Волосковые клетки - киноцилий. Физиология полукружных каналов
Добавил пользователь Евгений Кузнецов Обновлено: 01.02.2026
А киноцилий это особый вид ресничка на вершине волосковые клетки расположен в сенсорном эпителии позвоночное животное внутреннее ухо.
Содержание
Анатомия человека
Киноцилии находятся на апикальной поверхности волосковые клетки и участвуют как в морфогенезе пучка волос, так и в механотрансдукция. Вибрации (движения или звуковые волны) вызывают смещение пучка волос, что приводит к деполяризации или гиперполяризации волосковой клетки. Деполяризация волосковых клеток в обоих случаях вызывает преобразование сигнала через нейротрансмиттер релиз.
Роль в морфогенезе пучка волос
Каждая волосковая клетка имеет одну, микротрубчатый киноцилии. Перед морфогенез Пучка волос киноцилия находится в центре апикальной поверхности волосковой клетки, окруженной 20-300 микроворсинками. Во время морфогенеза пучка волос киноцилия перемещается к периферии клеток, определяя ориентацию пучка волос. Поскольку киноцилия не двигается, окружающие ее микроворсинки начинают удлиняться и образовывать актин. стереоцилии. У многих млекопитающих киноцилии регрессируют, когда пучок волос созревает. [1]
Слуховая система
Движение пучка волос в результате эндолимфа [2] Поток вызовет открытие калиевых каналов в стереоцилиях. В основном это происходит из-за силы притяжения, которую стереоцилии оказывают на соседние стереоцилии через соединительные звенья, которые удерживают стереоцилии вместе (обычно от самых высоких до самых коротких), и это приводит к деполяризации волосковой клетки. Этот паттерн деполяризации не следует путать с более распространенной деполяризацией, которая включает приток Na + в клетку, в то время как K + каналы остаются закрытыми. Состав эндолимфы напоминает состав внутриклеточная жидкость (больше K + и меньше Na +) более точно по сравнению с его аналогом, перилимфа который напоминает внеклеточной жидкости (больше Na + и меньше K + по сравнению с внутриклеточным матриксом). Эта деполяризация откроет управляемые напряжением кальциевые каналы. Затем приток кальция заставляет клетку высвобождать везикулы, содержащие возбуждающие нейротрансмиттеры, в синапс. Постсинаптический нейрит затем посылает потенциал действия в спиральные ганглии Гарда. В отличие от волосковых клеток crista ampullaris или пятна мешочка и матрикса, волосковые клетки кохлеарный проток не обладают киноцилиями.
Вестибулярный аппарат
Киноцилии присутствуют в crista ampullaris полукружных протоков и сенсорных пятнах мешок и мешочек. Одна киноцилия - самая длинная ресничка, расположенная на волосковой клетке рядом с 40-70 стереоцилиями. Во время движения тела волосковая клетка деполяризуется, когда стероцилии движутся к киноцилии. Деполяризация волосковой клетки вызывает высвобождение нейромедиатора и увеличение частоты срабатывания черепной нерв VIII. Когда стероцилии отклоняются от киноцилии, волосковая клетка гиперполяризуется, уменьшая количество высвобождаемого нейромедиатора, что снижает частоту возбуждения VIII черепного нерва. [3]
Анатомия рыб и лягушек
Апикальная поверхность сенсорного рыбы волосковая клетка обычно имеет множество стереоцилии и единственная, гораздо более длинная киноцилия. В отличие от млекопитающих киноцилия не регрессирует и остается частью пучка волос после созревания волосковых клеток. Прогиб стереоцилий по направлению к киноцилии или от нее вызывает увеличение или уменьшение скорости возбуждения сенсорных нейрон иннервируя волосковая клетка на его базальной поверхности.
Волосковые клетки у рыб и некоторых лягушек используются для обнаружения движения воды вокруг своего тела. Эти волосковые клетки заключены в желеобразный выступ, называемый купула. Следовательно, волосковые клетки не видны и не появляются на поверхности кожи рыб и лягушек.
Полукружные каналы
От маточки под прямыми углами друг к другу отходят три полукружных канала. Расположения их таково, что каждый из них реагирует на угловое ускорение головы, т.е. ее внезапный поворот в одной из трех плоскостей — фронтальной, сагиттальной и горизонтальной.
В каждом канале имеется расширенный участок с чувствительными волосковыми клетками — ампула. Реснички этих клеток покрыты студенистым колпачком — купула. Купула выступает в просвет канала и легко смещается при движениях эндолимфы, заполняющей канал. Смещение купулы приводит к возбуждению погруженных в эндолимфу волосковых клеток. На каждой такой клетке расположена одна крупная киноцилия и множество мелких — стереоцилий. Ориентация всех волосковых клеток, в пределах каждой ампулы, одинакова. Поэтому, при смещении купулы в одном направлении, возникает возбуждение клеток, а в противоположном направлении — торможение.
Восковые клетки реагируют на угловое ускорение, возникающее при внезапных поворотах головы. При движении полукружного канала перемещение заполняющей его эндолимфы в силу инерции запаздывает. Во время вращения головы, а значит, полукружных каналов и купулы, эндолимфа стремится сохранить прежнее положение в пространстве, и вследствие этого она давит на купулу, смещая ее. В результате этого смещения стереоцилии сгибаются по направлению к киноцилии, и волосковые клетки подвергаются возбуждению. При внезапном прекращении вращения головы полукружные каналы испытывают отрицательное угловое ускорение. В течение короткого времени после остановки головы, полукружных каналов и связанных с ними купул эндолимфа продолжает двигаться и вновь смещает купулу. Так как при этом стереоцилии отклоняются от киноцилий, волосковые клетки в ответ на отрицательное угловое ускорение подвергаются торможению.
