Мышечное сокращение. Характеристика и механизм мышечного сокращения
Добавил пользователь Владимир З. Обновлено: 28.01.2026
1. Генерация потенциала действия. Передача возбуждения на мышечное волокно происходит с помощью ацетилхолина. Взаимодействие ацетилхолина (АХ) с холинорецепторами приводит к их активации и появлению потенциала действия, что является первым этапом мышечного сокращения.
2. Распространение потенциала действия. Потенциал действия распространяется внутрь мышечного волокна по поперечной системе трубочек, которая является связывающим звеном между поверхностной мембраной и сократительным аппаратом мышечного волокна.
3. Электрическая стимуляция места контакта приводит к активации фермента и образованию инозилтрифосфата, который активирует кальциевые каналы мембран, что приводит к выходу ионов Ca и повышению их внутриклеточной концентрации.
Хемомеханический этап мышечного сокращения.
Теория хемомеханического этапа мышечного сокращения была разработана О. Хаксли в 1954 г. и дополнена в 1963 г. М. Девисом. Основные положения этой теории:
1) ионы Ca запускают механизм мышечного сокращения;
2) за счет ионов Ca происходит скольжение тонких актиновых нитей по отношению к миозиновым.
В покое, когда ионов Ca мало, скольжения не происходит, потому что этому препятствуют молекулы тропонина и отрицательно заряды АТФ, АТФ-азы и АДФ. Повышенная концентрация ионов Ca происходит за счет поступления его из межфибриллярного пространства. При этом происходит ряд реакций с участием ионов Ca:
1)Ca 2+ реагирует с трипонином;2)Ca 2+ активирует АТФ-азу;3)Ca 2+ снимает заряды с АДФ, АТФ, АТФ-азы.
Взаимодействие ионов Ca с тропонином приводит к изменению расположения последнего на актиновой нити, открываются активные центры тонкой протофибриллы. За счет них формируются поперечные мостики между актином и миозином, которые перемещают актиновую нить в промежутки между миозиновой нитью. При перемещении актиновой нити относительно миозиновой происходит сокращение мышечной ткани.
Итак, главную роль в механизме мышечного сокращения играют белок тропонин, который закрывает активные центры тонкой протофибриллы и ионы Ca.
14.Режимы мышечного сокращения. Одиночное мышечное сокращение(омс) и его периоды. Суммация,тетанус,их мех-мы. Особенности мышечного сокращения у детей.
Исследование сократительной способности мышцы производят в различных режимах.Выделяют следующие режимы: изотонический режим - мышца укорачивается, но при этом еенапряжение не изменяется. Изометрический режим - меняется напряжение, но длина мышцы остается без изменений. Смешанный режим - меняется длина мышцы и ее напряжение. Виды мышеч сокращений:одичное мышечное сокращение, суммированное мыщечное сокращение, тетаническое мыш сок-ние, тоническ мыш сок-ние. Одиночное мыш сок-ние - сок-ние мышцы при раздр-е одиночным пороговым стимулом. Периоды:1)латентный(скрытй период от момента нанесения раздражения до появления мышеч сок-ния 0,01с); 2)укорочение(0,04с); 3)расслабление(0,05с). Величина одиночного сокр-я равна0,1с. Суммация - увелич амплитуды мыш сок-ния при действии на мышцу 2последоват стимулов, если интервал времени между ними меньше, чем длительность одиноч мыш сок-ния, но больше, чем латентный период. Виды суммации:полная(когда 2радражитель попадает в фазу укорочения мышцы) и неполная(когда 2раздражитель попадает в фазурасслабления). Тетанус - множественное сокращение м-ц под действием высокочастотного раздражителя. Тетанус может быть гладким(возникает при действии раздраж относит высок частоты и явл результатом полн мышечн суммации) и зубчатым (возникает при действии раздраж относит небольш частоты и явл результатом неполн мыш суммации). Зубчатый бывает мелко- и крупнозубчатый. Чем выше частота стимула, тем выше амплитуда тетануса.Особенностью мышц плода и новорожден явл медленность ОМС - как фазы укорочения, так и фазы расслабления. Также отсутствуют различия скорости будущих быстрых и медленных мышц, хотя сами мышцы отлич по цвету(красн и бел) и по гистохимич признакам. У новорожден наблюдается ускорение как бастр, так и медлен волокон, но у медлен происходит вторич замедление сокращения. Ускорение обусловлено интенсив активацией мышеч белков и увелич числа саркомеров. С возрасто увелич сила сок-ний мышц.
ЗАНЯТИЕ №5: Физиология мышц.
1. Виды мышц в организме, морфо-функциональная характеристика скелетных мышц. Физиологические свойства мышечной ткани.
2. Механизм мышечного сокращения.
