно, что транспорт белков из аппарата Гольджи требует участия специфических мембранных рецепторов, которые опознают "груз" и обеспечивают избирательную стыковку пузырька с той или иной органеллой.
Образование лизосом
Все гидролитические ферменты лизосом проходят через аппарат Гольджи, где они получают "метку" в виде специфического сахара - маннозо-6-фосфата (М6Ф)- в составе своего олигосахарида. Присоединение этой метки происходит при участии двух ферментов. Фермент N-ацетилглюкозаминфосфотрансфераза специфически опознает лизосомальные гидролазы по деталям их третичной структуры и присоединяет N-ацетилглюкозаминфосфат к шестому атому нескольких маннозных остатков олигосахарида гидролазы. Второй фермент - фосфогликозидаза - отщепляет N-ацетилглюкозамин, создавая М6Ф-метку. Затем эта метка опознается белком-рецептором М6Ф, с его помощью гидролазы упаковываются в везикулы и доставляются в лизосомы. Там, в кислой среде, фосфат отщепляется от зрелой гидролазы. При нарушении работы N-ацетилглюкозаминфосфотрансферазы из-за мутаций или пригенетических дефектах рецептора М6Ф все ферменты лизосом "по умолчанию" доставляются к наружной мембране и секретируются во внеклеточную среду. Выяснилось, что в норме некоторое количество рецепторов М6Ф также попадают на нарушную мембрану. Они возвращают случайно попавшие во внешнюю среду ферменты лизосом внутрь клетки в процессе эндоцитоза.
Транспорт белков на наружную мембрану
Как правило, ещё в ходе синтеза белки наружной мембраны встраиваются своими гидрофобными участками в мембрану эндоплазматической сети. Затем в составе мембраны везикул они доставляются в аппарат Гольджи, а оттуда - к поверхности клетки. При слиянии везикулы с плазмалеммой такие белки остаются в ее составе, а не выделяются во внешнюю среду, как те белки, что находились в полости везикулы.
Секреция
Практически все секретируемые клеткой вещества (как белковой, так и небелковой природы) проходят через аппарат Гольджи и там упаковываются в секреторные пузырьки. Так, у растений при участии диктиосом секретируется материал клеточной стенки.
Пластинчатый комплекс
Аппарат Гольджи
Аппара́т (ко́мплекс) Го́льджи — мембранная структура эукариотической клетки, органелла, в основном предназначенная для выведения веществ, синтезированных в эндоплазматическом ретикулуме. Аппарат Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1897 году. [1]
Комплекс Гольджи представляет собой стопку дискообразных мембранных мешочков (цистерн), несколько расширенных ближе к краям, и связанную с ними систему пузырьков Гольджи. В растительных клетках обнаруживается ряд отдельных стопок (диктиосомы), в животных клетках часто содержится одна большая или несколько соединённых трубками стопок.
В Комплексе Гольджи выделяют 3 отдела цистерн, окружённых мембранными пузырьками:
- Цис-отдел (ближний к ядру);
- Медиальный отдел;
- Транс-отдел (самый отдалённый от ядра).
Эти отделы различаются между собой набором ферментов. В цис-отделе первую цистерну называют "цистерной спасения", так как с её помощью рецепторы, поступающие из промежуточной эндоплазматической сети, возвращаются обратно. Фермент цис-отдела: фосфогликозидаза (присоединяет фосфат к углеводу - манназе). В медиальном отделе находится 2 фермента: манназидаза (отщепляет манназу) и N-ацетилглюкозаминтрансфераза (присоединяет определенные углеводы - гликозамины). В транс-отделе ферменты: пептидаза (осуществляет протеолиз) и трансфераза (осуществляет переброс химических групп).
Функции
- Сегрегация белков на 3 потока:
- лизосомальный - гликозилированные белки (с маннозой) поступают в цис-отдел комплекса Гольджи, некоторые из них фосфорилируются, образуется маркёр лизосомальных ферментов - манноза-6-фосфат. В дальнейшем эти фосфорилированные белки не будут подвергаться модификации, а попадут в лизосомы.
- конститутивный экзоцитоз (конститутивная секреция). В этот поток включаются белки и липиды, которые становятся компонентами поверхностного аппарата клетки, в том числе гликокаликса, или же они могут входить в состав внеклеточного матрикса.
- Индуцируемая секреция - сюда попадают белки, которые функционируют за пределами клетки, поверхностного аппарата клетки, во внутренней среде организма. Характерен для секреторных клеток.
- Формирование слизистых секретов - гликозамингликанов (мукополисахаридов)
- Формирование углеводных компонентов гликокаликса - в основном, гликолипидов.
- Сульфатирование углеводных и белковых компонентов гликопротеидов и гликолипидов
- Частичный протеолиз белков - иногда за счет этого неактивный белок переходит в активный (проинсулин превращается в инсулин).