От полукружных каналов поступает информация о плоскости, в которой происходит угловое ускорение (положительное или отрицательное) и его величине.
Информация о плоскости вращения коферуется в виде сигналов от какого-либо определенного канала (верхнего, заднего или горизонтального), так как каждый канал реагирует на угловое ускорение только в одной из плоскостей. Степень смещения купулы и, следовательно, частота импульсации в вестибулярном нерве, иннервирующем волосковые клетки, зависит от величины ускорения.
Центральные вестибулярные пути
Волосковые клетки вестибулярного аппарата иннервируются волокнами вестибулярного нерва. Эти волокна идут в составе вестибулокохлеарного нерва к продолговатому мозгу, где и заканчиваются в вестибулярных ядрах. Отростки нейронов этих ядер идут к мозжечку, ретикулярной формации и спинному мозгу — двигательным центрам, управляющим положением тела при движениях благодаря информации от вестибулярного аппарата, проприорецепторов шеи и органов зрения.
Поступление вестибулярных сигналов к зрительным центрам имеет первостепенное значение для важного глазодвигательного рефлекса — нистагма. Благодаря нистагму взор при движениях головы фиксируется на неподвижном предмете. Во время вращения головы глаза медленно поворачиваются в обратную сторону, и поэтому взор фиксирован на определенной точке. Если угол вращения головы больше, чем тот, на который могут повернуться глаза, то они быстро перемещаются в направлении вращения и взор фиксируется на новой точке. Это быстрое движение и есть нистагм. При повороте головы глаза попеременно совершают медленные движения в направлении поворота и быстрые в противоположном настроении.
1. «Основы физиологии». Под редакцией П. Стерки. Перевод с англ. М., 1984. Стр. 124—141.
Физиология вестибулярного анализатора
Периферическая часть этого анализатора носит название вестибулярного аппарата, поскольку расположена в преддверии улитки. В каждом ухе вестибулярный аппарат включает по пять структур: по три полукружных канала, маточке (утрикулюсу) и мешочку (сак- кулюсу).
Полукружные каналы. Каждый из трех полукружных каналов перепончатого лабиринта на большей части своего пути идет отдельно от других, но есть участки, принадлежащие соседним каналам. Поэтому каждый из них образует неполный круг, что и обусловило название — полукружный. Диаметр такого «полукруга» примерно 6,5 мм.
Соединительнотканные трубочки, образующие каналы, имеют на поперечных срезах диаметр около 0,25 мм. Все три канала замыкаются на утрикулюсе. В месте слияния с ним каждый канал расширяется до 2 мм. Такое расширение называется ампулой. В каждой из трех ампул полукружных каналов сосредоточены гребешки — рецепторные органы, реагирующие на угловые перемещения. Гребешок выступает внутрь ампулы примерно на 0,25 мм. Он выстлан рецепторным эпителием, площадь которого в гребешке достигает 1 мм 2 . В состав рецепторного (сенсорного) эпителия входят от 7000 до 7500 рецепторных (волосковых) и на порядок больше опорных клеток. Каждая волосковая клетка окружена 6—10 опорными.
Между рецепторными и опорными клетками существуют разнообразные типы клеточных контактов. Апикальные полюсы связаны плотными контактами. Ближе к базальным полюсам расположена область слипания клеток в виде ленты и отдельных десмосом. Между опорными клетками имеются щелевые контакты, осуществляющие их электрическое взаимодействие. Полагают, что у млекопитающих опорные клетки не играют важной роли в дифференцировке рецепторных клеток, но они поддерживают целостность сенсорного эпителия, а значит, и всего рецепторного органа. У рыб они служат камбием, из которого в течение жизни развиваются как опорные, так и рецепторные клетки. Сказанное об опорных клетках относится как к гребешкам, так и к пятнам.
Заметим также, что сенсорный эпителий обоих рецепторных органов равновесия (гребешка и пятна) отделен от подлежащей соединительной ткани базальной мембраной. Соединительная ткань, составляя структурную основу гребешка и пятна, служит своеобразной «подушкой» сенсорному эпителию. Она пронизана кровеносными капиллярами и нервными волокнами.
Вернемся к гребешкам. Со стороны эндолимфатического пространства волосковые клетки покрыты купулой (рис. 17.11), которая и придает всему этому рецепторному органу форму миниатюрного гребня. Купула тянется от сенсорного эпителия гребешка до противоположной стенки ампулы, перекрывая весь ее просвет. Высота купулы около 1 мм. Волоски рецепторных клеток глубоко вдаются в вещество купулы и вносят некоторый вклад в ее упругость (эластичность), которая обусловлена главным образом аморфным желеобразным веществом и переплетающимися фибриллами. Купула имеет примерно такую же плотность, как эндолимфа.