3. Одиночное мышечное сокращение скелетной мышцы, условия получения, фазы. Временные соотношения возбуждения и сокращения в мышцах разных видов.
4. Основные параметры мышечного сокращения. Зависимости «длина-сила» и «сила-время».
5. Тетаническое сокращение. Условия получения различных видов тетануса. Зависимость вида сокращения от лабильности ткани и частотных характеристик действующего раздражителя.
6. Регуляция мышечного сокращения. Понятие «двигательная единица».
7. Особенности строения и физиологических свойств гладкой мышцы. Автоматия, определение понятия, значение.
ДОМАШНЕЕ ЗАДАНИЕ
1. Дайте классификацию мышц, укажите их физиологические свойства.
В зависимости от особенностей строения мышцы человека делят на 3 типа или группы:
Первая группа мышц — скелетные, или поперечнополосатые мышцы. Скелетных мышц у каждого из нас более 600. Мышцы этого типа способны произвольно, по желанию человека, сокращаться и вместе со скелетом образуют опорно-двигательную систему. Общая масса этих мышц составляет около 40 % веса тела, а у людей, активно развивающих свои мышцы, может быть ещё больше. С помощью специальных упражнений размер мышечных клеток можно увеличивать до тех пор, пока они не вырастут в массе и объёме и не станут рельефными. Сокращаясь, мышца укорачивается, утолщается и движется относительно соседних мышц. Укорочение мышцы сопровождается сближением её концов и костей, к которым она прикрепляется. В каждом движении участвуют мышцы как совершающие его, так и противодействующие ему (агонисты и антагонисты соответственно), что придаёт движению точность и плавность.
Второй тип мышц, который входит в состав клеток внутренних органов, кровеносных сосудов и кожи, — гладкая мышечная ткань, состоящая из характерных мышечных клеток (миоцитов). Короткие веретеновидные клетки гладких мышц образуют пластины. Сокращаются они медленно и ритмично, подчиняясь сигналам вегетативной нервной системы. Медленные и длительные их сокращения происходят непроизвольно, то есть независимо от желания человека.
Гладкие мышцы, или мышцы непроизвольных движений, находятся главным образом в стенках полых внутренних органов, например пищевода или мочевого пузыря. Они играют важную роль в процессах, не зависящих от нашего сознания, например в перемещении пищи по пищеварительному тракту.
Отдельную (третью) группу мышц составляет сердечная поперечнополосатая (исчерченная) мышечная ткань (миокард). Она состоит из кардиомиоцитов. Сокращения сердечной мышцы не подконтрольны сознанию человека, она иннервируется вегетативной нервной системой.
2. Перечислите виды мышечных сокращений, характерные для разных видов мышечной ткани.
Существует два вида мышечных сокращений - одиночное и тетаническое.
Одиночное мышечное сокращение является единственным видом сокращений для сердечной мышцы, а в скелетной мускулатуре оно носит искусственную этиологию и возникает в ответ на одиночный электрический сигнал и возникновение потенциала действия (ПД). Такое сокращение, длящееся» 100 мс, имеет форму волны (см. рис.) и включает три фазы: 1 - латентный период (от 2-3 до 10 мс), длящийся от момента нанесения раздражения до начала сокращения, 2 - фаза укорочения или сокращения (40-50 мс) и 3 - фаза расслабления (около 50мс).
В естественных условиях импульсы поступают не одиночно, а сериями не менее 15-50 имп/с, на что мышца отвечает возникновением тетанического сокращения (тетануса). В его основе лежит явление суммации нескольких одиночных сокращений. В зависимости от частоты импульсов различают зубчатый и гладкий тетанус.
3. Схематически изобразить структуры, участвующие в механизме мышечного сокращения скелетной мышцы, указать его основные этапы на схеме.
Механизм мышечного сокращения.
1 - актиновый филламент,
2 - центр связывания,
3 - миозиновый филламент,
4 - головка миозина,
5 - Z -диск саркомера.
4. Укажите условия получения одиночного мышечного сокращения (ОМС).
При раздражении мышцы одиночным импульсом тока возникает одиночное мышечное сокращение.
Амплитуда одиночного сокращения мышцы зависит от количества сократившихся в этот момент миофибрилл. Возбудимость отдельных групп волокон различна, поэтому пороговая сила тока вызывает сокращение лишь наиболее возбудимых мышечных волокон. Амплитуда такого сокращения минимальна. При увеличении силы раздражающего тока в процесс возбуждения вовлекаются и менее возбудимые группы мышечных волокон; амплитуда сокращений суммируется и растет до тех пор, пока в мышце не останется волокон, не охваченных процессом возбуждения. В этом случае регистрируется максимальная амплитуда сокращения, которая не увеличивается, несмотря на дальнейшее нарастание силы раздражающего тока.