В цистернах аппарата Гольджи созревают белки предназначенные для секреции, трансмембранные белки плазматической мембраны, белки лизосом и т. д. Созревающие белки последовательно перемещаются по цистернам в органеллы, в которых происходят их модификации — гликозилирование и фосфорилирование. При О-гликозилировании к белкам присоединяются сложные сахара через атом кислорода. При фосфорилировании происходит присоединение к белкам остатка ортофосфорной кислоты.
Схема, показывающая цитоплазму, вместе с ее компонентами (или органеллами), в типичной животной клетке. Органеллы: 1 — ядрышко; 2 — ядро; 3 — рибосома (маленькие точки); 4 — везикула; 5 — шероховатый эндоплазматический ретикулум (ER); 6 — аппарат Гольджи; 7 — цитоскелет; 8 — гладкий эндоплазматический ретикулум; 9 — митохондрия; 10 — вакуоль; 11 — цитоплазма; 12 — лизосома; 13 — центриоль и центросома.
Не до конца понятно, каким образом созревающие белки перемещаются по цистернам аппарата Гольджи, в то время как резидентные белки остаются в большей или меньшей степени ассоциированы с одной цистерной. Существуют две взаимонеисключающие гипотезы, объясняющие этот механизм:
- согласно первой, транспорт белков осуществляется при помощи таких же механизмов везикулярного транспорта, как и путь транспорта из ЭПР, причём резидентные белки не включаются в отпочковывающуюся везикулу;
- согласно второй, происходит непрерывное передвижение (созревание) самих цистерн, их сборка из пузырьков с одного конца и разборка с другого конца органеллы, а резидентные белки перемещаются ретроградно (в обратном направлении) при помощи везикулярного транспорта.
В конце концов от транс-Гольджи отпочковываются пузырьки, содержащие полностью зрелые белки. Главная функция аппарата Гольджи — сортировка проходящих через него белков. В аппарате Гольджи происходит формирование «трехнаправленного белкового потока»:
- созревание и транспорт белков плазматической мембраны;
- созревание и транспорт секретов;
- созревание и транспорт ферментов лизосом.
С помощью везикулярного транспорта прошедшие через аппарат Гольджи белки доставляются «по адресу» в зависимости от полученных ими в аппарате Гольджи «меток». Механизмы этого процесса также не до конца понятны. Известно, что транспорт белков из аппарата Гольджи требует участия специфических мембранных рецепторов, которые опознают «груз» и обеспечивают избирательную стыковку пузырька с той или иной органеллой.
Все гидролитические ферменты лизосом проходят через аппарат Гольджи, где они получают «метку» в виде специфического сахара — маннозо-6-фосфата (М6Ф)- в составе своего олигосахарида. Присоединение этой метки происходит при участии двух ферментов. Фермент N-ацетилглюкозаминфосфотрансфераза специфически опознает лизосомальные гидролазы по деталям их третичной структуры и присоединяет N-ацетилглюкозаминфосфат к шестому атому нескольких маннозных остатков олигосахарида гидролазы. Второй фермент — фосфогликозидаза — отщепляет N-ацетилглюкозамин, создавая М6Ф-метку. Затем эта метка опознается белком-рецептором М6Ф, с его помощью гидролазы упаковываются в везикулы и доставляются в лизосомы. Там, в кислой среде, фосфат отщепляется от зрелой гидролазы. При нарушении работы N-ацетилглюкозаминфосфотрансферазы из-за мутаций или при генетических дефектах рецептора М6Ф все ферменты лизосом «по умолчанию» доставляются к наружной мембране и секретируются во внеклеточную среду. Выяснилось, что в норме некоторое количество рецепторов М6Ф также попадают на наружную мембрану. Они возвращают случайно попавшие во внешнюю среду ферменты лизосом внутрь клетки в процессе эндоцитоза.
Как правило, ещё в ходе синтеза белки наружной мембраны встраиваются своими гидрофобными участками в мембрану эндоплазматической сети. Затем в составе мембраны везикул они доставляются в аппарат Гольджи, а оттуда — к поверхности клетки. При слиянии везикулы с плазмалеммой такие белки остаются в ее составе, а не выделяются во внешнюю среду, как те белки, что находились в полости везикулы.
Комплекс (аппарат) Гольджи
Комплекс Гольджи — одна из органелл мышечного волокна общего назначения. Функциями комплекса Гольджи являются: перенос и преобразование белков, сборка мембран, транспорт различных веществ к клеточной мембране, формирование лизосом.
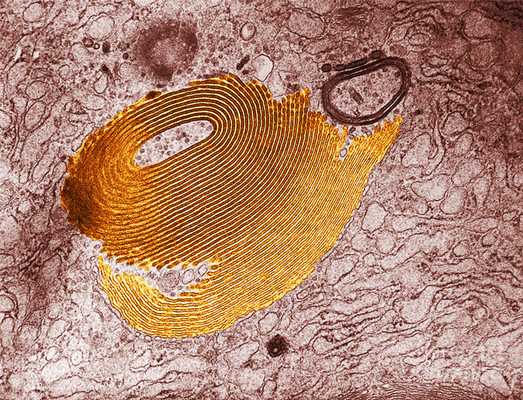
Как и в любой другой клетке, в мышечном волокне имеется органелла общего назначения, известная под названием комплекса или аппарата Гольджи. Эту органеллу обнаружил в 1889 году итальянский ученый Камилло Гольджи. В саркоплазме мышечного волокна таких органелл насчитывается тысячи.