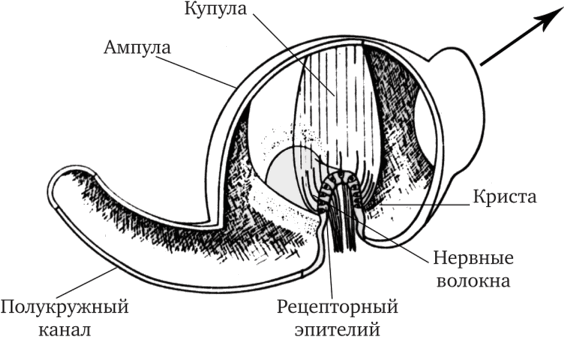
Рис. 17.11. Вскрытая ампула полукружного канала (по Линдеману, 1969).
Стрелка в правом углу рисунка указывает местоположение утрикулюса
При вращении головы вокруг оси, проходящей через центр круга, образованного полукружным каналом, реагирующим на такое вращение, его стенки тоже вращаются, благодаря тому что перепончатый лабиринт связан с костным соединительнотканными тяжами. Вращательное движение стенок полукружного канала не воспроизводится эндолимфой моментально, так как она в силу своей инерции движется с отставанием. Это приводит к сдвигу частиц эндолимфы относительно тех точек на стенках полукружного канала, к которым они прилежали до вращения (рис. 17.12). Перемещаясь относительно стенок канала, эндолимфа начинает оказывать давление на купулу, которая перегораживает просвет ампулы. Испытывая давление движущейся эндолимфы, купула смещается относительно поверхности сенсорного эпителия и наклоняет волоски рецепторных клеток, вдающиеся в купулу.
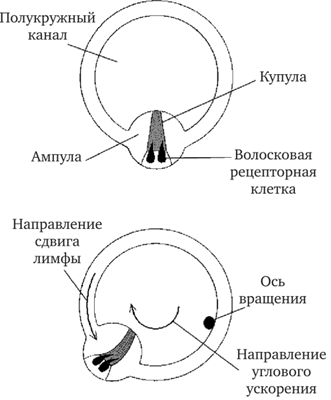
Рис. 17.12. Схема преобразования угловых ускорений в полукружном канале (по Хадспету, 1983):
вверху — положение купулы в покое; внизу — положение купулы при вращении
В зависимости от направления сдвига купулы наклон волосков вызывает либо возбуждение, либо торможение клеток, которым они принадлежат. При прекращении вращения купула за счет присущей ей упругости возвращается в исходное положение. Время возвращения занимает от единицы до десятков секунд в зависимости от величины действовавшего ускорения.
Каждый из трех полукружных каналов испытывает максимальную стимуляцию при вращении головы вокруг оси, перпендикулярной плоскости, в которой он лежит. Вращение вокруг осей, расположенных под углами к этой плоскости, меньшими 90°, будет одновременно стимулировать либо еще один, либо еще два других полукружных канала. Однако степень стимуляции гребешков каждого из задействованных каналов будет неодинакова.
Наличие трех парных каналов (в левом и правом лабиринтах) позволяет человеку и животному регистрировать любые вращения головы. В центральной части вестибулярного анализатора и ассоциативных областях коры больших полушарий головного мозга проводится векторный анализ, в ходе которого определяется пространственная ориентация оси вращения.
Линейные ускорения, включая ускорение свободного падения, не возбуждают рецепторы гребешков, поскольку при прямолинейном движении круговой ток эндолимфы не возникает. На прямолинейные перемещения реагирует другой рецепторный орган равновесия — пятна (макулы). Они расположены в утрикулюсе и саккулюсе.
Утрикулюс и саккулюс. Утрикулюс представляет собой вытянутый сплюснутый мешок в форме эллипсоида длиной около 4 мм и объемом примерно 8 мм 3 . Саккулюс имеет форму сплюснутого шара диаметром около 2,6 мм и объем примерно 2 мм 3 .
В утрикулюсе и саккулюсе одна стенка утолщена — такие утолщения называются пятнами. Рецепторный эпителий пятен содержит волосковые и опорные клетки. В утрикулюсе пятно имеет площадь в среднем 4 мм 2 и содержит до 33 тысяч рецепторных клеток. В саккулюсе площадь пятна вдвое меньше (примерно 2 мм 2 ), соответственно, в нем меньше и волосковых клеток — до 19 тысяч. Толщина каждого пятна составляет несколько десятков микрон. Ма- кула утрикулюса имеет форму отпечатка копыта лошади, а макула саккулюса — форму изогнутого крючка (рис. 17.13).
Со стороны эндолимфатического пространства пятна утрикулюса и саккулюса покрыты отолитовой мембраной (рис. 17.14). Она, как и купула, является кроющей внеклеточной структурой. Толщина отолитовой мембраны до 80—100 мкм. В ней есть органический матрикс, в котором сосредоточено множество кристаллических песчинок — отоконий (рис. 17.15), образующих в совокупности ото- литовый аппарат. Они являются мозаичными кристаллами, состоящими из множества микрокристаллов карбоната кальция. Самые крупные отоконии достигают длины до 30—40 мкм.
Микрокристаллы карбоната кальция, объединенные органическим матриксом, столь хорошо пригнаны друг к другу, что отдельные отоконии ведут себя в поляризованном свете как монолитные кристаллы кальцита. У отоконий плотность в 2,5 раза больше, чем у эндолимфы. Поэтому при действии линейных ускорений, включая ускорение свободного падения, отолитовая мембрана, не связанная жестко с макулой, за счет сил инерции отстает от движения эндолимфы.