5. Дайте определение и укажите условия получения различных видов тетануса:
Если раздражающие импульсы сближены и каждый из них приходится на тот момент, когда мышца только начала расслабляться, но не успела еще полностью расслабиться Зубчатый тетанус возникает при частоте нервных импульсов до 30 в 1с. Если мышца получает 10-20 нервных импульсов в 1с, то она находится в состоянии мышечного тонуса, т.е. умеренной степени напряжения
Если раздражающие импульсы сближены настолько, что каждый последующий приходится на время, когда мышца еще не успела перейти к расслаблению от предыдущего раздражения, то есть происходит на высоте ее сокращения, то возникает длительное непрерывное сокращение, получившее название гладкого тетануса.
Гладкий тетанус - нормальное рабочее состояние скелетных мышц обусловливается поступлением из ЦНС нервных импульсов с частотой 40-50 в 1с.
6. Перечислите свойства гладких мышц и особенности механизма сокращения.
Гладкие мышцы образуют стенки (мышечный слой) внутренних органов и кровеносных сосудов. В миофибриллах гладких мышц нет поперечной исчерченности. Это обусловлено хаотичным расположением сократительных белков. Волокна гладких мышц относительно короче.
Гладкие мышцы менее возбудимы, чем поперечнополосатые. Возбуждение по ним распространяется с небольшой скоростью - 2-15 см/с. Возбуждение в гладких мышцах может передаваться с одного волокна на другое, в отличие от нервных волокон и волокон поперечнополосатых мышц.
Сокращение гладкой мускулатуры происходит более медленно и длительно.
Рефрактерный период в гладких мышцах более продолжителен, чем в скелетных.
Важным свойством гладкой мышцы является ее большая пластичность, т.е. способность сохранять приданную растяжением длину без изменения напряжения. Данное свойство имеет существенное значение, так как некоторые органы брюшной полости (матка, мочевой пузырь, желчный пузырь) иногда значительно растягиваются.
Характерной особенностью гладких мышц является их способность к автоматической деятельности, которая обеспечивается нервными элементами, заложенными в стенках гладкомышечных органов.
Адекватным раздражителем для гладких мышц является их быстрое и сильное растяжение, что имеет большое значение для функционирования многих гладкомышечных органов (мочеточник, кишечник и другие полые органы)
Особенностью гладких мышц является также их высокая чувствительность к некоторым биологически активным веществам (ацетилхолин, адреналин, норадреналин, серотонин и др.).
Гладкие мышцы иннервируются симпатическими и парасимпатическими вегетативными нервами, которые, как правило, оказывают противоположное влияние на их функциональное состояние.
Факторы, определяющие силу и скорость сокращения мышц
Четвертая лекция по дисциплине «Биомеханика мышц» посвящена факторам, определяющим силу и скорость сокращения мышц. Выделены три группы факторов: анатомические (площадь поперечного сечения мышцы, ход мышечных волокон, состав мышц); физиологические (частота импульсации ДЕ, количество рекрутированных ДЕ, синхронизация активности ДЕ); биомеханические (длина мышцы, режим работы мышцы и величина внешнего отягощения).
Лекция 4
4.1. Основные понятия
При изучении дисциплин «Физика» и «Биомеханика» вы познакомились с понятием силы как количественной меры взаимодействия тел.
Под силой мышцы (или силой мышечной тяги) будем понимать силу, регистрируемую на ее конце (то есть количественную меру взаимодействия мышцы и регистрирующего прибора).
Условно можно выделить три вида факторов, определяющих силу и скорость сокращения мышц:
4.2. Анатомические факторы, определяющие силу и скорость сокращения мышц
4.2.1. Сила и скорость сократительного компонента мышцы
К анатомическим факторам, определяющим силу сократительного компонента мышцы и скорость его сокращения, относятся:
Площадь поперечного сечения мышечного волокна (Sмв)
Сила сократительного компонента мышцы во многом зависит от площади его поперечного сечения. Экспериментально установлено, что чем больше площадь поперечного сечения мышечного волокна, (то есть, чем оно толще), тем большую силу оно способно развить. У не занимающихся физической культурой и спортом диаметр мышечных волокон равен 20 - 80 мкм, следовательно, площадь поперечного сечения мышечных волокон варьирует от 300 мкм 2 до 5000 мкм 2 . У бодибилдеров площадь поперечного сечения мышечных волокон значительно больше. Она может достигать 20000 мкм 2. На площадь поперечного сечения мышц влияет ряд факторов, таких как: расположение мышцы (верхние или нижние конечности), пол, возраст, особенности конституции человека, а также его уровень тренированности или степень гиподинамии.
Количество мышечных волокон (nмв)
Количество мышечных волокон - второй анатомический фактор, определяющий силу сократительного компонента мышцы. Чем больше волокон содержит мышца, тем большую силу она способна проявить при прочих равных условиях. Число волокон в мышцах различно (табл. 4.1). Так, прямая мышца бедра содержит несколько десятков тысяч волокон, а икроножная - более миллиона!