Строение комплекса Гольджи
Комплекс Гольджи представляет собой стопку дискообразных мембранных цистерн (рис.1) и связанную с ними систему пузырьков (везикул). Между цистернами имеются связывающие их трубчатые структуры.
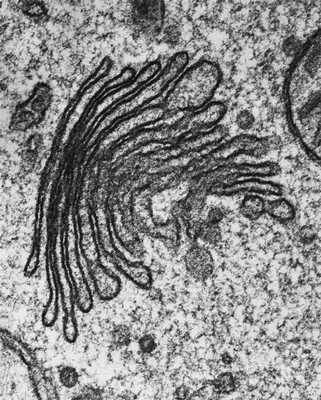
Рис.1. Комплекс Гольджи
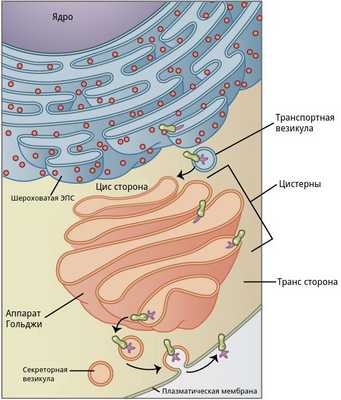
Рис.2. Структура и функции комплекса Гольджи
В комплексе Гольджи различают три отдела: цис-сеть Гольджи, некомпактные зоны и транс-сеть Гольджи.
Цис-сеть Гольджи расположена перед первой цистерной, ближе к ядру мышечного волокна. В транс-сеть Гольджи, переходит последняя цистерна комплекса. Она расположена на более удаленном расстоянии от ядра мышечного волокна. Некомпактные зоны расположены между соседними стопками аппарата Гольджи.
Функции комплекса Гольджи:
- Перенос и преобразование белков;
- Сборка мембран;
- Транспорт веществ к клеточной мембране;
- Формирование лизосом
Перенос и преобразование белков
Необходимые мышечному волокну белки синтезируются на рибосомах. Затем они перемещаются в шероховатую эндоплазматическую сеть. Эти белки называют «незрелыми», потому что они еще не приняли свою пространственную структуру (то есть не свернулись определенным образом). Из шероховатой эндоплазматической сети белки перемещаются в виде мембранных пузырьков в цис-сеть комплекса Гольджи. Известно, что в цистернах, располагающихся ближе к ядру мышечного волокна, содержатся наименее зрелые белки. В цистернах аппарата Гольджи белки «созревают», то есть приобретают свою трехмерную структуру. Биохимики называют этот этап процессингом белка. Каким образом созревающие белки перемещаются по цистернам комплекса Гольджи до сих пор непонятно.
В конце концов от цистерн, расположенных достаточно далеко от ядер отпочковываются пузырьки, содержащие полностью «зрелые» белки. С помощью мембранных пузырьков эти белки доставляются «по адресу» в зависимости от полученных ими в аппарате Гольджи «меток». Образно говоря, в мышечном волокне комплекс Гольджи играет роль почты, в которой сортируются и оправляются по адресам различные «посылки» в виде белков, заключенных в пузырьки.
Сборка мембран
В комплексе Гольджи происходит также сборка мембран. Вещества, из которых состоят мембраны (белки, липиды) поступают в комплекс Гольджи из эндоплазматической сети. Затем в цистернах комплекса Гольджи собираются участки мембран, из которых изготавливаются мембранные пузырьки. Они перемещаются в саркоплазме мышечного волокна в те места, где нужно достроить мембрану.
Транспорт веществ к клеточной мембране
Часть веществ, синтезированных в мышечном волокне, выводится наружу. Эти вещества накапливаются в комплексе Гольджи, упаковываются в мембранные пузырьки и транспортируются к сарколемме и выводятся за её пределы.
Образование лизосом
В комплексе Гольджи также образуются лизосомы - органеллы мышечного волокна, играющие существенную роль в катаболизме белков.
Читайте также:
- Вторичные ампутации. Показания к вторичным ампутациям. Виды рассечения тканей при ампутации. Гильотинная ампутация.
- Возраст пациента в хирургии. Анамнез заболеваний
- Посоветовали имбирь вместо виагры. Укрепляем мужскую потенцию народными средствами
- Гипоплазия и аплазия тимуса. Синдром Ди Джорджи
- Эндотелий сосудов. Роль ( значение ) эндотелия в регуляции просвета сосудов.