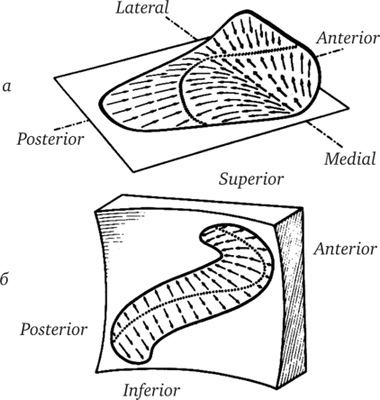
Рис. 17.13. Пространственная ориентация, форма и поляризация макул левого утрикулюса (а) и правого саккулюса [6).
В макулах есть особая область — стриола, в которой направление поляризации рецепторных клеток изменяется на противоположное. Линия перемены поляризации (линия стриолы) обозначена пунктиром. Стрелками указано направление поляризации
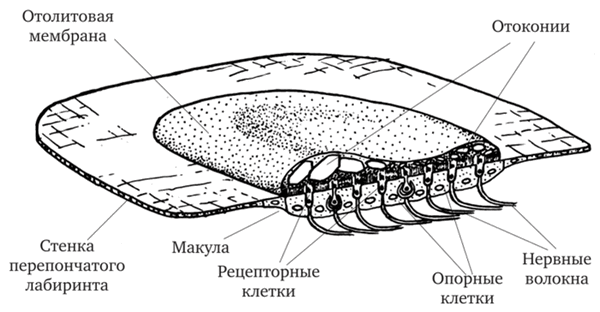
Рис. 17.14. Схема строения утрикулюса млекопитающих (по Д. В. Лычакову, 1988)
Благодаря тому, что пятна утрикулюса и саккулюса расположены в двух взаимно перпендикулярных плоскостях, линейные ускорения любого направления вызывают ответ по крайней мере в одном пятне. Сопоставление сигналов от парных (слева и справа) пятен утрикулюса и саккулюса позволяет центральной нервной системе выполнять векторный анализ и однозначно определять направление линейного ускорения.

Рис. 17.15. Отоконии морской свинки (сканирующая электронная микроскопия, масштаб 10 мкм)
Вторично-чувствующие рецепторы вестибулярного анализатора. Рецепторный эпителий гребешков и пятен содержит два типа волосковых клеток (рис. 17.16). Клетки первого типа имеют грушевидную или кувшинообразную форму и почти полностью окружены чашевидными нервными окончаниями. У клеток второго типа тело цилиндрическое. Базальный полюс каждой из них контактирует с несколькими нервными окончаниями бутоновидной формы.
От апикального полюса любой из рецепторных клеток в эндолимфу выступают от 30 до 100 стереоцилий и одна киноцилия. Последняя всегда длиннее самых длинных стереоцилий и располагается эксцентрично по отношению к их пучку (сбоку от него). Такое расположение свидетельствует о морфологической поляризации волосковых клеток. На схемах и фотографиях направление поляризации обозначается стрелками, идущими от пучка стереоцилий к киноцилии (см. рис. 17.13).
Самые длинные стереоцилии достигают 50 мкм, а их диаметр составляет от 0,1 до 0,8 мкм. Стереоцилии в клетках первого типа толще и длиннее, чем в соседних клетках второго типа. Эти волоски связаны между собой посредством гликокаликса, содержащего миозин и белки семейства кадхеринов. Кроме того, существуют вершинные нити (диаметром около 10 нм и длиной до 150 нм), соединяющие соседние стереоцилии. Макромолекулярные мостики объединяют стереоцилии в пучок. Цитоскелет каждой стереоцилии образован многочисленными (до 3000) актиновыми нитями, идущими параллельно друг другу. Актиновые филаменты связаны между собой и с плазмолеммой стереоцилии. У основания стереоцилии становятся тоньше и образуют корешок, входящий в толщу подмембранного слоя и называемый кутикулой. Она служит платформой для крепления стереоцилий. При действии механического стимула пучок чувствительных волосков смещается как единое целое. Отклонение пучка стереоцилий в сторону киноцилии приводит к деполяризации волосковой клетки, тогда как его противоположное смещение гиперполяризует ее. Сдвиги мембранного потенциала рецепторной клетки пропорциональны амплитуде колебаний пучка стереоцилий. Его пороговое смещение составляет примерно 5 нм.
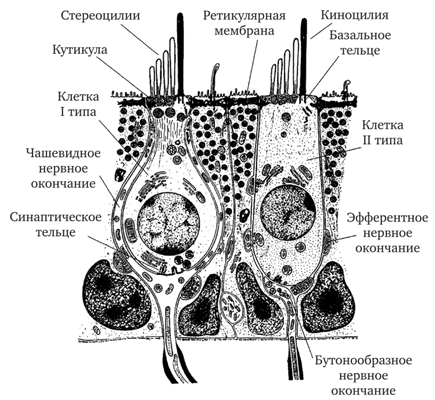
Рис. 17.16. Рецепторные клетки гребешков и пятен
Киноцилия, являющаяся по своей ультраструктуре типичной ресничкой, начинается от базального тельца на апикальном полюсе волосковой клетки, и на некотором расстоянии от него аксо- нема киноцилии приобретает характерную для реснички структуру, включающую 9 пар периферических и одну пару центральных микротрубочек, обозначаемую формулой: 9-2 + 2. Диаметр киноцилии составляет 0,2—0,3 мкм. Роль киноцилий в рецепции пока не вполне ясна. Считается, что они способствуют поляризации рецепторного эпителия в органах равновесия. Способность волоско- вых клеток градуально реагировать на изгиб пучка стереоцилий относительно киноцилии, а также поляризация сенсорного эпителия позволяют уже на уровне гребешков и пятен анализировать как величину, так и направление действующего на организм ускорения.