Таблица 4.1 Количество волокон в скелетных мышцах человека
МакКомас А. Дж., 2001
В анатомии, физиологии и биомеханике хорошо известен принцип Вебера: «Сила мышц, при прочих равных условиях, пропорциональна ее поперечному сечению» (Ухтомский А.А, 1927.- С. 72). Из мышц, обслуживающих тазобедренный сустав, самый большой физиологический поперечник у односуставных разгибателей бедра (большой ягодичной м.): 30,3±9,4 см 2 . Из мышц, обслуживающих движения в коленном суставе самый большой физиологический поперечник - у четырехглавой мышцы бедра: 56,0±11,0 см 2 . Из мышц, обслуживающих голеностопный сустав - у камбаловидной м.: 23,7±10,6 см 2 . Эти мышцы носят название антигравитационных.
Преобладание физиологических поперечников этих мышц над их антагонистами связано с необходимостью постоянно противодействовать силе тяжести (гравитации). Именно с этим связан тот факт, что площадь поперечного сечения мышц-разгибателей нижних конечностей (антигравитационных мышц) относится к площади поперечного сечения их антагонистов как 2:1, в то время как для мышц верхних конечностей это отношение составляет 1:1.
Следует запомнить, что тренировка на развитие силы мышцы приводит к увеличению площади ее поперечного сечения. Возрастание этого показателя происходит за счет увеличения площади поперечного сечения мышечных волокон (гипертрофии мышечных волокон). Увеличения количества мышечных волокон (гиперплазии) в мышце не наблюдается.
Ход мышечных волокон
Ход мышечных волокон определяет как силу сократительного компонента мышцы, так и скорость его сокращения.
В организме человека, кроме веретенообразных мышц с прямым ходом волокон, имеются перистые мышцы. В перистых мышцах не вся сила, генерируемая мышечным волокном, передается сухожилию. Это связано с тем, что мышечные волокна расположены под углом к длиннику мышцы. Этот угол называется углом перистости. Чем больше угол перистости, тем больше проигрыш в силе, передаваемой мышечным волокном сухожилию. Теоретически, если угол перистости равен 90 град, мышечное волокно не оказывает тянущего усилия на сухожилие. Однако в реальных условиях угол перистости варьирует в пределах от 10 до 30 град. Расчеты показывают, что проигрыш в силе из-за расположения мышечного волокна под углом к сухожилию небольшой. Однако благодаря перистой архитектуре в том же объеме, какой занимает веретенообразная мышца, может быть «упаковано» значительно больше мышечных волокон, поэтому значительно возрастает физиологический поперечник а, следовательно, и сила, развиваемая перистой мышцей. Благодаря перистой архитектуре камбаловидная мышца будет выигрывать в силе у мышцы с параллельным ходом мышечных волокон более чем в 10 раз. Следует отметить, что в гипертрофированных мышцах углы перистости больше.
Именно поэтому большинство антигравитационных мышц имеет перистое строение. К ним относятся: четырехглавая мышца бедра, трехглавая мышца голени. Следует отметить, что, с одной стороны, перистая мышца превышает показатели мышцы с прямым ходом мышечных волокон по силе сокращения, с другой - во столько же раз проигрывает в скорости сокращения.
Длина мышечных волокон
Длина мышечных волокон определяет как силу, так и скорость сокращения мышцы. Стало аксиомой утверждение, что «короткие мышцы сильные, длинные - быстрые». Принцип Бернулли гласит, что степень сокращения мышцы при прочих равных условиях пропорциональна длине ее волокон. Поэтому, чем длиннее мышца, тем в большей степени она способна укоротиться за единицу времени и, следовательно, тем больше ее скорость сокращения.
Состав мышц
От того, из какого типа волокон состоит мышца, зависит как сила сократительного компонента, так и скорость его сокращения.
- медленные неутомляемые (I тип);
- быстрые неутомляемые или промежуточные (IIА тип);
- быстрые утомляемые (IIВ тип).
Мышечным волокнам соответствуют различные двигательные единицы (ДЕ).
Состав мышечных волокон разных мышц сильно отличается и в одной и той же мышце имеет огромные индивидуальные различия, зависящие от врожденных типологических особенностей человека. Более подробная информация приведена в первой главе.
4.3. Физиологические факторы, определяющие силу и скорость сокращения мышц
4.3.1. Физиологические механизмы регуляции силы и скорости сокращения мышцы
К физиологическим механизмам регуляции силы и скорости сокращения мышцы относятся:
- частота (паттерн) разрядов двигательной единицы (ДЕ);
- число активных ДЕ;
- синхронизация работы ДЕ.
Прежде чем перейти к рассмотрению влияния этих механизмов на силу и скорость сокращения мышц, вспомним определение ДЕ.
ДЕ называется система, включающая α-мотонейрон и иннервируемые им мышечные волокна.
Частота разрядов ДЕ
При увеличении частоты разрядов ДЕ, то есть нервных импульсов, поступающих из ЦНС к мышце, происходит переход от слабого одиночного сокращения к сильным тетаническим сокращениям мышечных волокон.
Число активных ДЕ
Число ДЕ, активных в процессе сокращения мышцы, определяется «принципом размера» или правилом Хенеманна. Установлено, что имеется стабильный порядок рекрутирования ДЕ: вначале рекрутируются ДЕ S типа, иннервируемые мотонейронами, имеющими небольшой диаметр аксона. По мере усиления сокращений начинают рекрутироваться ДЕ FR типа, содержащие быстрые неутомляемые волокна, затем - ДЕ FF типа, содержащие быстрые утомляемые волокна, иннервируемые мотонейронами, имеющими самый большой диаметр аксона. С точки зрения механики, этот принцип очень целесообразен, так как создается возможность тонкой градации мышечной силы во всем физиологическом диапазоне.
Принцип размера позволяет объяснить факты, полученные эмпирическим путем. Все спортсмены, применяющие силовые упражнения, хорошо знают, что, используя небольшие отягощения, невозможно эффективно наращивать силу мышц. Для развития силовых способностей необходимо применять отягощения, близкие к максимальным.
Эту закономерность можно объяснить следующим. При развитии силовых способностей, если применяются небольшие отягощения, рекрутируются только мышечные волокна I типа, так как мышце нет необходимости развивать высокий уровень силы. Для преодоления субмаксимальных или максимальных отягощений мышца должна развить максимально возможную силу. Поэтому в сокращение вовлекаются все типы мышечных волокон, особенно IIB типа, дающие в процессе тренировочных воздействий максимальное увеличение поперечного сечения и, как следствие - силы мышц.
Синхронизация работы мышц
Синхронизация работы ДЕ - увеличение силы тяги мышцы за счет одновременной активации большого количества мышечных волокон. Исследованиями установлено, что тренировка силовой направленности повышает степень синхронизации работы ДЕ (А.С. Солодков, Е.С. Сологуб, 2001).
4.4. Биомеханические факторы, определяющие силу и скорость сокращения мышцы
Существует ряд факторов, влияющих на проявление силы и скорости сокращения мышцы в преодолевающем и уступающем режимах сокращения. К этим факторам относятся:
- длина мышцы;
- режим работы мышцы;
- значение внешней силы.
4.4.1. Длина мышцы
Еще в 1895 году М. Бликс показал, что при растягивании активной поперечно-полосатой мышцы под воздействием внешней нагрузки ее сила вначале возрастает, а затем уменьшается. Максимум силы получил название максимума Бликса, а длина, при которой он достигался - длины покоя. Последующие исследования показали, что появление максимума связано с особенностями строения мышцы (наличие мышечных волокон (сократительного компонента) и соединительно-тканных образований (упругого компонента мышцы)). Если мышцу освободить от нагрузки, она укоротится до определенной длины. Такая длина получила название равновесной.
Установлено, что у человека вид зависимости «длина-сила» активных мышц определяется соотношением сократительного и упругого компонентов. У мышц, богатых соединительно-тканными образованиями (например, у мышц нижних конечностей человека), зависимость «длина-сила» является монотонно-возрастающей (рис. 4.1а). Если соединительно-тканных образований мало, то кривая имеет минимум (рис. 4.1б).
Рис. 4.1. Зависимость «длина-сила» для мышц нижних (а) и верхних (б) конечностей. Обозначения: А - равновесная длина, Б - длина покоя
Из этого следует, что при одной и той же длине мышцы нижних конечностей при растягивании способны развить большую силу, чем мышцы верхних конечностей.
4.4.2. Характер работы мышц
Режим сокращения мышцы
Известно, что мышца может работать в нескольких режимах сокращения:
- преодолевающем (концентрическом) - длина мышцы уменьшается;
- уступающем (эксцентрическом) - длина мышцы увеличивается;
- изометрическом - длина мышцы не изменяется.
Установлено, что на силу, развиваемую мышцей, влияет режим сокращения. Если возбужденную мышцу растягивать и регистрировать силу при определенных значениях длины или, наоборот, растянув, дать возможность укорачиваться, то окажется, что при эксцентрическом режиме сокращения (мышца растягивается) при одних и тех же значениях длины мышца развивает большее усилие, по сравнению с концентрическим режимом.