Трансдукция механических стимулов в волосковой клетке. В отличие от киноцилий, стереоцилии играют важнейшую роль в трансдукции механического стимула. Каждая стереоцилия имеет от 1 до 6 механоэлектрических каналов, которые располагаются у ее вершины. Поскольку диаметр просвета этих каналов (около 0,6 нм) превосходит диаметры гидратированных ионов калия и натрия, их селективность невысокая, а электропроводность большая (до 100 пСм). Примерно через 10 мкс после действия механического стимула каналы открываются. В этом существенное значение имеют вершинные нити. Они непосредственно связаны с механо- электрическими каналами, расположенными в плазмолемме более длинных стереоцилий. Нити, находящиеся на вершинах коротких стереоцилий при наклоне всего пучка тянут на себя молекулярные структуры каналов, принадлежащих соседним более длинным сте- реоцилиям, и открывают каналы для ионов. Чем больше наклон стереоцилий, тем сильнее натяжение нитей и тем продолжительнее период открытого состояния канала. В таком состоянии через канал течет ток порядка нескольких сотен пикоамперов. Он обусловлен главным образом потоками ионов калия и кальция. В первом варианте возникает гиперполяризация, а во втором — деполяризация волосковой клетки.
Такая трансдукция сигнала происходит без участия вторичных мессенджеров, что обеспечивает короткий латентный период в преобразовании механического стимула в электрический ответ. Для него характерна асимметрия: при наклоне пучка стереоцилий к киноцилии (при натяжении вершинных нитей) деполяризация клетки по абсолютной величине превосходит гиперполяризацию, которая возникает при отклонении пучка стереоцилий от киноцилии, что происходит при ослаблении вершинных нитей. Электрический ток (ток трансдукции), возникающий под действием механического стимула (пропорционально ускорениям), течет через апикальный участок плазмолеммы волосковой клетки и вызывает сдвиг ее мембранного потенциала. Его называют рецепторным потенциалом. Он распространяется по плазмолемме волосковой клетки электротонически (с декрементом), поскольку она невозбудима (в ней отсутствуют потенциалзависимые ионные каналы). Достигнув базального полюса волосковой клетки, отчасти затухший РП способен вызвать там выделение медиатора, обеспечивающего передачу сигнала с рецепторной клетки на контактирующие с ним афферентные волокна, которые являются дендритами биполярных клеток, принадлежащих вестибулярному ганглию (ганглию Скарпа).
Медиаторы, осуществляющие синаптическую передачу во вто- рично-чувствующих рецепторах органов равновесия, до сих пор не идентифицированы. По мнению одних исследователей, медиатором служит у -аминомасляная кислота — NH2(CH2)3COOH, тогда как другие склонны отдать эту роль глутамату — соли глутаминовой кислоты — COOH(CH2)2CH(NH2)COOH. Разрешение вопроса о медиаторе в синаптической передаче афферентного сигнала осложняется тем, что волосковая клетка имеет наряду с афферентной и богатую эфферентную иннервацию. Эфферентные волокна образуют на плазмолемме рецепторной клетки синапсы, осуществляя тем самым обратную связь центральной нервной системы (главным образом, ядра Дейтерса и мозжечка) с рецепторами. В эфферентных синапсах медиаторами могут служить ацетилхолин, глицин, серотонин. Благодаря им регулируются функционирование и чувствительность вестибулярных рецепторных органов.
Вестибулярная сенсорная система
Общие представления о морфологических особенностях и функции вестибулярного анализатора. Все живые организмы развивались и живут в условиях действия на них силы тяжести, или гравитационного поля Земли, которое практически постоянно по величине и направлению к поверхности Земли и действует на любое тело на Земле. В таких условиях живые организмы на основе оценки направления действия гравитационного поля (то есть определения «верха», «низа», величины отклонения положения тела от нормального) должны уметь принимать определенное положение по отношению к линии действия силы тяжести, а также уметь координировать свою двигательную активность, при которой нередко требуется значительное отклонение положения тела от нормального.
Для оценки направления действия гравитационного поля, т. е. для определения положения организма в трехмерном пространстве
и возник вестибулярный (статокинетический) анализатор, или орган статического и динамического равновесия. У человека его периферический отдел представлен вестибулярным аппаратом, или вестибулярным (преддверным) лабиринтом.
Аппарат расположен в лабиринте пирамиды височной кости. В нем различают костную и перепончатую части. Внутри последней находится эндолимфа (ее вязкость в 2—3 раза выше вязкости воды), а между костным и перепончатым лабиринтом находится перилимфа. Костный и заполняющий его перепончатый лабиринты состоят из преддверия, трех полукружных каналов и улитки, которая не имеет отношения к органу равновесия.
Преддверие (vestibulum, вестибулюм) представляет собой овальную полость, в которой содержится два мешочка — эллиптический (маточка, utriculus) и сферический (собственно мешочек, sacculus), играющие важную роль в оценке положения тела в пространстве.
Полукружные каналы располагаются в трех взаимно перпендикулярных плоскостях: передний канал — в сагиттальной плоскости, задний — во фронтальной, а латеральный — в горизонтальной. Такое расположение позволяет осуществлять контроль за различными поворотами головы в любой плоскости.