Предшествующий режим сокращения мышцы
Работа мышц при выполнении движений человеком значительно отличается от таковой при имитации ее в лабораторных условиях. Это связано с тем, что обычно в движениях укорочению мышцы предшествует ее растяжение. Еще И.М. Сеченовым (1901) было отмечено, что мышца, сокращаясь в преодолевающем режиме, способна развить большую силу, если этому сокращению предшествовала работа в уступающем режиме (то есть мышца была предварительно растянута). Такой режим работы получил название «баллистический». Последующие исследования подтвердили этот факт. Следует отметить, что техника движений спортсменов учитывает этот фактор повышения силы мышц. Так, например, в метании копья, спортсмен перед выполнением финального усилия находится в позе «натянутого лука», то есть значительно растягивает основные мышцы, чтобы их сокращение в концентрическом режиме было более сильным. Та же закономерность характерна для техники бега и ходьбы, толкания ядра, метания диска, прыжка в высоту, выпрыгивания вверх с места, приседания со штангой.
4.4.3. Значение внешней силы
Преодолевающий режим сокращения мышцы (концентрический режим)
Если мышца сокращается, преодолевая внешнюю силу (например, вес груза), то с увеличением веса груза наблюдаются три закономерности:
- увеличивается время от момента стимуляции мышцы до начала ее укорочения;
- уменьшается высота подъема груза;
- скорость сокращения мышцы снижается.
Первыми зависимость между силой и скоростью (зависимость «сила-скорость») укорочения мышц лягушки получили В. Фенн и Б. Марч. Нобелевский лауреат Арчибальд Хилл (1961) посвятил много времени изучению энергетических процессов, протекающих в мышце. Ему удалось получить соотношение между скоростью сокращения мышцы и силой, которое носит теперь его имя. Оно называется «характеристическое уравнение Хилла».
где: P0 - максимальная масса груза, при которой не происходит укорочения мышцы; P - масса груза; a,b - константы.
Проведенные впоследствии эксперименты на мышцах человека подтвердили данные, полученные А. Хиллом: чем больше внешняя сила, тем ниже скорость сокращения мышцы. Исследования спортивных движений показали, что между дальностью метания и весом ядер наблюдается зависимость, близкая к гиперболической. То есть, чем больше вес снаряда, тем меньше результат.
Эксцентрический режим
Следует отметить, что изучение зависимости «сила-скорость» (концентрический режим) привлекало внимание многих исследователей, в то время как эксцентрический режим оставался менее исследованным. А. Хилл находил, что при эксцентрическом режиме сила мышцы возрастает с увеличением скорости растяжения. П.В. Коми (1973) использовал специальный динамометр для регистрации усилий, развиваемых двуглавой мышцей плеча человека в концентрическом и эксцентрическом режимах сокращения мышцы. Полученные результаты свидетельствуют о том, что при работе двуглавой мышцы плеча в концентрическом режиме зависимость «скорость-сила» может быть описана уравнением гиперболы. При эксцентрическом режиме сокращения увеличение скорости растяжения мышцы приводит к увеличению силы, развиваемой мышцей. Однако последующие исследования, проведенные при исследовании локомоций человека, не подтвердили этого. Дж. Моррисон (J.B. Morrison, 1970) изучал изменение длины, скорости сокращения мышц и силы при различных локомоциях. Он установил, что с ростом скорости растяжения у большинства мышц проявляемая ими сила быстро уменьшается до нуля.
Еще более убедительны исследования, проведенные П.В. Коми (P.V. Komi, 1992). На рис. 4.2 представлено изменение усилий, развиваемых в области ахиллова сухожилия во время фазы опоры бега со скоростью 5,8 м/с.
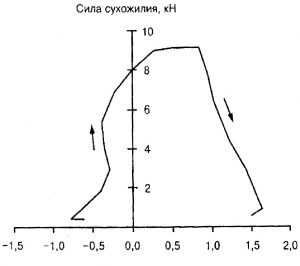
Рис. 4.2. Сила, развиваемая трехглавой мышцей голени во время фазы опоры в беге со скоростью 5,8 м/с (P.V.Komi, 1992)
Отрицательная скорость сокращения мышц соответствует эксцентрическому режиму, положительная - концентрическому. Данные, представленные на графике, свидетельствуют о том, что в фазу растяжения мышцы большие значения усилий, развиваемых мышцей, соответствуют небольшим скоростям растяжения мышцы. В концентрическом режиме большие значения силы также развиваются при небольших значениях скорости.
Таким образом, в настоящее время в мышечной механике достоверно установлена следующая закономерность: с увеличением значений внешней силы скорость сокращения мышц уменьшается. Зависимость нелинейная (гиперболическая).
Сокращение скелетных мышц человека
Описан процесс сокращения скелетных мышц человека, который состоит из ряда этапов. Первый этап связан с поступлением нервного импульса по аксону мотонейрона к мышечному волокну. Второй этап заключается в возникновении потенциала действия и распространении его вдоль мышечного волокна. В результате третьего этапа в саркоплазму выделяются ионы кальция, что приводит к началу взаимодействия толстого и тонкого филаментов. Четвертый этап представляет собой скольжение тонких филаментов относительно толстых, что составляет собственно сокращение мышцы.
Давайте теперь разберемся в механизме сокращения мышцы, точнее в механизме сокращения мышечных волокон, а еще более точно в механизме сокращения миофибрилл или другими словами, в механизме сокращения саркомера. Этот процесс можно условно разделить на несколько этапов.
Поступление нервного импульса к мышечному волокну
Чтобы сократиться, мышца должна получить сигнал из центральной нервной системы (ЦНС). Такими сигналами являются импульсы, поступающие по мотонейрону к мышце.
Вспомним, что при подходе к мышце аксон мотонейрона ветвится, то есть пускает веточки к мышечным волокнам. Если такого соединения нет, мышечное волокно сокращаться не будет и постепенно атрофируется.
Возникновение потенциала действия
После того, как по аксону мотонейрона к мышечным волокнам приходит импульс, из него в области соединения выделяется ацетилхолин. Выделение этого нейромедиатора (ацетилхолина) приводит к протеканию ряда процессов, в результате которых меняется полярность сарколеммы мышечного волокна. Это называется деполяризацией сарколеммы мышечного волокна. В результате развивается потенциал действия.
Выделение ионов кальция
Потенциал действия через отверстия в сарколемме «проникает» внутрь мышечного волокна и через Т-трубочки достигает саркоплазматического ретикулума (то есть происходит деполяризация не только мембраны мышечного волокна, но и мембран Т-трубочек и саркоплазматического ретикулума). Это в конечном счете приводит к выделению из саркоплазматического ретикулума ионов кальция в саркоплазму мышечного волокна (рис. 1).
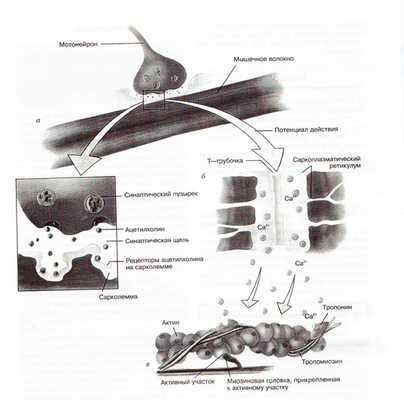
Рис.1. Механизм сокращения скелетных мышц
Затем ионы кальция соединяются с тропонином (тропонин - один из белков тонкого филамента). Этот белок имеет шарообразную форму и расположен в тонком филаменте регулярно через определенные расстояния. После соединения с ионами кальция, тропонин меняет свою конфигурацию и приподнимает длинные тропомиозиновые трубки. Когда мышца неактивна, длинные трубки белка тропомиозина закрывают активные центры на актине. После того как тропомиозиновые трубки приподнимаются, на актине открываются активные центры. К ним теперь могут прикрепляться миозиновые головки.
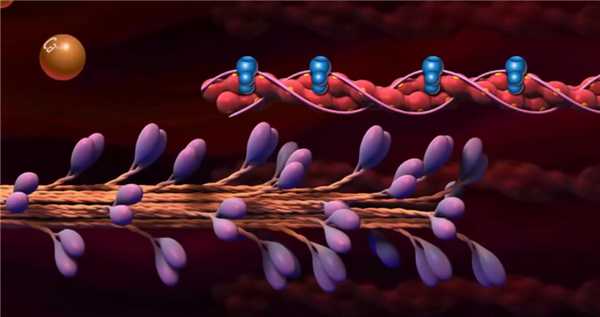
Сокращение саркомера (гребковая гипотеза, теория скользящих нитей)
Когда миозиновая головка толстого филамента прикрепляется к тонкому филаменту, между толстым и тонким филаментами начинается взаимодействия (говорят: «Образуется поперечный мостик» (рис. 2). При взаимодействии с актином каждая миозиновая молекула ежесекундно расщепляет с выделением энергии до 10 молекул АТФ. За счет энергии, высвобождающейся при расщеплении АТФ, миозиновая головка поворачивается и тянет тонкий филамент в направлении центра саркомера. Это приводит к скольжению толстого и тонкого филаментов относительно друг друга. В конце гребка (поворота) к миозиновой головке присоединяется новая молекула АТФ, что приводит к отделению головки от актина и присоединению её к новому активному участку тонкого филамента. Многократное повторение этого процесса приводит к тому, что расстояние между Z-дисками уменьшается. Следовательно, происходит уменьшение длины саркомера. Одновременное сокращение всех саркомеров, расположенных последовательно вдоль миофибриллы приводит к уменьшению её длины, длины мышечного волокна и всей мышцы в целом. Мышца работает в преодолевающем режиме.