Внутри полукружных каналов залегают одноименные полукружные перепончатые протоки. Одна из ножек каждого перепончатого протока, как и костного, расширяясь, образует перепончатую ампулу (переднюю, заднюю и латеральную). Оба конца каждого полукружного протока открываются в утрикулюс. Соседние ножки переднего и заднего каналов соединяются, образуя общую костную ножку, поэтому три канала открываются в утрикулюс пятью отверстиями.
Участки мешочков, содержащие чувствительные к гравитации и к линейным ускорениям клетки, называются пятнами, или маку- лами. Аналогичные участки ампул полукружных перепончатых каналов, содержащие клетки, чувствительные к угловым ускорениям или замедлениям, называются гребешками, или кристами.
Вестибулярный анализатор предназначен для пространственной ориентации человека (в том числе при активных и пассивных движениях) по отношению к гравитационному полю Земли, для оценки основных характеристик гравитационного поля, для определения положения головы и шеи по отношению к туловищу, для восприятия линейных и угловых ускорений или замедлений, а также для сохранения равновесия. На основе полученной информации от вестибулярного аппарата при пассивном движении корковые отделы вестибулярного анализатора запоминают направление движения, повороты и пройденное расстояние.
При активных движениях вестибулярная система получает, передает и анализирует информацию об ускорениях или замедлениях, возникающих в процессе прямолинейного или вращательного движения, а также при изменении положения головы в пространстве. При равномерном движении или в условиях покоя рецепторы вестибулярной сенсорной системы не возбуждаются.
В целом, вся информация, идущая от вестибулярного аппарата в мозг, используется, прежде всего, для регуляции позы и локомоций, т. е. в управлении скелетной мускулатурой.
Следует отметить, что вопрос о значении вестибулярной системы в пространственной ориентировке животных и человека обсуждается в течение длительного времени в связи с гипотезой И. Ф. Циона (1879) о вестибулярном аппарате как анализаторе пространства. В настоящее время существует несколько точек зрения о механизмах ориентации в пространстве, общей чертой которых является представление о комплексном участии ряда сенсорных систем в осуществлении этой функции.
Согласно одним представлениям, высшие млекопитающие животные и человек ориентируются в пространстве в основном с помощью зрения и лабиринтов. По мнению других, пространственный анализ представляет собой комплексную функцию динамического ансамбля многих анализаторов, деятельность которых интегрируется двигательным анализатором. Третьи рассматривают ориентацию организма в пространстве как процесс автоматического управления. Задача рецепторов лабиринта в этом процессе заключается в слежении за изменениями в пространстве положения анатомофизиологического центра черепа, а не всего тела в целом. Информация об изменениях в пространстве положения этого центра, поступающая от вестибулярного аппарата, обогащается информацией других модальностей — зрительной, слуховой, а также информацией о рефлексах положения.
Таким образом, все теории сходятся на представлении о комплексном участии сенсорных систем при ориентации в пространстве с непременным участием вестибулярного аппарата как необходимого условия для формирования адекватного поведения человека и животных в окружающей среде.
Чувствительность вестибулярной системы здорового человека очень высока: отолитовый аппарат дает возможность воспринять ускорение прямолинейного движения, равное всего 2 см/с 2 . Порог различения наклона головы в сторону — всего около 1 °, а вперед и назад — 1,5—2 °. Кристы полукружных каналов позволяют человеку замечать ускорения вращения на 2—3 см/с 2 .
Общие принципы функционирования вестибулярного аппарата. Для ориентации по отношению к гравитационному полю Земли большинство животных имеет датчик положения тела, т. е. специальный отолитовый орган, построенный по одному и тому же принципу. Орган равновесия, или гравирецептор, состоит из двух частей: «пробной массы», т. е. отолита, обладающего большей или
меньшей свободой перемещения в пределах органа, и системы рецепторов, воспринимающих положение или перемещение этой массы в органе. Любое отклонение в положении тела сопровождается смещением пробной массы (отолита, отоконий, отолитовой мембраны), возбуждающей соответствующую группу рецепторов органа равновесия. Сигналы этих рецепторов обрабатываются ЦНС, которая и посылает командный сигнал к мышцам, корректирующим положение тела.
Структуры, ответственные за восприятие угловых ускорений, представляют собой каналы, заполненные жидкостью, в стенках которых располагаются рецепторные клетки. Когда животное или человек перемещается в пространстве, жидкость в канале приходит в движение и отклоняет реснички клеток в направлении, которое зависит от направления ускорения. У человека, как и у высших млекопитающих животных, вестибулярный аппарат содержит одновременно и гравирецепторы, и рецепторы ускорения.
Эволюционные аспекты развития гравитационной сенсорной системы. У многих беспозвоночных (за исключением насекомых) универсальным органом восприятия гравитационного поля является статоцист — замкнутая полость, заполненная жидкостью и содержащая статолит или статоконии (секреты стаоциста или привнесенные из окружающей среды, например у омаров и креветок — это песчинки).
Полость статоциста выстлана реснитчатыми клетками, которые у одних животных расположены равномерно, у других — образуют скопления, называемые макулами. При изменении положения тела происходит смещение статокониев в статоцисте, которое, в свою очередь, вызывает отклонение ресничек рецепторных клеток. Рецепторные клетки статоциста беспозвоночных, в отличие от типичных волосковых клеток позвоночных, являются первичночувствующими и имеют центральные отростки. Аксоны рецепторных клеток образуют статический нерв, оканчивающийся в церебральных ганглиях.
В статоцистах головоногих моллюсков, которые занимают в эволюции более высокое положение, наряду с макулами, выполняющими функцию гравитационных рецепторов и обеспечивающими статическое равновесие, имеются и кристы (группы волосковых клеток, покрытые купулами) — рецепторы угловых ускорений, обеспечивающие динамическое равновесие. Таким образом, с развитием двигательной активности животных статоцист, возникший как орган чисто статического равновесия, превращается в орган, обладающий способностью регуляции не только поведения организма в гравитационном поле, но и координации его движений в трехмерном пространстве.
Важно также подчеркнуть, что уже у беспозвоночных животных система статоциста функционально связана с другими сенсорными системами, например зрительной, хеморецепторной, системой рецепторов растяжения, что обеспечивает координацию двигательных актов, регуляцию мышечного тонуса и адаптивное поведение.
Наряду с гравитационной функцией система статоциста обладает способностью к сейсмо- и виброрецепции, а у некоторых высокоорганизованных форм и к акустическому приему. Это позволяет рассматривать сенсорную систему статоциста ракообразных и моллюсков в качестве аналога акустико-вестибулярных систем позвоночных животных.
У насекомых вместо статоцистов функцию рецепторов гравитации выполняют наружные проприоцепторы — волосковые пластинки, а функцию рецепторов угловых ускорений — жужжальца, которые напоминают гироскопы, предназначенные для стабилизации самолетов и подводных лодок.
У позвоночных животных происходит дальнейшее совершенствование вестибулярной системы, особенно ее центрального отдела. Рецепторы гравитации у всех позвоночных животных и человека располагаются в лабиринте. Он представлен двумя (у миноги) и тремя (у всех остальных позвоночных) полукружными каналами, утри- кулюсом (маточкой), саккулюсом (мешочком) и лагеной. У высших позвоночных в нем появляется улитка как вырост саккулюса.
Верхняя часть лабиринта (полукружные каналы и утрикулюс с их сенсорными элементами) выполняет в основном гравитационные функции (отолитовый аппарат утрикулюса — как датчик равновесия, полукружные каналы — как датчик угловых ускорений), а нижняя часть (саккулюс, улитка и лагена) — слуховые функции. Поэтому верхнюю часть лабиринта обычно выделяют в качестве преддверия, или вестибулярного аппарата, противопоставляя его органу слуха — улитке.
У всех позвоночных, включая высших млекопитающих животных и человека, рецепция силы тяжести и углового ускорения осуществляется с участием вторичночувствующих волосковых клеток, возбуждение которых через холинергические синапсы, т. е. с участием ацетилхолина передается на афферентные биполярные нейроны вестибулярного ганглия. Различают колбообразные и цилиндрические волосковые клетки. Оба вида клеток располагаются между поддерживающими (опорными) клетками.
Каждая рецепторная, или волосковая, клетка снабжена одной подвижной ресничкой, или киноцилией, и 60—80 склеенными между собой неподвижными волосками, или стереоцилиями. Преобразование действующей силы тяжести, линейного или углового ускорения в рецепторный потенциал осуществляется за счет изменения положения стереоцилий под влиянием отолитовой мембраны (в макулах утрикулюса и саккулюса) или купулы (в кристах полукружных каналов). Это связано с тем, что механическое смещение
стереоцилий изменяет проницаемость для ионов (вероятнее всего, для ионов калия).
Смещение стереоцилии в сторону киноцилии приводит к деполяризации, т. е. к возбуждению рецептора, а смещение в противоположном направлении вызывает гиперполяризацию, т. е. тормоз- ный эффект. Киноцилия регулирует степень скольжения, давления и точности отхождения на исходные позиции отолитовой мембраны в области каждой рецепторной клетки.
У всех млекопитающих макула утрикулюса воспринимает положение организма по отношению к гравитационному полю, т. е. служит рецептором гравитации. Макула саккулюса (кроме рыб) является вспомогательной по отношению к утрикулюсу и, кроме того, хорошо реагирует на вибрацию. Кристы ампул трех полукружных каналов воспринимают угловые ускорения.
На морских свинках показано, что площадь утрикулярной макулы составляет всего 0,55 мм 2 и на ней содержится примерно 9000 во- лосковых клеток, для саккулярной макулы эти значения составляют 0,50 мм 2 и 7500 клеток, а для каждой кристы латерального, переднего и заднего каналов — соответственно 0,40 мм 2 и 5500 клеток.
Отолитовый аппарат (рис. 3.3). В отолитовых частях лабиринта, т. е. в утрикулюсе и саккулюсе волосковые клетки собраны в виде возвышений, или макул (macula sacculi, macula utriculi). Киноцилии этих клеток либо прикреплены к крупному отолиту (рыбы), либо пронизывают толстую отолитовуто мембрану (остальные позвоночные), содержащую кристаллики шестиугольной формы из углекислого кальция (СаС03); эти кристаллики называют отокониями, или отолитами, или статолитами; все кристаллы склеены между собой желеобразным веществом. Таким образом, отолитовая мембрана представляет собой студенистую, войлокообразную массу, покрывающую всю макулу.
Дистальные концы стереоцилий волосковых клеток макулы свободно подпирают или «зачесывают» отолитовую мембрану. Эта мембрана тяжелее остальной ткани и может оказывать давление на волоски рецепторных клеток. Смещение отолитовой мембраны при изменении положения тела в гравитационном поле сопровождается появлением режущих сил, действующих на волоски рецепторных клеток. Тем самым макулы воспринимают изменение силы тяжести и линейного ускорения.
В макулах отолитовых органов ориентация киноцилии такова, что при смещении отолитовой мембраны одни волосковые клетки макулы возбуждается, другие — затормаживаются. В утрикулярной макуле, т. е. в основном рецепторе гравитации позвоночных, рецепторные клетки со сходной поляризацией собираются в группы, которые в совокупности образуют четырехплановую мозаику. Каждой части этой мозаики или квадранта может соответствовать определенная группа мышц, принимающих участие в установке тела и регулируемых функций утрикулярной макулы.
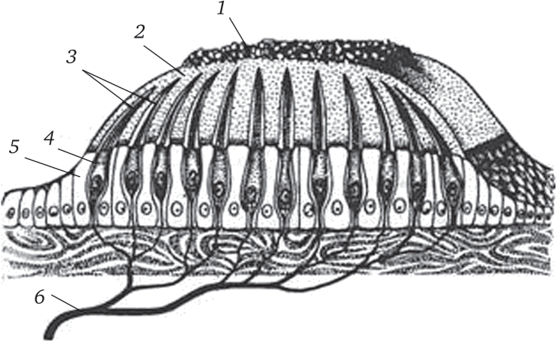
Рис. 3.3. Оттолитовый аппарат:
- 1 — отолиты; 2 — отолитовая мембрана; 3 — волоски рецепторных клеток;
- 4 — волосковые рецепторные клетки; 5 — опорные клетки; 6 — дендрит чувствительного нейрона
Отведение импульсной активности от волокон нервной веточки, иннервирующей утрикулярную макулу обезьяны, показало, что 70 % обследованных единиц возбуждалось при наклонах головы в ипсилатеральную сторону, 30 % — при наклонах в контралатеральную сторону. Кроме того, около 50 % единиц активировалось при наклонах «носом вверх» и примерно столько же при наклонах в противоположном направлении.
Эти результаты хорошо согласуются с особенностями пространственной поляризации волосковых клеток в утрикулярной макуле. С другой стороны, они показывают, что адекватным раздражением для утрикулюса являются наклоны головы (вправо, влево, вверх, вниз). Показано также, что рецепторный аппарат преддверия (утрикулюса и саккулюса) возбуждается при ускоряющемся или замедляющемся прямолинейном движении тела и тряске. Таким образом, отолитовый аппарат воспринимает как раздражения от ускорения или замедления прямолинейного движения, так и изменения в положении головы, вызывающие смещение отолитовой мембраны.
Перепончатые полукружные каналы. Волосковые клетки полукружных каналов, как уже отмечалось, сконцентрированы только в кристах ампул. Волоски погружены в полупрозрачную студенистую массу — купулу, которая не содержит отолитов и покрывает гребешок (рис. 3.4, Б). Эндолимфа, заполняющая перепончатый проток полукружных каналов, оказывает равномерное влияние на купулу, если голова находится в покое или если животное движется равно-
мерно и прямолинейно. Если же голова поворачивается в сторону, то давление эндолимфы на стенки полукружных каналов меняется. Вследствие инерции эндолимфа в канале смещается и сильнее давит на сторону, противоположную движению. Воспринимающие клетки реагируют на это изменение и посылают в мозг соответствующую информацию. Следовательно, воспринимающие клетки полукружных каналов реагируют на угловое ускорение, т. е. на перемену направления движения, если даже скорость его остается постоянной. Они воспринимают также вращательное ускорение и замедление.
Экспериментально показано, что в волосковых клетках кристы латерального (горизонтального) канала киноцилии в пучках стере- оцилий ориентированы в сторону утрикулюса, а в клетках переднего (сагиттального) канала — в сторону просвета канала. Такая морфологическая поляризация имеет, видимо, отношение к механизму направленной (дирекционной) возбудимости канала. Так, в горизонтальном канале ток эндолимфы вызывает возбуждающий эффект, если он направлен к утрикулюсу, а в сагиттальных — тормозной эффект.
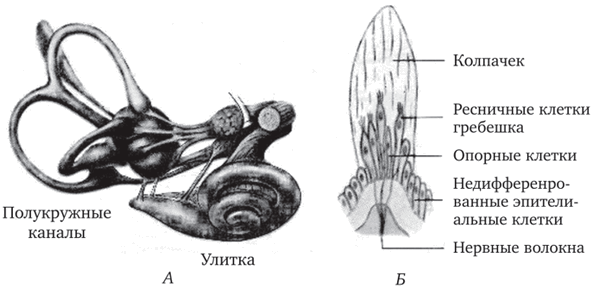
Рис. 3.4. Внутреннее ухо (А) и рецепторы на гребешках ампул (Б)
Итак, адекватным раздражением для полукружных каналов являются угловые ускорения и замедления, возникающие при вращении человека или животных в горизонтальной, фронтальной или сагиттальной плоскостях, а также смена направления движения, если даже скорость его остается постоянной.
Читайте также:
- Анатомия: Закономерности хода артерий от материнского ствола к органу.
- МРТ латеральных поддерживающих структур коленного сустава в норме
- Секреторная функция желудка. Желудочный сок. Состав желудочного сока.
- Лечение пульпита у летчиков. Отдаленные результаты лечения пульпитов
- УЗИ при краевом и оболочечном прикреплении пуповины плода