Прекращение импульсов, поступающих от мотонейрона к мышечному волокну приводит к расслаблению мышцы.
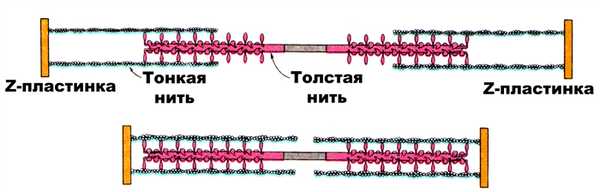
Рис.2. Схема, иллюстрирующая взаимодействие толстого и тонкого филаментов (Л. Страйер, 1985)
Биомеханика мышечного сокращения
Сила — векторная величина, являющаяся мерой механического воздействия на материальную точку или тело со стороны других тел или полей (Б. М. Яворский, А. А. Детлаф, 1974). Сила полностью задана, если указаны ее численное значение, направление и точка приложения.
В теории и методике физического воспитания рассматривают физическое качество силу как способность человека напряжением мышц преодолевать механические и биомеханические силы, препятствующие действию (Зациорский В. М., 1972; Л. П. Матвеев, 1991).
Мышцы могут проявлять силу: без изменения своей длины (изометрический режим), при уменьшении длины (изотонический режим), при удлинении (эксцентрический режим), при использовании специальной аппаратуры возможно соблюдение изокинетического режима (в ходе сокращения мышц соблюдается либо постоянная скорость, либо сила).
Силовое проявление мышцы зависит от:
— интенсивности активации мотнейронного пула спинного мозга данной мышцы;
— количества активированных двигательных единиц и мышечных волокон;
— количества миофибрилл в каждом мышечном волокне;
— скорости сокращения миофибрилл, которая зависти от активности миозиновой АТФ-азы и величины внешнего сопротивления;
— законов механики мышечного сокращения (сила — длина мышцы, сила — скорость сокращения);
— начального состояния исполнительного аппарата (утомленные мышечные волокна демонстрируют меньшую силу).
Спортсмен при желании сократить какую-либо мышцу активизирует соответствующий двигательный нейрон в коре головного мозга, который посылает импульсы в спинной мозг к мотонейронному пулу, обслуживающему данную мышцу. Поскольку в мотонейронном пуле размеры мотонейронов различаются, то при низкой частоте импульсации из ЦНС могут активироваться только низкопороговые мотонейроны. Каждый мотонейрон иннервирует свои мышечные волокна. Поэтому активация мотонейрона приводит к рекрутированию или возбуждению соответствующих мышечных волокон. Каждое активное мышечное волокно под влиянием электрических импульсов выпускает из СПР ионы кальция, которые снимают ингибитор с активных центров актина. Это обеспечивает образование актин-миозиновых мостиков и начало их поворота и мышечного сокращения. На поворот мостиков и отсоединение актина от миозина тратится энергия одной молекулы АТФ. Продолжительность работы мостика составляет 1 мс. Вероятность образования мостиков зависит от взаимного расположения между собой нитей актина и миозина, отсюда возникает зависимость сила — длина активной мышца, а также от скорости взаимного перемещения (скольжения) их одной по отношению к другой, соответственно, имеем зависимость «сила-скорость».
Зависимость «сила — длина активного мышечного волокна» определяется, как правило, относительным расположением между собой головок миозина и активных центров актина. Максимальное количество мостиков возникает при некоторой средней длине мышцы. Отклонение от этой длины в большую или меньшую сторону ведет к снижению силовых проявлений мышечного волокна (мышцы). Однако, в случае растяжения некоторых мышц, еще не в активном состоянии, могут возникать значительные силы сопротивления растяжению, например, в мышцах сгибателях голеностопного или лучезапястного сустава. Эти силы связаны с растяжением соединительных тканей, например, перемезиума. В биомеханике в таком случае говорят о параллельном упругом компоненте мышцы. Упругостью обладают сухожилия, зет-пластинки саркомеров и нити миозина, к которым прикреплены головки. Такую упругость называют последовательной упругой компонентой.
Наличие последовательной упругой компоненты в мышечных волокнах приводит к тому, что с ростом числа рекрутированных МВ увеличивается жесткость мышцы -коэффициент упругости (В. М. Зациорский с соав., 1981).
Растягивание активной мышцы приводит не только к накоплению энергии упругой деформации в последовательной упругой компоненте, но и к прекращению работы мостиков, а именно, они перестают отцепляться за счет энергии молекул АТФ. Разрыв мостиков происходит благодаря действию внешней — механической силы. В итоге отрицательная работа мышц выполняется с очень высоким коэффициентов полезного действия, с минимальными затратами АТФ, а значит и кислорода.
Читайте также: